
Abstract
Due to the advancing agricultural frontier in the Brazilian Amazon, the present rate of deforestation engenders a pessimistic scenario for vertebrate diversity in the area. Protected areas are an essential conservation tool to limit biodiversity loss, but their efficiency have yet to be proven. Here, we used camera-trap data on the presence of medium and large-size vertebrates in a protected area (Cantão State Park) and a neighbouring private forest reserve (Santa Fé Ranch) to evaluate their effectiveness in protecting biodiversity. We also gathered information on seasonality and activity patterns. A total sampling effort of 7929 trap-nights revealed a diverse vertebrate fauna in the region. A total of 34 mammal species, belonging to 8 different orders was detected in the study area, some of which have a high level of conservation interest and value. The photographic index showed that diversity was more abundant outside the protected area of Cantão State Park, where seasonality could play a major role in vertebrate occurrence. Overall, the influence of seasonality on distribution appears to be species-specific. During the wet season around 40% of the common species were not detected inside the park, whereas in Santa Fé Ranch most species (62.5%) suffered only a slight decrease in relative abundance probably due to changes in the availability of food resources. Our results highlight the importance of private land for vertebrate conservation in the Amazon and alert to the need for increased law enforcement in these areas to secure biodiversity preservation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
With a growing human population and a higher demand for natural resources, protected areas represent one of the main conservation measures used to avoid species extinction and habitat loss (Cardillo et al. 2004; Joppa et al. 2009). Despite covering over 12% of earth’s terrestrial surface, there are still some issues concerning the efficiency of protected areas in securing species richness preservation (Parrish et al. 2003). One of the questions that remain open is the usefulness of private reserves in preserving the extraordinary biodiversity of tropical forests, such as in the Amazon. In Brazil, the agricultural frontier is quickly advancing, with a concomitant fragmentation of the Amazon forest. By law, new farms and ranches must establish a private forest reserve that covers 80% of the private land accessioned. In theory, this legal requirement preserves, by far, much more land than the more traditional public protected areas. However, it is not clear how useful they are in preserving biodiversity because most of our conservation and scientific efforts are focused towards public reserves.
The Amazon basin represents one of the most important regions in the world in terms of biodiversity (Costa et al. 2005; Stone et al. 2009). This, the largest rainforest on the planet, harbours the greatest number of endemic species per unit area and one of the highest diversities of vertebrates (IUCN et al. 2008). Habitat loss is the most important threat to vertebrate species and, even if relatively well known, only a small number of studies provide information on their status in this tropical forest (Voss and Emmons 1996; Fonseca et al. 1999). The rate of deforestation of Brazilian Amazonia has reached considerably high rates in recent years and it is estimated that the advance of the agricultural frontier over the region could cause a reduction by 50% of its forest cover in a few decades and, with it, the majority of its vertebrate diversity (Laurence et al. 2001; Azevedo-Ramos et al. 2006). This scenario highlights the imperativeness of establishing an effective network of protected areas (Schulman et al. 2007). In particular, within the “arc of deforestation” the eastern/south-eastern region of the Amazon, where increased human pressure results in a highly fragmented landscape of agricultural farms and forest patches, evaluation of the conservation value of the remaining forest fragments is needed (Lopes and Ferrari 2000; Morton et al. 2006).
There has been an increase in research on the distribution and ecology of medium and large-sized mammals and birds in the tropics, but their biological characteristics (nocturnal, low density and cryptic) make them difficult to census and study (Silveira et al. 2003; Lyra-Jorge et al. 2008). The use of camera trapping in surveys has been intensified recently due to the advantages of being cost-effective for detecting animals with inconspicuous habits and its reduced disturbance (Zielinski et al. 1995; Kelly 2008; Rowcliffe and Carbone 2008). It constitutes a non-invasive method that provides considerable information on occurrence, population density and other biological parameters (sociality, activity or reproduction) of target and non-target species (Silveira et al. 2003; Gómez et al. 2005; Stein et al. 2008).
This research is part of a long-term monitoring programme for jaguar density. Our intensive sampling effort over several years has resulted in a considerable number of photos of medium to large-sized vertebrates (mostly mammals, and to a limited extend birds) within Cantão State Park (CS-Park) and the adjacent Santa Fé Ranch (SF-Ranch). We use this information to report on vertebrate species richness, their activity patterns and the influence of seasonality and flooding both in a public reserve (CS-Park) and in a private forest fragment within a cattle ranch (SF-Ranch) in the Amazon arc of deforestation.
Materials and methods
Study area
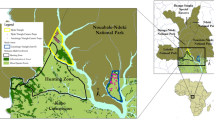
The study was carried out in the middle Araguaia river basin in two areas on opposite sides of the river: the Cantão State Park (CS-Park) on the right river bank and the Santa Fé Ranch (SF-Ranch) on the left river bank (Fig. 1). Cantão State Park (09°36′ S, 50°03′ W) is an 89,000 ha conservation unit situated in the transitional area between the Amazon and Cerrado biomes. Water abundance undergoes dramatic cyclical changes due to an extended network of rivers, canals and lakes. The dynamics created by the wet season (November–March) and the prolonged dry season (April–October) influences vegetation structure (SPMA 2000; Vitt et al. 2007). With an annual average precipitation of 1,710 mm/year and a difference of more than 4 m in river level between seasons (data from Santa Fé Ranch), flooding conditions influence available resources (food and shelter) for the fauna (Fig. 2). The vegetation, mainly represented by secondary growth tropical rainforest with some small grassland areas, suffers partial flooding during the wet season.

Study area showing the Santa Fé Ranch and Cantão State Park and their ecotonal location in the Brazilian biomes of the Amazon and Cerrado

Average precipitation and river level recorded for the Cantão State Park region, central Brazil (data from Santa Fé Ranch)
Santa Fé Ranch (09°34′ S, 50°21′ W) is a 65,000 ha beef cattle ranch in southeast Pará State, bordering the Araguaia River. Around 65% of the ranch represents a continuous, semi-deciduous seasonal tropical forest (similar as the CS-Park) patch that extrapolates the farm boundaries, while the other 35% is occupied almost entirely by pastures.
Field methods
We conducted five camera trap surveys between July 2005 and November 2007 during both the dry (three surveys) and wet (two surveys) seasons. A variable number of stations (from 10 to 22) were established throughout each study area, with a distance of 1–3 km between stations (Rabinowitz and Nottigham 1986; Karanth and Nicholds 2002; Silver et al. 2004). One passive infra-red camera was placed approximately 50–70 cm above ground on dirt roads or trails (animal or human made) at each station, except during the 2007 dry season survey at SF-Ranch when two cameras per station were used (Silver 2004). Two different camera types were used: Camtrakker® (Cam Trakker, Watkinsville, USA) and C1-BU® (Vibrashine Inc., Taylorsville, MS 3968, USA). Each camera was programmed to take photographs 24 h/day with a 5-min interval between photos. All stations were checked on a regular basis (5–20 days) throughout all surveys for maintenance. In addition to camera trapping, field observations contributed data on the occurrence of primates and aquatic mammals during the expeditions for camera installation and monitoring.
Data analysis
Species identification, number of individuals, sex, age, date and hour were determined, where possible, for each photograph. Following O’Brien et al. (2003), photos were considered independent events only if they met at least one of three criteria: consecutive photographs of different individuals of the same or different species; consecutive photographs of individuals of the same species taken more than 1 h apart; non-consecutive photos of individuals of the same species.
The relative abundance index (RAI) was determined for all species by dividing the number of independent photo captures, by effort (trap-nights) times 100 (O’Brien et al. 2003; Kawanishi and Sunquist 2004). A photographic base index of abundance is considered a consistent method to infer the relative abundance of cryptic mammals, assuming that the cameras did not affect the rates of movement of animals (Carbone et al. 2001; Goulart et al. 2009). The criteria used to set-up the cameras was always to maximise jaguar detection and, therefore, we assume that the same bias in species detection occurs across the entire study area, making the data comparable between sites and over time (Stein et al. 2008).
We anticipated that populations would remain relatively stable during our short study period (3 years) and also within seasons in the absence of any catastrophic phenomenon (Krebs 1994; Harmsen 2006). Taking this into account, results from different surveys were pooled and we compared RAI for the most frequent species between seasons (dry and wet) for both areas (SF-Ranch and CS-Park) using Mann–Whitney tests.
Factors affecting diversity and species detection
Using generalised linear mixed models (GLMM), we evaluated the factors that could affect species richness according to sampling features (year, season, area: CS-Park vs. SF-Ranch) and environmental predictors (distance to water, distance to pasture, camera location i.e. road vs. trail), considering the number of different species as the dependent variable and trap station included as a random variable. We also analysed the factors affecting RAI according to some species characteristics: weight, conservation status, trophic niche (predator vs. prey) and social behaviour (solitary vs. group-living), using RAI as the dependent variable. We used R v.8.2 free statistical software and the Lme4 package for mixed models (Bates and Sarkar 2006) to fit the statistical models.
Activity patterns
We generated the activity pattern in 1-h intervals for those species with more than 10 independent photographic events. We used Chi-square tests to compare data from CS-Park and SF-Ranch, pooling them for further analysis if they were not significantly different (P > 0.05). Then, each capture was classified into three categories: nocturnal (18:31–05:00 h), diurnal (06:31–17:00 h), and crepuscular (17:01–18.30 and 05:01–06:30 h) (Schaik and Griffiths 1996).
Results
Species richness
A total sampling effort of 7929 trap-nights were conducted over several, continuous, two-month periods (average 61 days), with a variable number of camera stations (average 15; range 10–22) and trap nights (965 trap-nights; 525–1681) at each site (Table 1). Camera trap effort on CS-Park was lower (3183 trap nights) than in SF-Ranch (4746 trap nights), but sampling season lasted on average 61 days in both areas, and the average number of camera stations established on each site was slightly higher in CS-Park (18) than in SF-Ranch (15). In 2007 we only sampled SF-Ranch.
Mammals constituted the greater proportion of the utilised photos (65.7%), followed by birds (32.9%) and reptiles (1.4%). This proportion nevertheless changes, when the two study areas are considered separately. In CS-Park, 53.9% of the photo captures were of birds, compared to 44.6% for mammals and 1.1% for reptiles. On SF-Ranch, mammals were the most frequently captured group (70.2%), followed by birds (28.4%) and reptiles (1.4%).
A total of 34 mammal species, belonging to eight different orders was detected in the study area during the course of the survey period (Table 2). We registered more mammal species in the SF-Ranch forest reserve (N = 30) than in CS-Park (N = 16), where some common species like brocket deer (Mazama spp.), peccaries (Tayassu pecari and Pecari tacaju), crab-eating fox (Cerdocyon thous) and armadillos (Priodontes maximus and Dasypus novemcinctus) were absent, contrary to a previous census carried out in the park (Silveira 2004).
In terms of conservation status, we emphasise the presence of two endangered species (EN), Giant Otter (Pteronura brasiliensis) and Uta Hick’s Bearded Saki (Chiropotes utahickae); and three species classified as vulnerable (VU) according to IUCN (2008), Brazilian Tapir (Tapirus terrestris), Marsh Deer (Blastocerus dichotomus) and Giant Armadillo (Priodontes maximus) (Table 2) in the studied areas. Two aquatic species, the Pink River Dolphin (Inia geoffrensis) and the Giant Otter (Pteronura brasiliensis) were only present in CS-Park. Thus, comparing the two sampled areas, we recorded four species classified under Threatened Categories (EN or VU) from SF-Ranch, whereas only three were recorded from CS-Park (Table 2).
Photo capture rate differences between places and seasons
All the species seem to be more abundant in SF-Ranch than in the CS-Park, with the exception of Cuniculus paca and Crax fasciolata (Table 3). When comparing RAI between seasons we can verify the absence of some species during the wet season, such as Puma concolor, Mazama americana, Hydrochaeris hydrochaeris and Penelope spp. inside CS-Park and Mazama gouazoupira, Didelphis spp., Hydrochaeris hydrochaeris and Mitu tuberosum in SF-Ranch. For CS-Park, some species show a reduction (Leopardus pardalis, Didelphis spp. and Crax fasciolata) or increase (Tapirus terrestris, Panthera onca and Cuniculus paca) in RAI from the dry to the wet season, but the difference is only significant for the jaguar (Mann–Whitney W = 32; P < 0.001). The majority of the species that occur in SF-Ranch reveal a fluctuation in capture rates from the dry to the wet season. However, only the reduction of Mazama americana, Pecari tacaju, Cerdocyon thous, Leopardus pardalis, Dasyprocta azarae, Penelope spp. and Crax fasciolata were significant (Mann–Whitney test P < 0.05 for all comparisons).
Biological and sampling factors affecting RAI
GLMM analysis revealed that the number of species detected at each station was significantly higher outside the park than inside (Table 4). Also RAI increased with the number of days of sampling during the study years and revealed a negative correlation with season (lower richness during the wet season). Camera location seemed to be of importance, resulting in a higher number of species photographed on roads compared to trails.
The relationship between the photographic rate (RAI) and some species characteristics reveal that detection was positively and strongly dependent on animal body mass (favouring larger species) and aggregation (species that moved in groups) (Table 4). Considering that threatened level is directly associated with abundance, it is logical that vulnerable species were less frequently captured than less threatened ones, which was confirmed by a negative correlation. Trophic niche, on the other hand, did not present a significant association with RAI.
Activity patterns
We determined the activity pattern for 16 species of mammals and birds. Most were nocturnal, with the exception of the gray brocket deer (Mazama gouazoupira), collared peccary (Pecari tajacu) and agouti (Dasyprocta azarae) (Fig. 3). All of the bird species presented crepuscular/diurnal behaviour. The jaguar was significantly more diurnal inside the park than in SF-ranch (χ2 = 55.71, DF = 22, P < 0.001). There were no differences in the activity patterns of the other common species between both areas.

Activity patterns of some mammal and bird species in the study area in central Brazil, recorded by camera trapping (nocturnal 18:31–05:00 h, diurnal 06:31–17:00 h and crepuscular 17:01–18.30 and 05:01–06:30 h)
Discussion
Photo capture rates are biased towards animals that spend most of the time on the ground and, since cameras were set to detect and evaluate jaguar density, it is also biased towards larger and more abundant animals, making the inventory of mammal species present in the area incomplete. Nevertheless, it contributes important information concerning the occurrence of mammals with different conservation status along the Amazonian agricultural frontier where there is no room for complacency about the security of species not currently considered globally threatened (Cardillo et al. 2004). For the majority of the mammal species the activity patterns found in our study area are similar to those described in the literature, with a typical predominance of nocturnal species (Gómez et al. 2005). Nevertheless, as occurs in CS-Park, jaguar can be considerably active in the daytime in areas of the Amazon and in the Pantanal, which could be associated with main prey species activity (Schaller and Crawshaw 1980; Crawshaw and Quigley 1991; Gómez et al. 2005; Weckel et al. 2006).
CS-Park is influenced by a strong seasonal environment: the flooding regime during the winter season covering around 70% of its area between the Araguaia and Côco Rivers (SMPA 2000), making it very susceptible to catastrophic flooding or, at other times, even droughts. The consequences in terms of habitat availability for terrestrial species are evident, forcing animals to move in search of more suitable habitat and causing seasonal differences in species occurrence and abundance inside the park. Comparing the results on species richness in CS-Park with a previous inventory done by Silveira (2004) using the same methodological approach, we show that 42.3% (N = 21) of the mammal species previously reported in the area were not detected in our three subsequent sampling periods. The number of mammal photos actually decreased from 8–10 photos/100 trap nights to 3–4 photos/100 trap nights from 2002–2003 to 2005–2006, respectively. Also, a decreased pattern can be observed in RAI from 2002–2003 to 2005–2006 for almost all the species, despite a superior sampling effort (2615 camera-nights for 2002–2003 and 3183 camera-nights for 2005–2006) (Fig. 4). These differences can be explained by the extremely wet 2004 season that flooded almost the entire park as a result of elevated precipitation and the consequent increase in river water levels (Brazilian National Water Agency data). During the 2004 flooding many populations of terrestrial mammals probably dispersed from CS-Park in search of more suitable dry land, which is mainly available in the areas surrounding the park. We heard several anecdotal accounts of abnormal numbers of drowned animals and considerable numbers of animals crossing the river during that period. If reductions in the mammal population occur inside the park during wet seasons, the re-colonisation process totally depends on the level of habitat preservation in the surrounding areas, which is mainly occupied by agricultural farms (with their forest reserves) and human rural settlements. Some species with higher mobility, like Panthera onca and Tapirus terrestris, may continue to use CS-Park during the wet season, exploiting the small areas that remain above water. Due to the reduced areas of dry land, a concentration of activity in these areas could result in the increased capture rates (RAI) of these species. In contrast, due to the severe flood in 2004, other species may have been extirpated from the CS-Park area and have not managed to reoccupy the park since. Only Cuniculus paca, Nasua nasua and Hydrochaeris hydrochaeris showed an increase in RAI after 2003.

Photographic rate fluctuation between different years of sampling in Cantão State Park for the most common mammal and bird species (2002–2003 data from Silveira 2004)
The forested reserve in SF-Ranch is less effected by river flooding and, thus, animal distribution throughout the seasons should be based on other resource availability; namely food (Bodmer 1990). In fact, the majority of species demonstrated a decrease in relative abundance, especially the large herbivorous such as Mazama spp., Pecari tacaju, Tapirus terrestris. Mendes Pontes and Chivers (2007) observed how, in an area in central Amazonia, fluctuations in food supply regulated forest use by peccary species and consequently conditioned jaguar and puma whereabouts. With widely distributed food resources during the wet season, peccaries exploited forested areas less frequently and presented broader spatial use. Our results concerning RAI variation for herbivorous species within seasons supports this. In order to understand the influence of seasonality on mammal movement in forested areas in central Brazil further research should be performed, enlarging the scale beyond forest limits, supported by a carefully structured sampling design and taking into account food availability measures.
Our camera trapping research revealed mammal diversity in a region covering both a protected and private area within the “arc of deforestation”. The results highlight the importance of private forest reserves for mammal conservation and the misleading idea that nature reserves per se can secure species richness. On the other hand, private forest reserves can be more susceptible to pressure (e.g. deforestation and hunting) due to lower law enforcement, especially in the Amazonian agricultural frontier (Cardillo et al. 2004). In this region, for conservation measures to succeed, combined action within protected and private areas should be established.
References
Azevedo-Ramos C, Do Amaral BD, Nepstad DC, Soares Filho B, Nasi R (2006) Integrating ecosystem management, protected areas, and mammal conservation in the Brazilian Amazon. Ecol Soc 11(2):17 [online] URL:http://www.ecologyandsociety.org/vol11/iss2/art17/
Bates D, Sarkar D (2006) Lme4: linear mixed-effect models using S4 classes. http://www.R-project.org
Bodmer RE (1990) Fruit patch size and frugivory in lowland tapir (Tapirus terrestris). J Zool 222:121–128
Carbone C, Christie S, Conforti K, Coulson T, Franklin N, Ginsberg JR, Griffiths M, Holden J, Kawanishi K, Kinnaird M, Laidlaw R, Lynam A, Macdonald DW, Martyr D, McDougal C, Nath L, O’Brien T, Seidensticker J, Smith DJL, Sunquist M, Tilson R, Shahruddin WN (2001) The use of photographic rates to estimate densities of tigers and other cryptic mammals. Anim Conserv 4(1):75–79
Cardillo M, Purvis A, Sechrest W, Gittleman JL, Bielby J, Mace GM (2004) Human population density and extinction risk in the world’s carnivores. Plos Biol 2:909–914
Costa LP, Leite YLR, Mendes SL, Ditchfield AD (2005) Mammal conservation in Brazil. Conserv Biol 19(3):672–679
Crawshaw PG, Quigley HB (1991) Jaguar spacing, activity and habitat use in a seasonally flooded environment in Brazil. J Zool 223:357–370
Fonseca GAB, da Herrmann G, Leite YLR (1999) Macrogeograhy of Brazilian mammals. In: Eisenberg JF, Redford KH (eds) Mammals of the Neotropics: the central Neotropics, vol 3. The University of Chicago Press, Chicago USA, Ecuador, Peru, Bolivia, Brazil, pp 549–563
Gómez H, Wallace RB, Ayala G, Tejada R (2005) Dry season activity periods of some Amazonian mammals. Stud Neotrop Fauna Environ 40(2):91–95
Goulart FVB, Cáceres NC, Graipel ME, Tortato MA, Ghizoni IR, Oliverira-Santos LGR (2009) Habitat selection by large mammals in southern Brazilian Forest. Mammal Biol 74(3):182–190. doi:10.1016/j.mambio.2009.02.006
Harmsen BJ (2006) The use of camera traps for estimating abundance and studying the ecology of jaguars (Panthera onca). PhD Dissertation, University of Southampton
IUCN, Conservation International, Arizona State University, Texas A&M University, University of Rome, University of Virginia, Zoological Society London (2008) An Analysis of Mammals on the 2008 IUCN Red List. www.iucnredlist.org/mammals. Cited on 12 December 2008
Joppa LN, Loarie SR, Pimm SL (2009) On population growth near protected areas. PLoS One 4(1):e4279
Karanth KU, Nicholds JD (2002) Monitoring tigers and their prey: a Manual for researchers, managers and conservationists in Tropical Asia. Center for Wildlife Studies, Bangalore, India, p 193
Kawanishi K, Sunquist ME (2004) Conservation status of tigers in a primary rainforest of Peninsular Malaysia. Biol Conserv 120:329–344
Kelly M (2008) Design, evaluate, refine: camera trap studies for elusive species. Anim Conserv 11:182–184
Krebs CJ (1994) Ecological methodology. Addison-Welsey Educational Publishers Inc, Menlo Park
Laurence WF, Cochrane MA, Bergen S, Fearnside PM, Delamonica P, Barber C, D’Angelo S, Fernandes T (2001) The future of the Brazilian Amazon. Science 291:438–439
Lopes MA, Ferrari SF (2000) Effects of human colonization on the abundance and diversity of mammals in eastern Brazilian Amazonia. Conserv Biol 14(6):1658–1665
Lyra-Jorge MC, Ciocheti G, Pivello VR, Meirelles ST (2008) Comparing methods for sampling large- and medium-sized mammals: camera traps and track plots. Eur J Wildl Res 54:739–744
Mendes Pontes AR, Chivers DJ (2007) Peccary movements as determinants of the movements of large cats in Brazilian Amazon. J Zool 273:257–265
Morton DC, DeFries RS, Shimabukuro YE, Anderson LO, Arai E, Espirito-Santo F, Freitas R, Morisette J (2006) Cropland expansion changes deforestation dynamics in the southern Brazilian Amazon. Proc Natl Acad Sci 103(39):14637–14641
O′Brien T, Kinnard MF, Wibisono HT (2003) Crouching tigers, hidden prey: Sumatran tiger and prey populations in a tropical forest landscape. Anim Conserv 6:131–139
Parrish JD, Braun DP, Unnasch RS (2003) Are we conserving what we say we are? Measuring ecological integrity within protected areas. Bioscience 53:851–860
Rabinowitz AR, Nottigham BG Jr (1986) Ecology and behaviour of the jaguar (Panthers onca) in Belize, Central America. J Zool 210(1):149–159
Rowcliffe JM, Carbone C (2008) Surveys using camera traps: are we looking to a brighter future? Anim Conserv 11:185–186
Schaik CP, Griffiths M (1996) Activity periods of Indonesian rain forest mammals. Biotropica 28:105–112
Schaller GB, Crawshaw PG (1980) Movement patterns of jaguar. Biotropica 12:161–168
Schulman L, Ruokolainen K, Junikka L, Sääksjärvi IE, Salo M, Juvonen S-K, Salo J, Higgings M (2007) Amazonian biodiversity and protected areas: do they meet? Biodivers Conserv 16:3011–3051
Silveira L (2004) Ecologia comparada e conservação da onça-pintada (Panthera onca) e onça-parda (Puma concolor), no Cerrado e Pantanal. PhD Dissertation, University of Brazilia (Brazil)
Silveira L, Jácomo ATA, Diniz-Filho JAF (2003) Camera trap, line transect census and track surveys: a comparative evaluation. Biol Conserv 114:351–355
Silver S (2004) Assessing jaguar abundance using remotely triggered cameras. Wildlife Conservation Society, New York, p 25
Silver S, Ostro LE, Marsh LK, Maffei L, Noss AJ, Kelly MJ, Wallace RB, Gómez H, Ayala G (2004) The use of camera traps for estimating jaguar Panthera onca abundance and density using capture/recapture analysis. Oryx 38(2):148–154
SPMA-Secretaria de Planejamento e Meio Ambiente (2000) Avaliação ecológica rápida do Parque Estadual do Cantão. Governo do Estado do Tocantins, Palmas, Tocantins, Brazil, p 133
Stein AB, Fuller TK, Marker LL (2008) Opportunistic use of camera traps to assess habitat-specific mammal and bird diversity in north-central Namibia. Biodivers Conserv 17:3579–3587
Stone AI, Lima EM, Aguiar GFS, Camargo CC, Flores TA, Kelt DA, Marques-Aguiar SA, Queiroz JAL, Ramos RM, Silva Júnior JS (2009) Non-volant mammalian diversity in fragments in extreme eastern Amazonia. Biodivers Conserv 18(6):1685–1694. doi:10.1007/s10531-008-9551-9
Vitt LJ, Shepard DB, Caldwell JP, Veira GHC, França FGR, Colli GR (2007) Living with your food: geckos in termitaria of Cantão. J Zool 272(3):321–328
Voss R, Emmons LH (1996) Mammalian diversity in Neotropical lowland rainforest: a preliminary assessment. Bull Am Museum Nat Hist 230:1–115
Weckel M, Giuliano W, Silver S (2006) Jaguar (Panthera onca) feeding ecology: distribution of predator and prey through time and space. J Zool 270:25–30
Zielinski WJ, Kucera TE, Barrett RH (1995) Current distribution of the fisher, Martes pennanti, in California. Calif Fish Game 81:104–112
Acknowledgments
This study was funded by the Jaguar Conservation Fund, Ideawild and Ecotropical Institute. Nuno Negrões was supported by a grant from Fundação para a Ciência e a Tecnologia (FCT-MCT) (SRFH/BD/23894/2005). We are indebted to Naturatins, the Cantão State Park personnel and Fazenda Santa Fé staff; especially to Marcos Mariani, who allowed this study on his property as well as for his logistic support.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Negrões, N., Revilla, E., Fonseca, C. et al. Private forest reserves can aid in preserving the community of medium and large-sized vertebrates in the Amazon arc of deforestation. Biodivers Conserv 20, 505–518 (2011). https://doi.org/10.1007/s10531-010-9961-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-010-9961-3