
Abstract
Habitat suitability models developed for non-native, invasive species often implicitly assume that projected invasion risk equates to risk of impact. I aim to test to what extent this assumption is true by comparing commonly-used invasive plant distribution datasets to abundance records. I compared herbarium occurrence records (downloaded from an online database) and regional occurrence records (compiled from individual states) to abundance estimates collected from over 300 invasive plant experts for 9 invasive species in the western U.S. I also created habitat suitability models (HSMs) using these datasets and compared the areas of predicted suitability. Sixty percent of the time, herbarium occurrences were located in regions where the species was rare enough to be undetected by experts, while only 26 % coincided with locations identified as having high abundance. Regional occurrences were located in areas where the species was not detected 32 % of the time, and on high abundance 42 % of the time. HSMs based on herbarium records encompassed 89 % of land area at risk of abundance, but overestimated the area of estimated risk (27–46 % false positive rate). HSMs based on regional occurrences had a smaller false positive rate (22–31 %), but encompassed only 67–68 % of area suitable for abundance. Herbarium records are strongly skewed towards locations with low invasive plant abundance, leading to invasion risk models that vastly overestimate abundance risk. Models based on occurrence points should be interpreted as risk of establishment only, not risk of abundance or impact. If HSMs aim to be more management relevant, invasion risk models should include abundance as well as occurrence.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Non-native invasive plant species are well known threats to native ecosystems (Mack et al. 2000), with transformative invaders (sensu Richardson et al. 2000) causing significant ecological (e.g., Wilcove et al. 1998; Vilà et al. 2011) and economic (Pimentel et al. 2005) damage. In an effort to forecast future invasions, models of invasion risk based on correlations between species distribution and environmental variables (hereafter habitat suitability models) have become increasingly common (Table 1), and are often presented as a tool for managers to prioritize treatment for preventing invasions in their early stages. However, habitat suitability models (HSMs) for invasive plants are almost universally based on occurrence data, which to date have an unknown relationship with invasive plant impact. Occurrence points have a clear link to risk of species establishment (Lockwood et al. 2007), but how well do they model risk of impact?
Management plans for invasive species often focus on the potential impacts of that species as a means of assessing risk (Parker et al. 1999; Hulme 2006). And, because the effects of individual invasive species are so difficult to identify (Parker et al. 1999), abundance is often presented as a proxy for impact (Ricciardi 2003; Kulhanek et al. 2011). Abundance of invasive plants in particular likely correlates with impact. For example, in agricultural systems, managers are concerned about invasive plants that are or are likely to become abundant and damaging to crops (McDonald et al. 2009). Invasive plants that establish, but maintain low densities are of minimal concern (McDonald et al. 2009). Similarly, in arid ecosystems, invasion of annual plants is primarily a concern where those species are (or are likely to become) abundant enough to carry fire (D’Antonio and Vitousek 1992; Brooks et al. 2004). Where invasive species are actively being controlled, the management aim is not to extirpate the species (a virtually impossible task), but to reduce the population to a level where it is no longer abundant, and hence no longer problematic (see for examples Hulme 2006). Thus, an assumption that modeled establishment risk is the same as impact risk, when in fact it is not, could waste limited management resources by unnecessarily targeting non-native species in areas where they are unlikely to become problematic.
Impact does not relate only to abundance. Some species invasions are problematic more as a result of per capita effect rather than abundance. For example, small numbers of non-native species with unique qualities (‘quirks’ sensu Simberloff 1985; Parker et al. 1999) could alter an ecosystem as much as high abundance of a species with low per capita effect. Per capita effects of the same species can also vary with both abundance and environmental conditions (e.g., Brewer 2011), reinforcing the importance of both characteristics. Either abundance or per capita effect within the species range can be considered a reasonable proxy for impact (Parker et al. 1999), and if the species is known to have a large effect as a single individual then establishment risk alone would be an appropriate approximation of impact risk. However, given that plant abundance clearly relates to impact in many cases and given the difficulty of measuring per capita effect (and corresponding lack of data for most species), abundance is currently the more viable option as a proxy for impact.
To date, most projections of both current and future invasion risk based on HSMs rely on occurrence data alone. This practice identifies the potential range of invasion, or the environmental requirements of the modeled species (Leibold 1995). Indeed, it is common for HSMs to use both native and invaded ranges when predicting invasion risk (e.g., Peterson and Vieglais 2001; Loo et al. 2007) to identify all locations where the invader could potentially establish (i.e. the ‘establishment niche’). However, this practice ignores any consideration of impact that invasive species have on the environment. Use of the term invasion risk implies risk of impact. In 1995, Leibold called for greater distinction of the ‘impact niche’ (i.e. locations where the invader could potentially have an impact) from the establishment niche. But, these concepts remain muddled in invasion biogeography, with only a handful of studies explicitly modeling ‘impact niche’ based on areas of high abundance (Ricciardi 2003; McDonald et al. 2009; Kulhanek et al. 2011).
Studies from other taxonomic groups suggest that occurrence records are ineffective predictors of abundance. For example, VanDerWal et al. (2009) regressed measured abundance of 69 vertebrate species against suitability modeled from occurrences and found a significant correlation. However, the correlation coefficient (R2) averaged only 0.12 across all species (VanDerWal et al. 2009), and other similar studies have found either no significant correlation (Jiménez-Valverde et al. 2009; Tôrres et al. 2012), or that the relationship is best represented as a step function (Pearce and Ferrier 2001; Nielsen et al. 2005; Estes et al. 2012). After a quickly reached threshold, increasing environmental suitability modeled by HSMs typically correlates poorly (or not at all) to increasing abundance.
As a result, a more management-relevant approach to modeling invasion risk could be to create HSMs based on locations with high invasive species abundance (Ramcharan et al. 1992; Ricciardi 2003; McDonald et al. 2009; Kulhanek et al. 2011), which translate more closely to impact risk. HSMs for invasive plants typically employ either occurrence records from herbaria, or from ancillary records compiled from local or regional datasets (e.g., Crall et al. 2006) (Table 1). But, how well do these occurrence points represent abundance (and therefore impact)? And, how similar are models based on occurrence data (establishment niche) to models based on abundance data (impact niche)? Here, I compare herbarium occurrence records and regionally-compiled occurrence records to abundance and occurrence data for nine invasive species in the western U.S. I determine the extent to which occurrence records represent areas of high invasive species abundance, and compare suitability model results based on the four datasets to determine how well typical HSMs estimate risk of invasive species impact.
Methods
Collection of abundance data
This research targeted nine problematic invasive plant species in the western U.S.: Acroptilon repens (Russian knapweed), Bromus tectorum (cheatgrass), Centaurea maculosa (spotted knapweed), Cirsium arvense (Canada thistle), Cynoglossum officinale (hound’s tongue), Lepidium draba (white top), Lepidium latifolium (perennial pepperweed), Linaria dalmatica (Dalmatian toadflax), and Taeniatherum caput-medusae (medusahead). These species were selected because they have large ecological impact in the areas where they are abundant and each is a management priority in some portion of the study area. Abundance estimates for these species were based on the expert knowledge of over 300 county weed managers employed by state departments of agriculture, or the bureau of land management (Bradley and Marvin 2011). These experts were very familiar with the location and extents of invasions in their counties, and were familiar with all of the target species due to widespread concern about their invasion across the western U.S. However, the resulting maps have not been validated, so it is likely that this dataset suffers from both under-reporting and over-reporting. Further, although the collected data are distributed across fifteen western states (Washington, Oregon, California, Idaho, Nevada, Montana, Wyoming, Utah, Arizona, Colorado, New Mexico, North Dakota, South Dakota, Nebraska, and Kansas), the coverage is not complete and some spatial bias likely exists. Experts estimated qualitative abundance of the nine species on paper maps with 4 × 4 km grid cells overlaid on the county or land area they manage. These results were then digitized, creating abundance estimates for 43 % of the western U.S. with mapped area distributed nearly evenly across all states except Arizona and New Mexico, which had fewer responses. Of the nine species mapped, B. tectorum and C. arvense were by far the most widespread and regionally abundant invaders (Bradley and Marvin 2011).
Abundance estimates included the following potential rankings: not detected, rare, few small invasions, many small invasions, and many large invasions. For comparison with occurrence locations, I combined those categories into three: absent (from not detected), low abundance (from rare and few small invasions), and high abundance (from many small and many large invasions).
Comparing point occurrence locations to abundance
I compared abundance estimates to point occurrences of the nine invasive plants collected from one of two sources: herbarium occurrences or regional management occurrences. Herbarium occurrences were downloaded from records compiled by the Global Biodiversity Information Facility (GBIF).Footnote 1 This online, searchable database of museum and herbarium records is frequently used as a source of distribution data for habitat suitability modeling and invasion risk assessments. Regional management occurrences were compiled from Oregon weedmapper (weedmapper.org), Nevada natural heritage (heritage.nv.gov), Idaho department of agriculture (agri.state.id.us), and the southwest exotic plant mapping program (SWEMP). Regional management occurrences were collected by both scientists and managers, often with a specific focus on invasive species control.
I compared herbarium and regional occurrence records to gridded abundance estimates for each species. Only occurrence points that overlapped the abundance map were retained, and duplicate records intersecting the same 4 km abundance grid cell were excluded. I counted the number of occurrence records in the herbarium and regional datasets that intersected abundance estimates of absent, low abundance, and high abundance for each of the nine species. I also averaged the data across all species to quantify how well occurrence points represent invasive plant abundance.
Comparing modeled invasion risk
In addition to evaluating the occurrence datasets relative to abundance, I compared the spatial extents of invasion risk maps created using habitat suitability modeling. For this analysis, I created four separate models for each species, one based on all of the available herbarium occurrences (not just those overlapping the abundance dataset), a second based on all of the regional management occurrences, and a third based on points at the center of all expert-identified occurrence points (both low and high abundance). These three datasets were each compared to suitability models based on expert-identified locations of high abundance. To create the suitability models, I used MAXENT version 3.3.3 (Phillips et al. 2006), which uses a maximum entropy methodology as well as Mahalanobis distance (MD; calculated in Matlab version 7.10), which calculates distance from a centroid of n-dimensional climate space.
Suitability models such as MAXENT which use occurrence points and generate pseudo-absences may be prone to overfitting occurrence data (Jiménez-Valverde et al. 2008; Lobo et al. 2010) thereby underestimating invasion risk. Presence-only models such as MD (see for examples Tsoar et al. 2007) might therefore be a better choice for erring on the side of overprediction, particularly if occurrence data are likely to be biased (Jiménez-Valverde et al. 2008) as often happens early in an invasion (Václavík and Meentemeyer 2012). I chose to use MAXENT and MD in this case in order to ensure that any observed differences in modeled suitability are a result of the input data and not the choice of model.
In order to reduce sampling bias in MAXENT’s pseudo-absence dataset, I applied a bias correction to the analysis to limit the extents of pseudo-absence points to areas likely to have been sampled (thereby making pseudo-absences a better approximation of actual absences). For the expert-identified occurrence and abundance records, the bias file extents included all sampled counties. For regional management and herbarium records, the bias file included all areas within one 4 km pixel of a recorded occurrence for any species in the regional and herbarium datasets respectively. The total area modeled was limited to the 15 western U.S. states sampled for expert knowledge. All other parameters in the MAXENT model were kept as the default settings.
I used six climatic predictors important for plant growth in the western U.S.: each of four quarterly precipitation variables, minimum winter temperature, and maximum summer temperature. I used PRISM climate data (Daly et al. 2002) at 2.5 arc minute resolution and resampled the grids to a 4 km resolution Albers equal area projection (which matches the resolution of the expert-identified datasets). In order to turn the continuous model results into discrete invasion risk maps, I selected thresholds of MAXENT suitability (‘gain’) and Mahalanobis distance that resulted in maps encompassing 95 % of the input occurrence or abundance dataset.
I compared the total area identified as suitable for each of the four models (based on herbarium, regional management, expert occurrence and expert abundance). I assumed that the suitability model based on abundance (hereafter abundance niche) represented risk of invasive plant abundance. Based on this assumption, I calculated false positive (predicted risk outside of the abundance niche) and false negative (predicted no-risk inside of the abundance niche) rates for the HSMs based on herbarium, regional management and expert-identified occurrences. Both false positives and false negatives were calculated relative to total available land area within the 15 state study area.
Results
Comparing point occurrence locations to abundance
Herbarium occurrence records were most likely to fall on areas where the species was not detected in the abundance dataset. There were a total of 1,640 overlapping herbarium records for all of the nine species when duplicates on abundance grid cells were excluded. Across all the invasive species tested, 60 % of presence locations from herbarium records were located in areas designated as ‘not detected’ by weed experts (Fig. 1). The percentage of herbarium occurrences within designated absences ranged from 25 % (for B. tectorum) to 80 % (for A. repens), and for four of the nine species herbarium occurrences fell on designated absences more than 70 % of the time. Meanwhile, averaged across all species, only 26 % of herbarium occurrences were located in areas designated as high abundance.

Comparison of percent of point occurrences from herbarium records (black) and regional databases (white) falling in areas where the species was not detected (absent), present at low abundance, and present at high abundance by county weed managers. Point occurrences from herbarium records are strongly biased towards locations where the species was not detected. For all but two of the above invasive plant species, a strong majority of herbarium occurrence points fell within areas where the species was not detected by weed experts. Regional occurrences were located in areas where the species was not detected half as often as herbarium occurrences
Regional management occurrence records were more likely than the herbarium records to fall on areas identified as high abundance. There were a total of 26,309 overlapping regional records for all of the nine species when duplicates on abundance grid cells were excluded. Averaged across all the invasive species tested, 32 % of presence locations from regional records were located in areas designated as ‘not detected’ by weed experts (Fig. 1). The percentage of regional occurrence points within designated absences ranged from 2 % (for B. tectorum) to 49 % (for T. caput-medusae). Averaged across all species, 42 % of regional occurrences were located in areas designated as high abundance.
Comparing modeled invasion risk
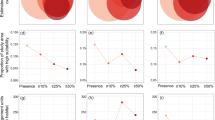
HSMs based on expert-identified occurrences or based on herbarium records predicted much more land area at risk of invasion than models based on regional occurrences or points with high abundance (Fig. 2). This trend was consistent across 8 of the 9 species. The only exception was T. caput-medusae which had only 114 total herbarium records clustered in the Pacific northwest, likely due to biased sampling. Models based on expert-identified occurrences had 1.31 and 1.44 times more land area than models based on expert-identified abundance using MD and MAXENT respectively. Similarly, models based on herbarium occurrences averages 1.36 and 1.75 (MD, MAXENT) times the land area of models based on abundance. Herbarium records produced models of invasion risk that were much more extensive than the abundance niche despite having only a small fraction of the total number of point locations. However, suitability models based on regional management occurrences were smaller than the abundance niche, averaging 0.69 and 0.98 (MD, MAXENT) times the land area (Fig. 2).

Estimated land area of suitability based on a MAXENT and b MD models. Distribution datasets are based on expert-identified abundance (grey with hash), regional management occurrences (white), expert-identified occurrences (grey), and herbarium occurrences (black)
Treating the abundance niche as ‘true’ abundance risk to evaluate model performance, models based on expert-identified and herbarium occurrences vastly overestimated suitability for abundance. Averaged across all species, 22–30 % (MD, MAXENT) of the area modeled as suitable based on expert-identified occurrences falls outside the abundance niche (Fig. 3a, b). Similarly, averaged across all species, 27–46 % (MD, MAXENT) of the area modeled as suitable based on herbarium records falls outside the abundance niche (Fig. 3a, b). On the other hand, models based on expert-identified and herbarium occurrences captured most of the abundance niche, both missing less than 10 % of land area using either MD or MAXENT (Fig. 3c, d). As a result, the average model based on expert-identified occurrences had an overall accuracy of 80–83 % (MD, MAXENT), while the models based on herbarium occurrences had an overall accuracy of 66–71 % (MAXENT, MD) relative to the abundance niche.

Assessment of suitability map accuracy relative to the abundance niche for models based on regional occurrences (white), expert-identified occurrences (grey) and herbarium occurrences (black). a False positive rates (pixels identified as suitable that were unsuitable for abundance) for MAXENT models. b False positive rates for MD models. c False negative rates (pixels identified as unsuitable that were suitable for abundance) for MAXENT models. d False negative rates for MD models
In contrast, HSMs based on regional occurrences had lower false positive rates. Averaged across all species, 3–30 % (MD, MAXENT) of the area modeled as suitable by regional data falls outside the abundance niche (Fig. 3a, b). However, models based on regional occurrences failed to capture an average of 21–38 % (MAXENT, MD) of the abundance niche (Fig. 3c, d). The average model based on herbarium occurrences had an overall accuracy of 76–79 % (MAXENT, MD) for predicting abundance risk.
Discussion
Modeling whether an invasive plant could establish in a given location is useful information. Indeed, climate matching as a tool for identifying potential invasive species (e.g., Thuiller et al. 2005) forms a cornerstone of successful import risk assessments (e.g., Pheloung et al. 1999). However, once a non-native species has arrived and become a transformative invader (sensu Richardson et al. 2000), modeling its potential for establishment (establishment niche) is much less relevant than modeling is potential for impact (impact niche; Leibold 1995), and the impact niche is better approximated by invasive species abundance than by presence (Ricciardi 2003; McDonald et al. 2009; Kulhanek et al. 2011). HSMs based on abundance records, such as the expert dataset used here, will provide better estimates of current and future impact risk.
Herbarium occurrences are most likely to be found in areas where weed managers have not detected the species (Fig. 1). This does not necessarily mean that the herbarium occurrences are wrong, although errors leading to range over-estimation have been identified in herbarium records (Miller et al. 2007). It may be that the species was once there, but did not persist. Or, it may be that weed managers have not yet discovered the species, which is plausible given the mapped grid resolution of 4 × 4 km. Either way, a species that has not been detected by invasive plant managers is unlikely to be a concern and therefore unlikely to be having a substantial impact. This skew towards low abundance locations is consistent with a goal of collecting rare species for herbarium collections, however it is ultimately problematic for modeling invasion risk.
Regional occurrences were more likely than herbarium records to be found in areas of high abundance (Fig. 1). However, 58 % of points were still located where the species is not detected or achieves only low abundance. This finding is consistent with Marvin et al. (2009) who found that regional occurrence points in the southeast U.S. were more likely to be found in locations with low abundance. Marvin et al. (2009) hypothesized that this could be due to a management focus on collecting points for early detection and rapid response, which focuses on small infestations in their early stages. This practice may also bias occurrences collected in the western U.S. towards lower abundance. However, it is clear that regional occurrences are more likely to be found in locations where the species is abundant than are herbarium records (Fig. 1).
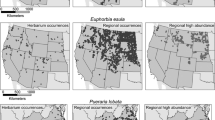
Because herbarium occurrences are so often located in areas where the species is of minimal concern, corresponding HSMs overestimate potential impact. In HSMs based on herbarium records, a third to nearly half of the projected area at risk was outside of the ‘impact niche’ modeled by the abundance data (Fig. 3a, b). Example MAXENT models for perennial pepperweed (L. latifolium) and hound’s tongue (C. officinale), which are the two species most closely matching the average pattern, are shown in Fig. 4 for illustration. Further, the herbarium occurrence points selected were only those located within and applied to the western U.S. If the national or global distribution had been used instead, the modeled establishment niche would have been much more extensive, while the impact niche would have changed only marginally because these species are known to have high impact primarily within the western U.S. Many recent HSM-based studies of invasion risk have relied on herbarium occurrence records assembled from a variety of global herbaria (Table 1). These models are correctly interpreted as predicting risk of establishment, but they cannot be considered accurate predictions of abundance or impact.

Comparison of distribution data and resulting HSMs for two example species. The top row shows high abundance points and modeled abundance niche. The middle row shows regional and herbarium points and modeled establishment niche. The bottom row shows a comparison between abundance and establishment niche for the different data sources
Modeled suitability for the expert-identified occurrences was more similar in land area to the herbarium records than to the abundance data (Fig. 2) and showed high false positive rates relative to the abundance niche (Fig. 3a, b). This finding suggests that the differences presented here between the abundance niche and herbarium-based establishment niche are not simply an artifact of biased collection of the abundance data. Rather, the sampling effort (represented by the expert-identified occurrences) encompassed nearly the same range of climatic space represented by the herbarium records. However, the expert-identified occurrence data do not encompass the entire establishment niche as defined by the herbarium records, so it is likely that the true abundance niches for these species are slightly larger than the ones modeled here because the range has not been completely sampled.
HSMs based on regional occurrences were more likely to under-estimate rather than over-estimate invasion risk, failing to identify, on average, 21–38 % of the abundance niche (Fig. 3c, d). In this example, the under-estimation of regional occurrences is likely due to a concentration of compiled data in 3 states (Oregon, Nevada, and Idaho), which represents a sampling bias that may be common to regional datasets if some areas do not collect or compile invasive species distribution data. Even with bias corrections applied to the models, biased distribution data are known to create biased suitability model results (Beaumont et al. 2009b; Wolmarans et al. 2010), and for invasive species are likely to underestimate risk. Models of invasion risk based on regional occurrences (Table 1) can therefore be interpreted as an underestimate of impact risk (capturing 62–79 % of the abundance niche), and an extreme underestimate of establishment risk.
Abundance data are rarely collected for any species, let alone invasive species, and even more rarely at regional scales appropriate for distribution modeling. Percent cover or biomass is time consuming to measure, and visual estimates of cover are often unreliable. However, even qualitative rankings of abundance would be a huge boon for predictive modeling efforts. The difference between a single roadside weed and a landscape of near-monoculture is enormous, but that distinction is not made in presence only datasets. I urge the scientific and management communities to collect invasive species abundance data, and to make existing data publicly available through online archives (e.g., EDDMaps.org; Simpson et al. 2009).
In the absence of abundance data, suitability modelers can be more vigilant in searching for regional distribution data to supplement herbarium records. Regional datasets are rarely centrally located, but are collected by many local agencies (Crall et al. 2006). By creating two HSMs, one based on herbarium records, and one based on regional records, modelers can be more explicit about how they define invasion risk. The smaller area identified as suitable by regional records is likely to underestimate impact risk, while the larger area identified as suitable by herbarium records is likely to model establishment risk, but vastly overestimate impact. This level of transparency would improve the translation of predictive models into invasive species management.
Estimates of false positive and false negative rates in HSMs derived from regional and herbarium occurrences assume that the projected abundance niche represents true potential for invasive plant abundance, and therefore impact. This assumption is almost certainly flawed. All of these species continue to expand, hence estimates of both abundance niche and occurrence niche are likely underestimates. Time lags between introduction and impact of non-native species are well known (Mack et al. 2000; Essl et al. 2011) and HSMs based on data collected at early stages of invasion will underestimate potential distribution (Václavík and Meentemeyer 2012). However, all of these species have existed in the U.S. for decades (most arrived in the 19th or early 20th century) and were widely introduced through contaminated seed and hay crops. Welk (2004) estimated that the geographic area of modeled suitability stabilized after 100–150 years of invasion, comparable to the time since introduction for most of the species presented here. While many of these species may continue to expand locally, at regional scales it is likely that they encompass a majority of suitable climate space.
How worried should we be about overestimating impact? In a perfect world, managers would have all the resources necessary to eradicate non-native species. In reality, limited staff and finances force constant prioritization of target species and locations. HSMs are increasingly being used to inform invasive species management (e.g., Stohlgren et al. 2010; CAL-IPC 2012). Yet, it is clear from these results that occurrence data are a poor proxy for abundance. By relying on models of occurrence risk to inform treatment plans, managers risk wasting resources in areas where target species are unlikely to ever have a substantial impact. The invasive species modeling community must move towards modeling abundance, and thereby impact, in order to become more management relevant.
Notes
Biodiversity occurrence data provided by: Arizona State University Vascular Plant Herbarium, Botany Vascular Plant Collection, CSU Herbarium, Consortium of California Herbaria, Herbarium GZU, Herbarium of The New York Botanical Garden, Missouri Botanical Garden, New Mexico Biodiversity Collections Consortium database, UA Herbarium, USDA PLANTS Database, USU-UTC Specimen Database, University and Jepson Herbaria DiGIR provider, Utah Valley State College Herbarium, Vascular Plant Collection, Vascular Plant Collection—University of Washington Herbarium (WTU) (Accessed through GBIF Data Portal, data.gbif.org, 2010-09-20).
References
Albright TP, Anderson DP, Keuler NS, Pearson SM, Turner MG (2009) The spatial legacy of introduction: Celastrus orbiculatus in the southern Appalachians, USA. J Appl Ecol 46(6):1229–1238. doi:10.1111/j.1365-2664.2009.01707.x
Albright TP, Chen H, Chen LJ, Guo QF (2010) The ecological niche and reciprocal prediction of the disjunct distribution of an invasive species: the example of Ailanthus altissima. Biol Invasions 12(8):2413–2427. doi:10.1007/s10530-009-9652-8
Barney JN, Whitlow TH, Lembo AJ (2008) Revealing historic invasion patterns and potential invasion sites for two non-native plant species. PLoS ONE 3(2):e1635. doi:10.1371/journal.pone.0001635
Beaumont LJ, Gallagher RV, Downey PO, Thuiller W, Leishman MR, Hughes L (2009a) Modelling the impact of Hieracium spp. on protected areas in Australia under future climates. Ecography 32(5):757–764. doi:10.1111/j.1600-0587.2009.05705.x
Beaumont LJ, Gallagher RV, Thuiller W, Downey PO, Leishman MR, Hughes L (2009b) Different climatic envelopes among invasive populations may lead to underestimations of current and future biological invasions. Divers Distrib 15(3):409–420. doi:10.1111/j.1472-4642.2008.00547.x
Bradley BA (2009) Regional analysis of impacts of climate change on cheatgrass invasion shows potential risk and opportunity. Glob Change Biol 15(1):196–208
Bradley BA, Marvin DC (2011) Using expert knowledge to satisfy data needs: mapping invasive plant distributions in the Western U.S. West N. Am Nat 71(3):302–315
Bradley BA, Oppenheimer M, Wilcove DS (2009) Climate change and plant invasion: restoration opportunities ahead? Glob Change Biol 15(6):1511–1521
Bradley BA, Wilcove DS, Oppenheimer M (2010) Climate change increases risk of plant invasion in the Eastern United States. Biol Invasions 12(6):1855–1872. doi:10.1007/s10530-009-9597-y
Brewer JS (2011) Per capita community-level effects of an invasive grass, Microstegium vimineum, on vegetation in mesic forests in northern Mississippi (USA). Biol Invasions 13(3):701–715. doi:10.1007/s10530-010-9861-1
Brooks ML, D’Antonio CM, Richardson DM, Grace JB, Keeley JE, DiTomaso JM, Hobbs RJ, Pellant M, Pyke D (2004) Effects of invasive alien plants on fire regimes. Bioscience 54(7):677–688. doi:10.1641/0006-3568(2004)054[0677:eoiapo]2.0.co;2
CAL-IPC (2012) Statewide risk mapping for early detection. California Invasive Plant Council. http://www.cal-ipc.org/ip/mapping/statewide_maps/index.php. Accessed 16 Jan 2012
Chejara VK, Kriticos DJ, Kristiansen P, Sindel BM, Whalley RDB, Nadolny C (2010) The current and future potential geographical distribution of Hyparrhenia hirta. Weed Res 50(2):174–184. doi:10.1111/j.1365-3180.2010.00765.x
Christenhusz MJM, Toivonen TK (2008) Giants invading the tropics: the oriental vessel fern, Angiopteris evecta (Marattiaceae). Biol Invasions 10(8):1215–1228. doi:10.1007/s10530-007-9197-7
Crall AW, Meyerson LA, Stohlgren TJ, Jarnevich CS, Newman GJ, Graham J (2006) Show me the numbers: what data currently exist for non-native species in the USA? Front Ecol Environ 4(8):414–418
Daly C, Gibson WP, Taylor GH, Johnson GL, Pasteris P (2002) A knowledge-based approach to the statistical mapping of climate. Clim Res 22:99–113
D’Antonio CM, Vitousek PM (1992) Biological invasions by exotic grasses, the grass fire cycle, and global change. Annu Rev Ecol Syst 23:63–87
Dunlop EA, Wilson JC, Mackey AP (2006) The potential geographic distribution of the invasive weed Senna obtusifolia in Australia. Weed Res 46(5):404–413
Essl F, Dullinger S, Rabitsch W, Hulme PE, Hulber K, Jarosik V, Kleinbauer I, Krausmann F, Kuhn I, Nentwig W, Vila M, Genovesi P, Gherardi F, Desprez-Loustau ML, Roques A, Pysek P (2011) Socioeconomic legacy yields an invasion debt. Proc Natl Acad Sci USA 108(1):203–207. doi:10.1073/pnas.1011728108
Estes LD, Bradley BA, Beukes H, Hole DG, Lau M, Oppenheimer M, Schulze R, Tadross MA, Turner WR (2012) Comparing mechanistic and empirical model projections of crop suitability and productivity: implications for projecting ecological change. Princeton, NJ
Follak S, Strauss G (2010) Potential distribution and management of the invasive weed Solanum carolinense in Central Europe. Weed Res 50(6):544–552. doi:10.1111/j.1365-3180.2010.00802.x
Gasso N, Sol D, Pino J, Dana ED, Lloret F, Sanz-Elorza M, Sobrino E, Vila M (2009) Exploring species attributes and site characteristics to assess plant invasions in Spain. Divers Distrib 15(1):50–58. doi:10.1111/j.1472-4642.2008.00501.x
Hulme PE (2006) Beyond control: wider implications for the management of biological invasions. J Appl Ecol 43(5):835–847. doi:10.1111/j.1365-2664.2006.01227.x
Ibanez I, Silander JA, Wilson AM, Lafleur N, Tanaka N, Tsuyama I (2009) Multivariate forecasts of potential distributions of invasive plant species. Ecol Appl 19(2):359–375
Jarnevich CS, Reynolds LV (2011) Challenges of predicting the potential distribution of a slow-spreading invader: a habitat suitability map for an invasive riparian tree. Biol Invasions 13(1):153–163. doi:10.1007/s10530-010-9798-4
Jarnevich CS, Stohlgren TJ (2009) Near term climate projections for invasive species distributions. Biol Invasions 11:1373–1379
Jiménez-Valverde A, Lobo JM, Hortal J (2008) Not as good as they seem: the importance of concepts in species distribution modelling. Divers Distrib 14(6):885–890. doi:10.1111/j.1472-4642.2008.00496.x
Jiménez-Valverde A, Diniz F, de Azevedo EB, Borges PAV (2009) Species distribution models do not account for abundance: the case of arthropods on Terceira Island. Ann Zool Fenn 46(6):451–464
Kriticos DJ, Sutherst RW, Brown JR, Adkins SW, Maywald GF (2003) Climate change and the potential distribution of an invasive alien plant: Acacia nilotica ssp indica in Australia. J Appl Ecol 40(1):111–124
Kriticos DJ, Watt MS, Potter KJB, Manning LK, Alexander NS, Tallent-Halsell N (2011) Managing invasive weeds under climate change: considering the current and potential future distribution of Buddleja davidii. Weed Res 51(1):85–96. doi:10.1111/j.1365-3180.2010.00827.x
Kulhanek SA, Leung B, Ricciardi A (2011) Using ecological niche models to predict the abundance and impact of invasive species: application to the common carp. Ecol Appl 21(1):203–213
Le Maitre DC, Thuiller W, Schonegevel L (2008) Developing an approach to defining the potential distributions of invasive plant species: a case study of Hakea species in South Africa. Glob Ecol Biogeogr 17(5):569–584. doi:10.1111/j.1466-8238.2008.00407.x
Leibold MA (1995) The niche concept revisited—mechanistic models and community context. Ecology 76(5):1371–1382
Lobo JM, Jiménez-Valverde A, Hortal J (2010) The uncertain nature of absences and their importance in species distribution modelling. Ecography 33(1):103–114. doi:10.1111/j.1600-0587.2009.06039.x
Lockwood J, Hoopes M, Marchetti M (2007) Invasion ecology. Blackwell Publishing, Malden, MA
Loo SE, Mac Nally R, Lake PS (2007) Forecasting New Zealand mudsnail invasion range: model comparisons using native and invaded ranges. Ecol Appl 17(1):181–189
Mack RN, Simberloff D, Lonsdale WM, Evans H, Clout M, Bazzaz FA (2000) Biotic invasions: causes, epidemiology, global consequences, and control. Ecol Appl 10(3):689–710
Marvin DC, Bradley BA, Wilcove DS (2009) A novel, web-based ecosystem mapping tool using expert opinion. Nat Areas J 29(3):281–292
McDonald A, Riha S, DiTommaso A, DeGaetano A (2009) Climate change and the geography of weed damage: analysis of US maize systems suggests the potential for significant range transformations. Agric Ecosyst Environ 130(3–4):131–140. doi:10.1016/j.agee.2008.12.007
Miller BP, Enright NJ, Lamont BB (2007) Record error and range contraction, real and imagined, in the restricted shrub Banksia hookeriana in south-western Australia. Divers Distrib 13(4):406–417. doi:10.1111/j.1472-4642.2007.00348.x
Nielsen SE, Johnson CJ, Heard DC, Boyce MS (2005) Can models of presence-absence be used to scale abundance?—two case studies considering extremes in life history. Ecography 28(2):197–208
Nielsen C, Hartvig P, Kollmann J (2008) Predicting the distribution of the invasive alien Heracleum mantegazzianum at two different spatial scales. Divers Distrib 14(2):307–317. doi:10.1111/j.1472-4642.2007.00456.x
Parker IM, Simberloff D, Lonsdale WM, Goodell K, Wonham M, Kareiva PM, Williamson MH, Von Holle B, Moyle PB, Byers JE, Goldwasser L (1999) Impact: toward a framework for understanding the ecological effects of invaders. Biol Invasions 1(1):3–19. doi:10.1023/a:1010034312781
Pearce J, Ferrier S (2001) The practical value of modelling relative abundance of species for regional conservation planning: a case study. Biol Conserv 98(1):33–43
Peterson AT, Vieglais DA (2001) Predicting species invasions using ecological niche modeling: new approaches from bioinformatics attack a pressing problem. Bioscience 51(5):363–371
Peterson AT, Stewart A, Mohamed KI, Araujo MB (2008) Shifting global invasive potential of European plants with climate change. PLoS ONE 3(6):e2441
Pheloung PC, Williams PA, Halloy SR (1999) A weed risk assessment model for use as a biosecurity tool evaluating plant introductions. J Environ Manag 57(4):239–251
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecol Model 190(3–4):231–259
Pimentel D, Zuniga R, Morrison D (2005) Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol Econ 52(3):273–288
Ramcharan CW, Padilla DK, Dodson SI (1992) Models to predict potential occurrence and density of the zebra mussel, Dreissena-polymorpha. Can J Fish Aquat Sci 49(12):2611–2620
Ricciardi A (2003) Predicting the impacts of an introduced species from its invasion history: an empirical approach applied to Zebra mussel invasions. Freshw Biol 48(6):972–981
Richardson DM, Pysek P, Rejmanek M, Barbour MG, Panetta FD, West CJ (2000) Naturalization and invasion of alien plants: concepts and definitions. Divers Distrib 6(2):93–107
Richardson DM, Iponga DM, Roura-Pascual N, Krug RM, Milton SJ, Hughes GO, Thuiller W (2010) Accommodating scenarios of climate change and management in modelling the distribution of the invasive tree Schinus molle in South Africa. Ecography 33(6):1049–1061. doi:10.1111/j.1600-0587.2010.06350.x
Simberloff D (1985) Predicting ecological effects of novel entities: evidence from higher organisms. In: Halvorson HO, Pramer D, Rogul M (eds) Engineered organisms in the environment/scientific issues. American Society for Microbiology, Washington, DC, pp 152–161
Simpson A, Jarnevich C, Madsen J, Westbrooks R, Fournier C, Mehrhoff L, Browne M, Graham J, Sellers E (2009) Invasive species information networks: collaboration at multiple scales for prevention, early detection and rapid response to invasive alien species. Biodiversity 10(2&3):5–13
Smolik MG, Dullinger S, Essl F, Kleinbauer I, Leitner M, Peterseil J, Stadler LM, Vogl G (2010) Integrating species distribution models and interacting particle systems to predict the spread of an invasive alien plant. J Biogeogr 37(3):411–422. doi:10.1111/j.1365-2699.2009.02227.x
Stohlgren TJ, Ma P, Kumar S, Rocca M, Morisette JT, Jarnevich CS, Benson N (2010) Ensemble habitat mapping of invasive plant species. Risk Anal 30(2):224–235. doi:10.1111/j.1539-6924.2009.01343.x
Thuiller W, Richardson DM, Pysek P, Midgley GF, Hughes GO, Rouget M (2005) Niche-based modelling as a tool for predicting the risk of alien plant invasions at a global scale. Glob Change Biol 11(12):2234–2250
Tôrres NM, De Marco P, Santos T, Silveira L, de Almeida Jácomo AT, Diniz-Filho JAF (2012) Can species distribution modelling provide estimates of population densities? A case study with jaguars in the Neotropics. Divers Distrib. doi:10.1111/j.1472-4642.2012.00892.x
Tsoar A, Allouche O, Steinitz O, Rotem D, Kadmon R (2007) A comparative evaluation of presence-only methods for modelling species distribution. Divers Distrib 13:397–405
Václavík T, Meentemeyer RK (2012) Equilibrium or not? Modelling potential distribution of invasive species in different stages of invasion. Divers Distrib 18(1):73–83. doi:10.1111/j.1472-4642.2011.00854.x
van Klinken RD, Lawson BE, Zalucki MP (2009) Predicting invasions in Australia by a Neotropical shrub under climate change: the challenge of novel climates and parameter estimation. Glob Ecol Biogeogr 18(6):688–700. doi:10.1111/j.1466-8238.2009.00483.x
VanDerWal J, Shoo LP, Johnson CN, Williams SE (2009) Abundance and the environmental niche: environmental suitability estimated from niche models predicts the upper limit of local abundance. Am Nat 174(2):282–291. doi:10.1086/600087
Vilà M, Espinar JL, Hejda M, Hulme PE, Jarošík V, Maron JL, Pergl J, Schaffner U, Sun Y, Pyšek P (2011) Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities and ecosystems. Ecol Lett 14(7):702–708. doi:10.1111/j.1461-0248.2011.01628.x
Watt MS, Kriticos DJ, Manning LK (2009) The current and future potential distribution of Melaleuca quinquenervia. Weed Res 49(4):381–390. doi:10.1111/j.1365-3180.2009.00704.x
Welk E (2004) Constraints in range predictions of invasive plant species due to non-equilibrium distribution patterns: Purple loosestrife (Lythrum salicaria) in North America. Ecol Model 179(4):551–567
Welk E, Schubert K, Hoffmann MH (2002) Present and potential distribution of invasive garlic mustard (Alliaria petiolata) in North America. Divers Distrib 8(4):219–233
Wilcove DS, Rothstein D, Dubow J, Phillips A, Losos E (1998) Quantifying threats to imperiled species in the United States. Bioscience 48(8):607–615
Wolmarans R, Robertson MP, van Rensburg BJ (2010) Predicting invasive alien plant distributions: how geographical bias in occurrence records influences model performance. J Biogeogr 37(9):1797–1810. doi:10.1111/j.1365-2699.2010.02325.x
Acknowledgments
Abundance estimates for invasive plants in the western U.S. would not have been possible without the participation of hundreds of weed managers in a regional mapping effort. My thanks to all who participated. This research was partially supported by the Department of Defense through the Strategic Environmental Research and Development Program (SERDP) grant number RC-1722.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Bradley, B.A. Distribution models of invasive plants over-estimate potential impact. Biol Invasions 15, 1417–1429 (2013). https://doi.org/10.1007/s10530-012-0380-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-012-0380-0