
Abstract
Introduced mammalian predators have had significant impacts on many native prey species. Although control of such predators for conservation management is becoming increasingly commonplace, it is often undertaken at a relatively small scale in relation to the overall predator population. Processes such as immigration mean that it remains difficult to determine the effectiveness of control measures. We investigated the impacts of feral ferret Mustela furo removal on the entire feral ferret population on Rathlin Island, UK. Removal of ferrets prior to breeding led to a substantial increase in the post-dispersal population through the enhanced survival of juveniles. Despite increased numbers, overwinter survival remained high, potentially aided by the reduced territoriality shown by this feral species compared to wild carnivores. The response of this ferret population to control is a further illustration of the complex ecological processes and outcomes arising from the anthropogenic disruption of wildlife populations. It highlights how partial or localised management may prove ineffective, and at worst might exacerbate the problems that management was designed to avert.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Invasive non-native predators are a major threat to the survival of many native species (Veitch and Clout 2002); a situation which is often compounded by habitat loss and fragmentation, and other human activities (Vitousek et al. 1997). When such alterations in land use benefit exotic species, many of which are omnivores or generalist predators (Findlay and Houlahan 1997; McKinney and Lockwood 1999; Lockwood et al. 2007), then intensive management of predators may be necessary in order to conserve vulnerable native species (Veitch and Clout 2002).
The eradication of whole populations of an introduced predator may be the desirable option, and such attempts are becoming both more frequent and more ambitious (Veitch and Clout 2002; Towns and Broome 2003; Morrison et al. 2007; Parkes and Panetta 2009). This approach is not always suitable, however, whether because of the large ranges of well-established introduced species (Macdonald and Harrington 2003), a lack of adequate funds or because of political or public opposition (Bertolino and Genovesi 2003; Bremner and Park 2007). In such instances, targeted predator control that is limited to important sites, such as around colonies of breeding birds, is often a more acceptable option (Greenwood et al. 1990; Keedwell et al. 2002; Bolton et al. 2007). However, such an approach to management then creates a control zone within a wider landscape matrix that continues to support predators (Janzen 1986). Without a permanent barrier, immigration is inevitable and many predatory species are capable of dispersing over long distances (Byrom 2002; King and Powell 2006). This means that continuous monitoring and at least episodic control is typically required. In some cases this can result in little or no effect on overall predator populations, and so predator control under these circumstances can often appear a Sisyphean task (Greentree et al. 2000).
Nevertheless, because predation can have significant impacts on small prey populations (Sinclair et al. 1998), control of both invasive and native predators is regularly undertaken (Dowding and Murphy 2001; Kinnear et al. 2002; Macdonald and Harrington 2003). Thus an understanding of its effectiveness in controlling predator populations, and in generating conservation benefits, is essential (Smith et al. 2010). For example, limited control may not be appropriate with regards to the spatial scale at which the predator operates (Lahti 2001); and many predator populations have social and/or spatial structure, and culling can disrupt these systems with unpredictable results (Woodroffe et al. 2006; McDonald et al. 2008). The design and implementation of control programmes therefore needs to be carefully considered in order to avoid or mitigate against such outcomes. In addition, evidence about what contributes to a successful control operation is often lacking (Sutherland et al. 2004; King et al. 2009), so the examination of trial control programmes provides such an opportunity.
In this study, we examined the impact of predator control on an introduced non-native mustelid, the feral ferret Mustela furo. Members of the Mustelidae have been deliberately and accidentally introduced to a number of countries and islands (Long 2003; Clapperton and Byrom 2005; Bonesi and Palazon 2007), where they have usually adapted well and become major predators of native species (Macdonald and Harrington 2003; King and Powell 2006). However, outside of New Zealand (Clapperton and Byrom 2005), the impact of introduced ferrets, although presumed to be negative, is essentially unknown. On our study island, for example, declines and changes in the distribution of ground and burrow nesting birds have been observed since the introduction of ferrets, despite positive habitat management for these species (Anonymous 2007, L. McFaul, unpublished data), but no contemporaneous assessment of ferret impact was conducted. When the impact of predation by ferrets has been detrimental, removing them has often been considered necessary (Clapperton and Byrom 2005), but the effect of such removal programmes on ferret populations is also unknown. We examined population level responses to control measures that were designed to represent predator control for the protection of key nesting areas for ground-nesting birds. We present a case whereby removal over such limited areas can make subsequent control efforts harder, and may give rise to a greater threat to prey populations that the management action was intended to protect.
Materials and methods
Study site
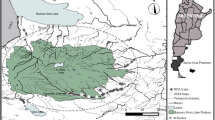
Fieldwork was carried out on Rathlin Island (55º18′N 06º13′W; 1,525 ha), located off the north-east coast of Northern Ireland (Fig. 1). The vegetation is dominated by maritime heath and acid grassland, with small flushes, wetlands and ponds. Most land is grazed by cattle and sheep at low densities, and the island has a year round population of approximately 80 people. The island supports internationally important populations of breeding seabirds (Mitchell et al. 2004), and is designated for protection under a number of conservation instruments. Ferrets were intentionally released to Rathlin over several years in the 1980s in an attempt at biological control of introduced rabbits Oryctolagus cuniculus (L. McFaul pers comm.).

Map of Rathlin Island study site showing randomly assigned ferret removal areas. Both T1 areas were trapped in 2006, both T2 areas were trapped in 2007, and neither T3 area was trapped in either year. Shaded areas represent buffer zones between treatments. Inset shows Rathlin in relation to Ireland and the UK
Population assessment
94 single entry mink cage traps (Solway Feeders Ltd.) were placed throughout the island on accessible land in an approximate grid system at 300–400 m spacing. Traps were placed in open-ended wooden tunnels to provide protection from the elements for trapped animals, as well as potentially making them more attractive to ferrets. They were baited with preserved fish, and were sited where animals were most likely to be moving naturally, such as alongside dry stone walls and other linear features. Trap sites remained the same between years. In 2005–2006 traps were operated in a rolling front of three groups of 32, 32 and 30 traps, effectively splitting the island into three sections. Traps were set for six consecutive nights, in November–December and again in February. In 2006–2007, with improved logistics and increased experience, all 94 traps were run for five successive nights per month from October–January inclusive. All traps were checked daily and rebaited if necessary. All captured ferrets were marked with individually numbered tags (1005–1003, National Band & Tag Co.) in both ears.
Radio collaring and tracking
In both years, a sample of ferrets spread throughout the entire island were fitted with radio collars. Before handling, ferrets were anaesthetised using isofluorane (Abbott Laboratories Ltd.). Mortality-sensing collars (11.5 g, TW-5, Biotrack Ltd.), representing a maximum of 2.5% of body mass, were fitted. All individuals were checked to ensure recovery was complete before being released within a few minutes of the initial procedure into a burrow or under thick vegetation close to the trapping site.
In 2005–2006, 18 individuals (nine male, nine female) were radio collared. One collar failed shortly after deployment, and two individuals died during the tracking period. In 2006–2007, 13 individuals (six males, seven females) were collared, with three individuals dying during this period. Animals were radiotracked on foot from the day after collaring through to early March to denning or activity spots, both during the day and night, using a 3 element Yagi antenna with either a TR-5 (Telonics) or a Mariner (Biotrack) receiver. Location co-ordinates were recorded using a handheld GPS unit (Garmin GPS72) with an accuracy of 5–10 m dependent on weather conditions, and checked on a topographic map. Intervals between tracking days were irregular, although they were typically less than 3 days and never more than seven.
Ferret control
The island was divided into six approximately equally sized treatment blocks (mean 151 ha, range 141–160 ha Fig. 1). The number of blocks was chosen as a balance between ensuring a degree of replication within the design while accommodating landowners and working with biologically meaningful space (Oksanen 2001). The size of a block was based on the results of the radio telemetry conducted over the winter of 2005–2006, and represents an area larger than the mean male ferret home range calculated from this study. Blocks were separated by buffer zones that were a minimum of 300 m across, a distance greater than the mean distance moved by male ferrets between consecutive fixes. Ferrets were removed from two randomly selected blocks in year 1 and from two different blocks in year 2, and were never removed from the final two blocks (Fig. 1). Thus both removal and non-removal treatments were replicated within each year, and the experimental removal treatment occurred as a crossover design between years. Clearly the confines of the island produce natural limitations to the degree to which treatment areas can be separated and replicated. However, ferrets held home ranges, and so while some of the male ranges crossed buffer zones (Fig. 2), the majority of animals were not found in multiple treatment areas. This was also found when marked individuals captured during population monitoring were captured in the same areas during removal treatments. This meant that we would be comparing removal areas (where ferrets were virtually absent) to non-removal areas (with a slightly reduced number of ferrets). In addition, we were not attempting to compare results from removal and non-removal areas; instead we were taking advantage of our understanding of the entire population to examine the effect of localised culling on population responses. Traps within removal areas were set using the same methods as for population monitoring, and were run immediately prior to and then throughout the bird nesting season (which commenced slightly earlier in 2007) from 10 April-15 July 2006, and 25 March-15 July 2007. Absence of ferrets from traps was corroborated with data from unbaited tracking tunnels (King et al. 1994) run monthly in all treatment blocks. No ferrets were detected in removal zones, but ferret tracks were found in non-removal zones in all months, including the presence of juveniles at the expected time (T Bodey unpublished data). In removal areas, captured animals were first anaesthetised using isofluorane, and then euthanized with an intra-peritoneal injection of sodium pentobarbital (Euthatal, Merial Animal Health Ltd.).

Overwinter home ranges of radiocollared ferrets as 95% minimum convex polygons in relation to treatment areas (T1, T2, T3) and buffer zones (shaded grey) (see materials and methods and Fig. 1 for details) for a males 2005–2006 (n = 6) b females 2005–2006 (n = 9) c males 2006–2007 (n = 5) d females 2006–2007 (n = 5)
Ageing methods
Skulls of euthanized individuals were prepared following the methodology of McDonald and Vaughan (1999). A lower canine was then extracted and sent for cementum aging (Matson’s Laboratory LLC, USA), with all individuals ascribed a common birth date of 1 April.
Data analysis
For 2005–2006 with two trapping sweeps of the island, the Lincoln-Petersen unbiased estimator (Chapman 1951) was used to estimate population size, with an unbiased variance calculated as per Seber (1973). For 2006–2007, a comparative Lincoln-Peterson unbiased estimator was calculated using the information from the first and last trapping occasions only to replicate the 2005–2006 protocol. However, the four trapping sessions allowed calculation of a more robust estimate for this second study season using maximum likelihood methods in Program MARK version 5.0 (White and Burnham 1999) under the assumptions of a closed population. Prior analysis of the 2006–2007 recapture data as an open population with a standard Cormack-Jolly-Seber model and goodness of fit tests carried out in RELEASE suggested the assumptions were not violated, but also produced high monthly survival estimates, which supported known overwinter survival from telemetry data. Thus a closed population model was appropriate as demographic closure is all but complete, and the assumptions of geographic closure and no loss or misidentification of marks were met.
All measures of home range analysis have limitations, some of which vary with the behaviour of the study species. Minimum convex polygons (MCPs) are robust with smaller sample sizes, and are necessary for comparison with other studies of feral ferrets (Kenward 2001). As 100% MCPs are sensitive to outliers, 95% MCPs were also calculated, with results presented for the latter although patterns did not vary between metrics. All home ranges were plotted in Arcview 3.2, and bootstrapping of 1,000 iterations of points plotted against increasing area was used to check whether asymptotes were approached. Animals with too few data points, whether through collar failure or lack of observations, were excluded from analyses. All locations were included for each individual as observations were made a minimum of 15 h apart, a time well in excess of that required for an individual to move to any point within its home range (Rooney et al. 1998). The influence of year and sex and their interaction term were examined for both measures of home range using generalised linear models with a Gaussian error structure. Values were natural log transformed prior to analysis in order to meet assumptions of normality. Home range percentage overlaps were calculated between collared ferrets for each home range measure as the geometric mean of the product of the ratios of the area of overlap (areaαβ) to home range size of each individual (homerangeα and homerangeβ) in the pair (Minta 1993):
Results
Population size
In 2005–2006, 35 individuals were caught over 1,145 trap nights, with 66% recaptured at least once. This gave a catch per unit effort of 9.5 ferrets per 100 corrected trap nights. In 2006–2007, 62 individuals new for this trapping period were caught in 1907 trap nights, with 84% recaptured at least once. This produced a catch per unit effort of 11.6 ferrets per 100 corrected trap nights. For 2005–2006 the Chapman correction of the Lincoln-Peterson estimator ±sd provided an estimate of abundance of 42 ± 4. In 2006–2007 the same measure provided an estimate of abundance of 62 ± 7. Applying maximum likelihood estimation to the data from 2006–2007, a model incorporating behavioural and individual responses to trapping (BH) was the most favoured, with twice as much support as for the model considering individual heterogeneity alone (H). Similar models incorporating an additional time component (BHT and HT) received much less support (Table 1). Population estimates from the first two models were almost identical, and model averaging across all four models produced an abundance estimate of 65 ± 3. Both methods thus suggest the island’s total ferret population increased by approximately 50% following removal trapping across approximately one quarter of the island area.
Age structure
17 ferrets were removed in 2006 and 27 were removed in 2007 (Fig. 3), with ten more animals classed as young of the previous year in 2007. However there was no significant difference in capture rates between 1 year old and older ferrets between the 2 years (likelihood ratio 0.885, df = 1, P = 0.347). This was to be expected as different areas were trapped in the 2 years and so captured ferrets will not fully reflect any changes in population structure.

Age of ferrets captured during predator control. Ferrets are all given the common birth date of April 1 so, for example, 1 year olds were all captured prior to April 1 the year after they were born and thus span the range from 0–1 years
Home ranges
Fully explored home ranges were determined for 16 ferrets in 2005–2006 (six male, nine female) and for 10 individuals in 2006–2007 (five male, five female) (Fig. 2). Ferret home ranges were substantially greater for males (the larger sex) than females in both years, but the home ranges of both sexes were smaller in 2006–2007 than in 2005–2006 (Figs. 2, 4) (95% MCP mean ± SD (ha) 2005–2006: males 136 ± 78, females 35 ± 18; 2006–2007: males 54 ± 21, females 22 ± 7). Both sex and year factors were significant (95% MCP: Year F1,23 = 8.68, P = 0.007; Sex F1,23 = 33.96, P < 0.001), but their interaction was not and was not included in the final model (Fig. 4). Where collared individuals were close enough together that range overlap was possible, substantial shared areas were observed, with the extent of overlap being largely consistent between years and sex combinations (Table 2). Den sharing was observed through radio telemetry in both years (n = 23 occasions) and involved 20 different individuals.

Ferret home range size (ha) in different study years. Home ranges are plotted as natural log transformed 95% minimum convex polygons (MCPs) of male (black circles) and female (white circles) ferrets. 100% MCPs presented the same pattern
Discussion
Our results indicate a substantial between-year increase in ferret population size following control efforts; an effect that was likely to have been driven by an increase in juvenile survival and recruitment to the adult population in individuals born outside trapped blocks which were then able to move into this unoccupied space. While a small number of individuals were removed even though their home ranges only partly spanned a removal area, this effect was limited to a few male ferrets in 2005–2006 only (Fig. 2), and does not alter this main conclusion. A population increase has an obvious significance for the prey species that predator control is designed to protect, and demonstrates that the development of predator management strategies should not ignore factors linked to the surrounding areas and the ecology of the species being controlled.
Population changes and survival
Prior to manipulation, ferret numbers on Rathlin were not high, with a density of approximately 3 ferrets per km². Models of ferret population dynamics from New Zealand, have demonstrated the importance of density dependent limitation in population growth, principally through low juvenile recruitment (Barlow and Norbury 2001; Caley et al. 2002). Barlow and Norbury (2001) suggested a 50% reduction in population size would be required to ensure a significant decrease in ferret numbers the following year. In our study, natural mortality over winter was low, and most individuals caught over winter were recaptured in the spring. Removal trapping over only part of the island was successful in substantially reducing the population in 2006, with at least 43% of the total population removed prior to the emergence of young of the year. Thus, adult densities were reduced in the period immediately prior to juvenile recruitment, reducing any potential impact of this constraint. However, while the total removed was close to the value suggested by Barlow and Norbury (2001) to be necessary for sustained population reduction, the opposite effect was observed.
Removal trapping in the second year was not implemented until after any density dependent recruitment effects could have operated (King et al. 1994; Byrom 2002), so the observed increase clearly represents enhanced juvenile survival, particularly where adults have been removed (Byrom 2002). In our study, removal blocks can only have been fully repopulated through immigration of juveniles from non-removal blocks, resulting in substantial overcompensation in recruitment levels across the entire population (Sinclair 1996). Although mortality (through decreased prey availability and colder temperatures over winter for example) might then be anticipated to reduce the population, this was not apparent. Concurrent work on ferret diet (Bodey et al. 2010) revealed the potential for anthropogenic resources in the form of livestock carrion to sustain the ferret population even following decreases in their main lagomorph prey. However, it also demonstrated that the larger ferret population did not proportionately increase its consumption of this resource, indicating live prey levels were sufficient to support the great majority of new recruits. Thus the island was able to support a larger ferret population and, while this could be maintained as a result of current land management practices even if favoured lagomorph prey were depleted (Bodey et al. 2010), for how long remains unknown. However, clearly such fluctuations in predator numbers can have important ramifications for protective management of prey species in subsequent seasons (Courchamp et al. 2000). Even if predation of a species is only incidental, more animals will inevitably lead to more predation incidents, and for small populations of rare species, particularly those limited to topographical or habitat islands, the presence of such a predator can then be the difference between survival and extinction (Sinclair et al. 1998). In a mainland situation, such an increase in predator populations may not have greater deleterious impacts on localised prey though, as young animals are more likely to disperse in an attempt to find suitable underpopulated areas (e.g. Byrom 2002).
Territoriality
Small carnivorous mustelids typically display intrasexual territoriality (Johnson et al. 2000). Although Powell (1994) suggested that either high or low densities of food can lead to territorial breakdown in mustelids, studies of several exclusively carnivorous mustelid species demonstrate the maintenance of intrasexual territories even in these situations (Balharry 1993; Murphy and Dowding 1994; Herr et al. 2009). Our results clearly demonstrate a lack of either spatial or temporal territoriality between individuals, with both substantial range overlaps and frequent sharing of den sites; especially as these observations are restricted to collared individuals and so represent the minimum of shared occupancy. Field studies in New Zealand (Norbury et al. 1998; Ragg 1998) concur with this conclusion. Home ranges reported here are within the range of values from studies in New Zealand, all of which are based on MCPs or trapping grid captures (summarised in Clapperton and Byrom 2005). However, both the mean and maximum MCP values for female home ranges are lower than those found in previous studies (Clapperton and Byrom 2005). Female home ranges did not shrink to the same extent as those of males, and so they may be closer to their lower limit on Rathlin.
Not only is the lack of territoriality in a carnivorous mustelid unusual, but so is the reduction in range size of both sexes between study years. While such an effect has been demonstrated in the red fox Vulpes vulpes even though ranges then fell below the optimum for resource acquisition (Sargeant 1972), ferret survival known from recapture rates, the observed body condition of animals (T Bodey unpubl. data), and prey consumption (Bodey et al. 2010) does not suggest resources were limiting here. However, even in cases where anthropogenic subsidies allow animals to reduce their home ranges (e.g. small Indian mongoose Herpestes javanicus, Quinn and Whisson 2005), spatial segregation was shown to be maintained. It is likely that the domesticated history of the feral ferret (Blandford 1987) is an important explanatory factor contributing to this increased tolerance of conspecifics and apparent lack of territoriality. This may be particularly relevant in relatively ‘new’ populations such as Rathlin’s (present for approximately 25 years) compared to the more established populations in areas such as New Zealand (>100 years). However, such behaviour reduces the effectiveness of several management options. For example, reducing the number of denning sites, which has been recommended as a non-lethal control option (Frey and Conover 2007), will have a reduced impact in animals that are prepared to share dens. Similarly, ferrets are known to share even small carcasses simultaneously (Ragg et al. 2000, T Bodey pers. obs), and so the availability of anthropogenic dietary subsidies such as edible waste or fallen livestock (Quinn and Whisson 2005; Bino et al. 2010; Bodey et al. 2010) may also be an important consideration as these can sustain higher densities of predators than would otherwise be anticipated.
Implications for predator control programmes
King et al. (2009) recently highlighted a number of the difficulties encountered in eradicating small mustelids. The key problems noted were a lack of accessible traps and latent individual variability, in particular wariness to trapping devices. A time component was not supported in our models, reflecting the high density and ease of access to traps for individuals in our study. Our best supported model did suggest individual variation in response to traps, which may be a result of an increase in young animals, but the behavioural response of the population suggested a general trend towards ‘trap happy’ rather than wary individuals. Although studies with other mustelid species have reported trap-shy behaviour after trapping for radio tracking studies (e.g. Zubergoitia et al. 2006), previous control of the ferret population on Rathlin has been irregular and very limited in extent, and so individuals may not have had an opportunity to learn or become wary (King et al. 2009). Ragg (1997) also found no difference in trappability between adult and juvenile ferrets, suggesting there may not be predictable differences between ‘experienced’ and ‘naïve’ individuals.
The restrictions inevitably imposed on the ferret population by virtue of our island study site are likely to contribute to reductions in home range size as juveniles are unable to disperse as, for example, occurs in mainland populations in New Zealand (Byrom 2002), so these results may be less applicable to a mainland situation. However, self-sustaining feral ferret populations in the UK are all found on relatively small islands such as on the Outer Hebrides and the Isle of Man (Kitchener and Birks 2008), probably because of their inability, again as a result of their domestic past, to successfully compete with wild carnivores (Kitchener and Birks 2008). These islands also frequently support internationally important breeding bird populations (e.g. Mitchell et al. 2004), and similar situations are found on several other European islands where feral ferrets have established such as the Azores, Madeira and Canary Islands (documented in Medina and Martin 2009). Thus our results are likely to have relevance to a number of other island situations in which ferrets are capable of threatening native fauna.
Clearly, where ferrets are introduced to an island, complete eradication is the obvious solution. However, on inhabited islands, this is only feasible if both local and wider-scale public support is obtained, particularly if this will necessitate lifestyle changes perceived as inconvenient, unnecessary or costly to islanders (Oppel et al. in press). In the meantime, it might seem sensible to protect important and/or rare populations through localised control, but control operations will only be effective if reductions in predator density exceed any potential density-dependent response in the population (Barlow et al. 1997). On a small island, it is feasible to examine the impacts of control on the overall population of an introduced vertebrate predator (Russell et al. 2009), a situation we were able to exploit here to examine population responses to the effect of localised culling. However, control over limited areas effectively represents a haphazard harvest of the predator population, and may well be irrelevant to its long-term size, particularly with highly fecund species such as mustelids (McDonald and Harris 2002). It can also, as our study suggests, be counter-productive if it results in an overall increase in predator numbers.
While complete eradication of invasive predators in a mainland context, as opposed to ongoing control, is difficult, advances are being made (e.g. Zabala et al. 2010). Such an approach is to be encouraged, particularly if it can be implemented prior to small scale control that may only serve to ‘educate’ intelligent species (King et al. 2009). Such control is also, in many instances, ultimately a continuous drain on resources, with repeated reinvasion from surrounding unmanaged areas (Parkes and Panetta 2009). In addition, while conditions on a protected area may be known and held constant, this is unlikely to apply to the surrounding environment, and so monitoring is necessary on a wider scale if any meaningful understanding of population-level dynamics, and thus the likely effectiveness of control, is to emerge (Wolff et al. 2002; King et al. 2007).
Consideration of the ecology of the species involved is also essential in order to properly assess levels of threat and determine how management options can most effectively deliver conservation benefits. In territorial species, removal of individuals may lead to increased territory size and decreased predator density (Gese 1998; Baker et al. 2000). Alternatively it may lead to higher rates of dispersal (McDonald and Harris 2002; Woodroffe et al. 2006), and so habitat management may be a more suitable option (Jiménez and Conover 2001). The non-territorial structure of feral ferret populations found both in this study and in New Zealand (Norbury et al. 1998; Ragg 1998) reduces the effectiveness of such management options. The presence of individuals does not preclude the presence of additional animals, and therefore control may seem the best option. But removal can result in enhanced juvenile survival and reinvasion from surrounding areas, leading to the rapid recovery and, particularly within an island context, even increases in the total population. This paradox merits further consideration as it may also act for other flexible species, particularly if defining resources such as shelter or food are not limiting (Frey and Conover 2007).
References
Anonymous (2007) Rathlin seabird colony survey 2007. Allen & Mellon Environmental Ltd, UK
Baker PJ, Funk SM, Harris S, White PCL (2000) Flexible spatial organization of urban foxes, Vulpes vulpes, before and during an outbreak of sarcoptic mange. Anim Behav 59:127–146
Balharry D (1993) Social organization in martens: an inflexible system? In: Dunstone N, Gorman MC (eds) Mammals as predators. Clarendon, Oxford, pp 321–345
Barlow ND, Norbury GL (2001) A simple model for ferret population dynamics and control in semi-arid New Zealand habitats. Wildl Res 28:87–94
Barlow ND, Kean JM, Briggs CJ (1997) Modelling the relative efficacy of culling and sterilisation for controlling populations. Wildl Res 24:129–141
Bertolino S, Genovesi P (2003) Spread and attempted eradication of the grey squirrel (Sciurus carolinensis) in Italy, and consequences for the red squirrel (Sciurus vulgaris) in Eurasia. Conserv 109:351–358
Bino G, Dolev A, Yosha D, Guter A, King R, Saltz D, Kark S (2010) Abrupt spatial and numerical responses of overabundant foxes to a reduction in anthropogenic resources. J Appl Ecol. doi:10.1111/j.1365-2664.2010.01882.x
Blandford PRS (1987) Biology of the polecat Mustela putorius–a literature review. Mammal Rev 17:155–198
Bodey TW, Bearhop S, McDonald RA (2010) The diet of an invasive alien predator the feral ferret Mustela furo: implications for the conservation of ground nesting birds. Eur J Wildl Res. doi:10.1007/s10344-010-0404-y
Bolton M, Tyler G, Smith K, Bamford R (2007) The impact of predator control on lapwing Vanellus vanellus breeding success on wet grassland nature reserves. J Appl Ecol 44:534–544
Bonesi L, Palazon S (2007) The American mink in Europe: status, impacts, and control. Biol Conserv 134:470–483
Bremner A, Park K (2007) Public attitudes to the management of invasive non-native species in Scotland. Biol Conserv 139:306–314
Byrom AE (2002) Dispersal and survival of juvenile feral ferrets Mustela furo in New Zealand. J Appl Ecol 39:67–78
Caley P, McElrea LM, Hone J (2002) Mortality rates of feral ferrets (Mustela furo) in New Zealand. Wildl Res 29:323–328
Chapman DG (1951) Some properties of the hypergeometric distribution with applications to zoological censuses. U Calif Publ Stats 1:131–160
Clapperton BK, Byrom A (2005) Feral ferret. In: King CM (ed) The handbook of New Zealand mammals, 2nd edn. Oxford University Press, Melbourne, pp 294–308
Courchamp F, Langlais M, Sugihara G (2000) Rabbits killing birds: modelling the hyperpredation process. J Anim Ecol 69:154–164
Dowding JE, Murphy EC (2001) The impact of predation by introduced mammals on endemic shorebirds in New Zealand: a conservation perspective. Biol Conserv 99:47–64
Findlay CS, Houlahan J (1997) Anthropogenic correlates of species richness in southeastern Ontario wetlands. Conserv Biol 11:1000–1009
Frey SN, Conover MR (2007) Influence of population reduction on predator home range size and spatial overlap. J Wildl Manag 71:303–309
Gese EM (1998) Response of neighboring coyotes (Canis latrans) to social disruption in an adjacent pack. Can J Zool 76:1960–1963
Greentree C, Saunders G, McLeod L, Hone J (2000) Lamb predation and fox control in south-eastern Australia. J Appl Ecol 37:935–943
Greenwood RJ, Arnold PM, McGuire BG (1990) Protecting duck nests from mammalian predators with fences, traps, and a toxicant. Wildl Soc Bull 18:75–82
Herr J, Schley L, Roper TJ (2009) Socio-spatial organization of urban stone martens. J Zool 277:54–62
Janzen DH (1986) The eternal external threat. In: Soulé ME (ed) Conservation biology: the science of scarcity and diversity. Sinauer Assoc, Sunderland, pp 286–303
Jiménez JE, Conover MR (2001) Ecological approaches to reduce predation on ground-nesting gamebirds and their nests. Wildl Soc Bull 29:62–69
Johnson DDP, Macdonald DW, Dickman AJ (2000) An analysis and review of models of the sociobiology of the Mustelidae. Mammal Rev 30:171–196
Keedwell RJ, Maloney RF, Murray DP (2002) Predator control for protecting kaki (Himantopus novaezelandiae)–lessons from 20 years of management. Biol Conserv 105:369–374
Kenward R (2001) A Manual for Wildlife Radio Tagging. Academic Press, London
King CM, Powell RA (2006) The natural history of weasels and stoats: ecology, behaviour and management, 2nd edn. Oxford University Press, UK
King CM, O’Donnell CFG, Phillipson SM (1994) Monitoring and control of mustelids on conservation lands. Part 2: field and workshop guide. Dept Cons Tech Ser 4:36
King CM, McDonald RM, Martin RD, MacKenzie DI, Tempero GW, Holmes SJ (2007) Continuous monitoring of predator control operations at landscape scale. Ecol Manag Restor 8:133–139
King CM, McDonald RM, Martin RD, Dennis T (2009) Why is the eradication of invasive mustelids so difficult? Biol Conserv 142:806–816
Kinnear JE, Sumner NR, Onus ML (2002) The red fox in Australia—an exotic predator turned biocontrol agent. Biol Conserv 108:335–359
Kitchener AC, Birks JDS (2008) Ferret. In: Harris S, Yalden DW (eds) Mammals of the British isles: handbook, 4th edn. The Mammal Society, Southampton, pp 485–487
Lahti DC (2001) The “edge effect on nest predation” hypothesis after twenty years. Biol Conserv 99:365–374
Lockwood JL, Hoopes MF, Marchetti MP (2007) Invasion Ecology. Blackwell, UK
Long JL (2003) Introduced mammals of the world. CABI Publishing, Wallingford
Macdonald DW, Harrington LA (2003) The American mink: the triumph and tragedy of adaptation out of context. N Z J Zool 30:421–441
McDonald RA, Harris S (2002) Population biology of stoats Mustela erminea and weasels Mustela nivalis on game estates in Great Britain. J Appl Ecol 39:793–805
McDonald RA, Vaughan N (1999) An efficient way to prepare mammalian skulls and bones. Mammal Rev 29:265–266
McDonald RA, Delahay RJ, Carter SP, Smith GC, Cheeseman CL (2008) Perturbing implications of wildlife ecology for disease control. Trend Ecol Evol 23:53–56
McKinney ML, Lockwood JL (1999) Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trend Ecol Evol 14:450–453
Medina FM, Martin A (2009) A new invasive species in the Canary Islands: a naturalized population of ferrets Mustela furo in La Palma Biosphere Reserve. Oryx 44:41–44
Minta SC (1993) Sexual differences in spatiotemporal interaction among badgers. Oecologia 96:402–409
Mitchell PI, Newton SF, Ratcliffe N, Dunn TE (2004) Seabird populations of Britain and Ireland: results of the ‘Seabird 2000’ census 1999–2002. Poyser, London
Morrison SA, Macdonald N, Walker K, Lozier L, Shaw MR (2007) Facing the dilemma at eradication’s end: uncertainty of absence and the Lazarus effect. Front Ecol Environ 5:271–276
Murphy EC, Dowding JE (1994) Range and diet of stoats (Mustela erminea) in a New Zealand beech forest. N Z J Ecol 18:11–18
Norbury GL, Norbury DC, Heyward RP (1998) Space use and denning behaviour of wild ferrets (Mustela furo) and cats (Felis catus). N Z J Ecol 22:149–159
Oksanen L (2001) Logic of experiments in ecology: is pseudoreplication a pseudoissue? Oikos 94:27–38
Oppel S, Beaven BM, Bolton M, Vickery J, Bodey TW (in press) Eradication of invasive mammals on islands inhabited by humans and domestic animals. Conserv Biol. doi: 10.1111/j.1523-1739.2010.01601.x
Otis DL, Burnham KP, White GC, Anderson DR (1978) Statistical inference from capture data on closed animal populations. Wildl Monogr 62
Parkes J, Panetta D (2009) Eradication of invasive species: progress and emerging issues in the 21st century. In: Clout MN, Williams PA (eds) Invasive species management. A handbook of principles and techniques. Oxford University Press, Oxford, pp 47–60
Powell RA (1994) Structure and spacing of Martes populations. In: Buskirk SW, Harested AS, Raphael MG, Powell RA (eds) Martens, sables, and fishers, biology and conservation. Cornell University Press, Ithaca, pp 101–121
Quinn JH, Whisson DA (2005) The effects of anthropogenic food on the spatial behaviour of small Indian mongooses (Herpestes javanicus) in a subtropical rainforest. J Zool 267:339–350
Ragg JR (1997) Tuberculosis (Mcyobacterium bovis) epidemiology and the ecology of ferrets (Mustela furo) on New Zealand farmland. PhD thesis, University of Otago, Dunedin
Ragg JR (1998) The denning behaviour of feral ferrets (Mustela furo) in a pastoral habitat, South Island, New Zealand. J Zool 246:471–477
Ragg JR, Mackintosh CG, Moller H (2000) The scavenging behaviour of ferrets (Mustela furo), feral cats (Felis domesticus), possums (Trichosurus vulpecula), hedgehogs (Erinaceus europaeus) and harrier hawks (Circus approximans) on pastoral farmland in New Zealand: Implications for bovine tuberculosis transmission. N Z Vet J 48:166–175
Rooney SM, Wolfe A, Hayden TJ (1998) Autocorrelated data in telemetry studies: time to independence and the problem of behavioural effects. Mammal Rev 28:89–98
Russell JC, Abdelkrim J, Fewster RM (2009) Early colonisation population structure of a Norway rat island invasion. Biol Invasions 11:1557–1567
Sargeant AB (1972) Red fox spatial characteristics in relation to waterfowl predation. J Wildl Manag 36:225–236
Seber GAF (1973) The estimation of animal abundance and related parameters. Griffin, London
Sinclair ARE (1996) Mammal populations: fluctuation, regulation, life history theory and their implications for conservation. In: Floyd RB, Sheppard AW, De Barro PJ (eds) Frontiers of Population Ecology. CSIRO, Melbourne, pp 127–154
Sinclair ARE, Pech RP, Dickman CR, Hik D, Mahon P, Newsome AE (1998) Predicting effects of predation on conservation of endangered prey. Conserv Biol 12:564–575
Smith RK, Pullin AS, Stewart GB, Sutherland WJ (2010) Effectiveness of predator removal for enhancing bird populations. Conserv Biol 24:820–829
Sutherland WJ, Pullin AS, Dolman PM, Knight TM (2004) The need for evidence-based conservation. Trend Ecol Evol 19:305–308
Towns DR, Broome KG (2003) From small Maria to massive Campbell: forty years of rat eradications from New Zealand islands. N Z J Zool 30:377–398
Veitch CR, Clout MN (2002) Turning the tide: the eradication of invasive species In: proceedings of the international conference on eradication of island invasives. IUCN, Gland, Switzerland and Cambridge, UK
Vitousek PM, Mooney HA, Lubchenco J, Melillo JM (1997) Human domination of Earth’s ecosystems. Science 277:494–499
White GC, Burnham KP (1999) Program MARK: survival estimation from populations of marked animals. Bird Study 46(S):120–138
Wolff A, Dieuleveut T, Martin JL, Bretagnolle V (2002) Landscape context and little bustard abundance in a fragmented steppe: implications for reserve management in mosaic landscapes. Biol Conserv 107:211–220
Woodroffe R, Donnelly CA, Cox DR, Bourne FJ, Cheeseman CL, Delahay RJ, Gettinby G, McInerney JP, Morrison WI (2006) Effects of culling on badger Meles meles spatial organization: implications for the control of bovine tuberculosis. J Appl Ecol 43:1–10
Zabala J, Zuberogoitia I, González-Oreja JA (2010) Estimating costs and outcomes of invasive American mink (Neovison vison) management in continental areas: a framework for evidence based control and eradication. Biol Invasions. doi:10.1007/s10530-010-9690-2
Zubergoitia I, Zabala J, Martínez JA (2006) Evaluation of sign surveys and trappability of American mink: management consequences. Folia Zool 55:257–263
Acknowledgments
TWB was supported by a PhD studentship through the Quercus partnership between Northern Ireland Environment Agency & Queen’s University Belfast. We thank the residents and landowners of Rathlin for allowing access; Liam McFaul, Rosalind Kennerley, Jonathan Reeves, Oisin Sweeney, Rory Finlay, Marc Ruddock and David Tosh for assistance with fieldwork; and three anonymous reviewers for improving the manuscript. All fieldwork on animals was approved by the Queen’s University, Belfast ethics committee, and scientific procedures on animals were carried out under Home Office licence where appropriate.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Bodey, T.W., Bearhop, S. & McDonald, R.A. Localised control of an introduced predator: creating problems for the future?. Biol Invasions 13, 2817–2828 (2011). https://doi.org/10.1007/s10530-011-9965-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-011-9965-2