
Abstract
Thirty-nine strains representing 12 diverse phylogenetic clades of Aureobasidium pullulans were surveyed for lipase production using a quantitative assay. Strains in clades 4 and 10 produced 0.2–0.3 U lipase/ml, while color variant strain NRRL Y-2311-1 in clade 8 produced 0.54 U lipase/ml. Strains in clade 9, which exhibit a dark olivaceous pigment, produced the highest levels of lipase, with strain NRRL 62034 yielding 0.57 U lipase/ml. By comparison, Candida cylindracea strain NRRL Y-17506 produced 0.05 U lipase/ml under identical conditions. A. pullulans strain NRRL 62034 reached maximal lipase levels in 5 days on lipase induction medium, while A. pullulans strain NRRL Y-2311-1 and strains in clades 4 and 10 were highest after 6 days. A. pullulans strain NRRL Y-2311-1 and strains in clade 9 produced two extracellular proteins in common, at >50 and <37 kDa.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Aureobasidium pullulans is a polymorphic fungus, considered to be a filamentous ascomycete in class Dothideomycetes, subclass Dothideomycetidae (Hibbett et al. 2007; Schoch et al. 2006). It is well known as the source of the commercial polysaccharide, pullulan (Leathers 2002; Singh et al. 2008). Strains of A. pullulans also produce numerous degradative enzymes, including fructofuranosidase, glucoamylase, laccase, and xylanase (Leathers 1989; Deshpande et al. 1992; Rich et al. 2013). We recently developed a multilocus molecular phylogeny of A. pullulans based on sequences of five genetic loci (Manitchotpisit et al. 2009). Interestingly, certain phylogenetically defined clades produced high levels of specific valuable bioproducts, including pullulan, xylanase, poly(β-l-malic acid) (PMA), laccase, and liamocins (heavy oil) (Manitchotpisit et al. 2009, 2011, 2012; Rich et al. 2013). However, relatively little information has been available concerning lipase production by A. pullulans.
Microbial lipases have numerous biotechnological applications in the detergent, food, and pharmaceutical industries (Hasan et al. 2006; Salihu and Alam 2012; Singh and Mukhopadhyay 2012). Lipases also can be used in the production of biodiesel fuels (Tan et al. 2010; Ghaly et al. 2010). Numerous bacteria and fungi produce lipases, including species of the yeastlike fungus Candida (Sharma et al. 2011; Patil et al. 2011). Federici (1982) reported that all of 198 strains of A. pullulans produced lipase activity in semi-quantitative plate assays. Buzzini and Martini (2002) found that 20 of 46 tropical isolates from the Brazilian rain forest produced lipase activity in a qualitative plate assay. Similarly, Kudanga et al. (2007) reported that 20 of 42 tropical isolates from Zimbabwe produced lipase. Wang et al. (2007) isolated a lipase-producing marine fungus which they described as A. pullulans, and only lipase from this single strain has been studied in greater detail (Liu et al. 2008a, b). In this study we survey 39 strains representing 12 diverse phylogenetic clades of A. pullulans for lipase production using quantitative assays and identify specific clades that produce lipase.
Materials and methods
Organisms and growth conditions
Strains used in this study were obtained from the ARS Culture Collection, Peoria, IL (Table 1). Strains were maintained at 28 °C on potato/dextrose/agar (PDA). Single colonies from 24 h plates were used to inoculate liquid preinocula, which were grown overnight in 10 ml yeast peptone/dextrose broth (YPD) in 50 ml flasks at 28 °C, 200 rpm. Preinocula were used to inoculate triplicate 250 ml flasks to a final OD600 of 0.04 in 50 ml lipase induction medium (Liu et al. 2008a) containing 0.4 % (w/v) glucose, 0.6 % (w/v) (NH4)2SO4, 0.1 % (w/v) K2HPO4, and 0.05 % (w/v) MgSO4·7H2O, pH 7.0. After 6 h at 25 °C, 130 rpm, 3 % (v/v) sterile olive oil was added and cultures were further incubated for up to 6 days (Liu et al. 2008a).
Lipase activity assay
One ml samples of lipase induction cultures were centrifuged for 20 min at ~16000×g and supernatants were assayed for lipase activity by measurement of p-nitrophenol released from p-nitrophenyl laurate. Substrate emulsions were prepared as described by Liu et al. (2008a). Six microliters of sample were added to 194 μl of freshly prepared substrate emulsion and incubated in a Molecular Devices SpectraMax M5 plate reader at 35 °C for 25 min. Product was detected at 410 nm. Lipase from porcine pancreas served as a standard and positive control. Boiled samples served as negative controls. Enzyme activity was expressed in units/ml (1 U = 1 μmol product formed/min) and as specific activity (U/mg protein). Standard errors are reported. Protein concentrations were determined using the Bio-Rad protein assay, based on the Bradford dye-binding method with bovine serum albumin as standard.
Polyacrylamide gel electrophoresis
Samples were concentrated 50-fold by ultrafiltration (Nanosep 3 K Omega low protein binding spin cells) and denatured for 2 min at 95 °C in 2 × SDS sample buffer (4.0 % (w/v) SDS, 20 % (v/v) glycerol, 0.005 % (w/v) bromophenol blue, 0.126 M Tris–HCl pH 6.8, and 5.0 % (v/v) β-mercaptoethanol). Samples and Bio-Rad Precision Plus Unstained Protein Standards (Bio-Rad) were applied to an SDS-PAGE gel (5 % v/v stacking, 10 % v/v resolving). After electrophoresis at 100 V for ~1 h, the gel was stained with SYPRO Ruby protein gel stain (Invitrogen, Grand Island, NY).
Results and discussion
Production of lipase by diverse phylogenetic clades of A. pullulans
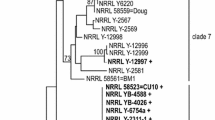
Based on a multilocus molecular phylogeny of A. pullulans (Manitchotpisit et al. 2009), we surveyed 39 strains representing 12 diverse phylogenetic clades for production of lipase (Table 1). Seven reference strains previously described as lipase producers also were tested as controls. A. pullulans strains in phylogenetic clade 7 produced no detectable lipase (<0.01 U/ml) after 6 days of induction under conditions tested (Table 1). Several clades (1, 2, 3, 5, 6, 11, and 13) produced lipase at levels generally less than 0.1 U lipase/ml. Some of these clades produce other valuable bioproducts. For example, strains in clade 1 produce high levels of pullulan without contaminating pigment (Manitchotpisit et al. 2009). Strains in clade 5 produce relatively high levels of laccase (Rich et al. 2013), while strains in clade 13 produce high levels of PMA (Manitchotpisit et al. 2012). Strains in clades 4 and 10 produced 0.2–0.3 U lipase/ml. A. pullulans strain NRRL Y-2311-1 in clade 8 produced 0.54 U lipase/ml. Clade 8 includes the so-called “color variant” strains that produce brilliant pigments of red, yellow, and orange (Wickerham and Kurtzman 1975). These strains produce high levels of xylanase (Leathers 1986; Manitchotpisit et al. 2009). Strains in clade 9 consistently produced the highest levels of lipase among those tested (Table 1). In particular A. pullulans strain NRRL 62034 produced 0.57 U lipase/ml under these conditions. By comparison, the best of seven reference strains, Candida cylindracea strain NRRL Y-17506, produced 0.05 U lipase/ml (Table 1). Under optimized conditions, strain NRRL Y-17506 has been reported to produce 20 U lipase/ml (Salihu et al. 2011). Strain HN2.3 of a marine fungus described as A. pullulans, of unknown phylogenetic affiliation, reportedly produced 8.02 U lipase/ml (Liu et al. 2008a). Lipase activities from other fungi vary considerably, but are often in this range (Singh and Mukhopadhyay 2012). A. pullulans phylogenetic clade 9 was not previously recognized for production of valuable bioproducts. However, this clade is distinguished by production of dark olivaceous pigment (Manitchotpisit et al. 2009).
We have observed a general relationship in A. pullulans between the capacity to produce pigment and the production of certain hydrolytic enzymes. Color variant strains in clade 8 are high-level producers of xylanase, while strains in clade 5, which exhibit a dark vinaceous pigment, produce relatively high levels of laccase. In this study, clade 9, strains of which exhibit a dark olivaceous pigment, produced the highest levels of lipase. On the contrary, clades that produce high levels of pullulan, PMA, and liamocins are not associated with pigment production (Manitchotpisit et al. 2009, 2011, 2012). The reason for the association of enzyme production with pigment is unclear. Most of the genetic diversity of A. pullulans occurs in tropical environments, and in fact most isolates from temperate climates have thus far been found in clade 13. Although pigment production might be assumed to be protective against environmental exposure to sunlight, strains from diverse phylogenetic clades have been isolated side-by-side from similar collection sites (Manitchotpisit et al. 2009). In the case of laccase production, the enzyme may be directly involved in pigment biosynthesis (Rich et al. 2013).
Extracellular proteins from lipase-producing strains of A. pullulans
Total extracellular proteins from strains representing diverse phylogenetic clades of A. pullulans were compared by SDS-PAGE (Fig. 1). Strains from clade 9 showed two extracellular proteins in common, at >50 and <37 kDa (Fig. 1). Interestingly, clade 8 strain NRRL Y-2311-1, which made similar levels of lipase, produced similar protein species (Fig. 1). Strains in clades 1, 2, 3, 4, 11, and 13 also produced one or more of these species, as did Penicillium citrinum reference strain NRRL 3754. Fungal lipases vary considerably in molecular weight but have often been reported in this range (Sharma et al. 2011; Patil et al. 2011). Lipase isoforms of approximately 60 kDa have been purified from C. cylindracea strain NRRL Y-17506 (Rua et al. 1993; Benjamin and Pandey 2001). Similarly, a lipase purified from A. pullulans strain HN2.3 was estimated to be 63.5 kDa (Liu et al. 2008a). Strains in clades 5, 6, and 10 produced diffuse, high molecular weight protein species, which may be glycosylated forms (Fig. 1).

SDS-PAGE of extracellular proteins produced by Aureobasidium pullulans strains from diverse phylogenetic clades and reference strains after 6 days on lipase induction medium. Lane M Bio-Rad Precision Plus protein standards; lane 1 A. pullulans strain NRRL 58555 (clade 1); lane 2 A. pullulans strain NRRL 62032 (clade 1 or 2); lane 3 A. pullulans strain NRRL 58522 (clade 2); lane 4 A. pullulans strain NRRL 62043 (clade 3); lane 5 A. pullulans strain NRRL 58534 (clade 4); lane 6 A. pullulans strain NRRL 58519 (clade 5); lane 7 A. pullulans strain NRRL 58546 (clade 6); lane 8 A. pullulans strain NRRL Y-2311-1 (clade 8); lane 9 A. pullulans strain NRRL 62034 (clade 9); lane 10 A. pullulans strain NRRL 62039 (clade 9); lane 11 A. pullulans strain NRRL 62040 (clade 9); lane 12 A. pullulans strain NRRL Y-12974 (clade 10); lane 13 A. pullulans strain NRRL 62031 (clade 11); lane 14 A. pullulans strain NRRL 58012 (clade 13); lane 15 Penicillium citrinum strain NRRL 3754 (reference strain); lane 16 Candida cylindracea strain NRRL Y-17506 (reference strain)
Time course of lipase production by A. pullulans strains
Aureobasidium pullulans strains representing the highest lipase-producing clades (4, 8, 9, and 10) were cultured for 6 days in lipase induction medium. Clade 9 strain NRRL 62034 reached maximal lipase production levels (~0.6 U lipase/ml) within 5 days, while clade 8 strain NRRL Y-2311-1 required an additional day to reach equivalent levels (Fig. 2). Strains in clades 4 and 10 produced about 0.2 U lipase/ml in 6 days. By comparison C. cylindracea strain NRRL Y-17506 produced a maximum of about 0.1 U lipase/ml in 5 days. Lipase from C. cylindracea strain NRRL Y-17506 is widely used in research due to its high activities in hydrolysis and synthesis (Rua et al. 1993), and the enzyme is commercially produced (Singh and Mukhopadhyay 2012).

Time course of lipase production by Aureobasidium pullulans strains and Candida cylindracea strain NRRL Y-17506 grown for 6 days on lipase induction medium
Conclusion
we report here for the first time lipase production by diverse phylogenetic clades of A. pullulans. Strains in phylogenetic clade 9 are most promising for lipase production, the best of which is strain NRRL 62034. Strains in this clade are distinguished by production of dark olivaceous pigment. This may provide a simple method to rapidly screen new isolates for lipase production. Under identical conditions, strain NRRL 62034 produced approx. six times as much lipase as commercial strain C. cylindracea NRRL Y-17506.
References
Benjamin S, Pandey A (2001) Isolation and characterization of three distinct forms of lipases from Candida rugosa produced in solid state fermentation. Braz Arch Biol Technol 44:213–221
Buzzini P, Martini A (2002) Extracellular enzymatic activity profiles in yeast and yeast-like strains isolated from tropical environments. J Appl Microbiol 93:1020–1025
Deshpande MS, Lynch JM, Rale VB (1992) Aureobasidium pullulans in applied microbiology: a status report. Enzyme Microbial Technol 14:514–527
Federici F (1982) Extracellular enzymatic activities in Aureobasidium pullulans. Mycologia 74:738–743
Ghaly AE, Dave D, Brooks MS, Budge S (2010) Production of biodiesel by enzymatic transesterification: review. Amer J Biochem Biotechnol 6:54–76
Hasan F, Shah AA, Hameed A (2006) Industrial applications of microbial lipases. Enzyme Microbial Technol 39:235–251
Hibbett DS, Binder M, Bischoff JF et al (2007) A higher-level phylogenetic classification of the fungi. Mycol Res 111:509–554
Hou CT, Johnston TM (1992) Screening of lipase activities with cultures from the agricultural research service culture collection. J Amer Oil Chem Soc 69:1088–1097
Kudanga T, Mwenje E, Mandivenga F, Read JS (2007) Esterases and putative lipases from tropical isolates of Aureobasidium pullulans. J Basic Microbiol 47:138–147
Leathers TD (1986) Color variants of Aureobasidium pullulans overproduce xylanase with extremely high specific activity. Appl Environ Microbiol 52:1026–1030
Leathers TD (1989) Purification and properties of xylanase from Aureobasidium. J Ind Microbiol 4:341–348
Leathers TD (2002) Pullulan. In: Vandamme EJ, De Baets S, Steinbüchel A (eds) Biopolymers, vol 6., Polysaccharides II: polysaccharides from eukaryotesWiley, Weinheim, pp 1–35
Liu Z, Chi Z, Wang L, Li J (2008a) Production, purification and characterization of an extracellular lipase from Aureobasidium pullulans HN2.3 with potential application for the hydrolysis of edible oils. Biochem Eng J 40:445–451
Liu Z, Li X, Chi Z, Wang L, Li J, Wang X (2008b) Cloning, characterization and expression of the extracellular lipase gene from Aureobasidium pullulans HN2-3 isolated from sea saltern. Antonie Van Leeuwenhoek 94:245–255
Manitchotpisit P, Leathers TD, Peterson SW, Kurtzman CP, Li X-L, Eveleigh DE, Lotrakul P, Prasongsuk S, Dunlap CA, Vermillion KE, Punnapayak H (2009) Multilocus phylogenetic analyses, pullulan production and xylanase activity of tropical isolates of Aureobasidium pullulans. Mycol Res 113:1107–1120
Manitchotpisit P, Price NPJ, Leathers TD, Punnapayak H (2011) Heavy oils produced by Aureobasidium pullulans. Biotechnol Lett 33:1151–1157
Manitchotpisit P, Skory CD, Peterson SW, Price NPJ, Vermillion KE, Leathers TD (2012) Poly(β-l-malic acid) production by diverse phylogenetic clades of Aureobasidium pullulans. J Ind Microbiol Biotechnol 39:125–132
Patil KJ, Chopda MZ, Mahajan RT (2011) Lipase biodiversity. Ind J Sci Technol 4:971–982
Rich JO, Leathers TD, Anderson AM, Bischoff KM, Manitchotpisit P (2013) Laccases from Aureobasidium pullulans. Enz Microb Technol 53:33–37. doi:10.1016/j.enzmictec.2013.03.015
Rua ML, Diaz-Maurino T, Fernandez VM, Otero C, Ballesteros A (1993) Purification and characterization of two distinct lipases from Candida cylindracea. Biochim Biophys Acta 1156:181–189
Salihu A, Alam MZ (2012) Production and applications of microbial lipases: a review. Sci Res Essays 7:2667–2677
Salihu A, Alam MZ, AbdulKarim MI, Salleh HM (2011) Optimization of lipase production by Candida cylindracea in palm oil mill effluent based medium using statistical experimental design. J Mol Catal B 69:66–73
Schoch CL, Shoemaker RA, Seifert KA, Hambleton S, Spatafora JW, Crous PW (2006) A multigene phylogeny of the Dothideomycetes using four nuclear loci. Mycologia 98:1042–1053
Sharma D, Sharma B, Shukla AK (2011) Biotechnological approach of microbial lipase: a review. Biotechnol 10:23–40
Singh AK, Mukhopadhyay M (2012) Overview of fungal lipase: a review. Appl Biochem Biotechnol 166:486–520
Singh RS, Saini GK, Kennedy JF (2008) Pullulan: microbial sources, production and applications. Carbohydr Polym 73:515–531
Tan T, Lu J, Nie K, Deng L, Wang F (2010) Biodiesel production with immobilized lipase: a review. Biotechnol Adv 28:628–634
Wang L, Chi Z, Wang X, Liu Z, Li J (2007) Diversity of lipase-producing yeasts from marine environments and oil hydrolysis by their crude enzymes. Ann Microbiol 57:495–501
Wickerham LJ, Kurtzman CP (1975) Synergistic color variants of Aureobasidium pullulans. Mycologia 67:342–361
Zhang A, Gao R, Diao N, Xie G, Gao G, Cao S (2009) Cloning, expression and characterization of an organic solvent tolerant lipase from Pseudomonas fluorescens JCM5963. J Mol Catal B 56:78–84
Author information
Authors and Affiliations
Corresponding author
Additional information
Mention of any trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the US Department of Agriculture. USDA is an equal opportunity provider and employer.
Rights and permissions
About this article
Cite this article
Leathers, T.D., Rich, J.O., Anderson, A.M. et al. Lipase production by diverse phylogenetic clades of Aureobasidium pullulans . Biotechnol Lett 35, 1701–1706 (2013). https://doi.org/10.1007/s10529-013-1268-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10529-013-1268-5