
Abstract
Chronic lymphocytic leukemia (CLL) is a clonal proliferative disease of mature B lymphocytes. To further improve the prognosis of patients, it is necessary to further elucidate the pathogenesis of CLL and find more effective therapeutic targets. Nuclear Factor of Activated T cells 5 (NFAT5) is the major activated transcription factor (TF) upon osmotic pressure increase in mammalian cells, and it also regulates many target genes to affect various cellular functions. The effects of NFAT5 on tumor growth and metastasis have also been widely revealed. However, the effects of NFAT5 on the progression of CLL are still unclear. In this study, we found abnormally high expression of NFAT5 in human CLL patients. Additionally, NFAT5 depletion suppressed proliferation and stimulated apoptosis of CLL cells. Our data further confirmed NFAT5 regulated AQP5 expression and the phosphorylation of p38 MAPK. We also found that AQP5 overexpression reversed the inhibitory effect of NFAT5 depletion on cell proliferation in CLL cells. Furthermore, we revealed STUB1 directly bound to NFAT5 and promoted its degradation. Taken together, our results indicate the involvement of NFAT5 in CLL progression and suggest that NFAT5 could serve as a promising therapeutic target for CLL treatment.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Chronic lymphocytic leukemia (CLL) is a clonal proliferative disease of mature B lymphocytes with slow clinical progression characterized by lymphocytic accumulation in peripheral blood, bone marrow, spleen, and lymph nodes (Xu et al. 2020b). This disease is one of the most common malignancies of the blood system in European and American countries (Koch et al. 2020). Early CLL often lacks significant symptoms, and there is a lack of effective treatments in the progressive stage (Sedlarikova et al. 2020). In recent years, a series of positive progress has been made in targeted therapy for CLL (Barthel et al. 2020). To further improve the prognosis of patients, it is necessary to further elucidate the pathogenesis of CLL and find more effective therapeutic targets.
Nuclear Factor of Activated T cells 5 (NFAT5) is the major activated transcription factor (TF) upon osmotic pressure increase in mammalian cells. NFAT5 protects cells from hypertonic stimulation by increasing transcription of genes (Kumar et al. 2020). In addition, as a pluripotent transcription factor, NFAT5 regulates many target genes and thus affects multiple cellular functions (Fernandez-Castillo et al. 2015; Halterman et al. 2012; Ito et al. 2007). A previous study showed that in skin macrophages, activation of NFAT5 exerted regulatory effects on vascular endothelial growth factor C (VEGF-C), maintaining normal cell function and blood pressure stability through the NFAT5-VEGF-C axis (Amara et al. 2016). In addition, NFAT5 coordinates HIF-1α and therefore regulates NKCC1 expression in hippocampal neurons after hypoxia–ischemia (Villanueva et al. 2012).
The effects of NFAT5 on tumor growth and metastasis have also been widely revealed (Kim et al. 2018; Meng et al. 2019; Qin et al. 2017). The expression of NFAT5 in lung adenocarcinoma (LUAD) cells was increased, and knockdown of NFAT5 inhibited the proliferation and migration of LUAD cells (Meng et al. 2019). In addition, the expression of NFAT5 in renal cancer cell line CaKi-1 was significantly higher than that in normal cells, and NFAT5 knockdown resulted in the decrease of S100A4 expression, which inhibited cancer cell proliferation and migration (Kuper et al. 2014). NFAT5 may also play an important role in the pathogenesis of inflammatory breast cancer (Remo et al. 2015).
It is worth noting that NFAT5 has a significant effect on lymphocyte survival (Trama et al. 2000). NFAT5 could regulate the expression of AQP5, while AQP5 could phosphorylate and activate p38 MAPK pathway (Lee et al. 2011). However, the effects of NFAT5 and NFAT5/AQP5 axis on the progression of CLL are still unclear. In this study, we found that the expression of NFAT5 in CLL cells was up-regulated, and further revealed the mechanisms of NFAT5 on the pathogenesis of CLL.
Material and Methods
CLL Samples
Blood samples from patients (40 samples) and healthy donors (20 samples) were collected after receiving informed consent from Affiliated Hospital of Guilin Medical University. B cells from CLL patients were obtained using RosetteSep™ B cell isolation cocktail (Stem Cell Technologies, Vancouver, Canada) per the manufacturer’s protocol (Chae et al. 2008).
All procedures performed in studies involving human participants were in accordance with the standards upheld by the Ethics Committee of Affiliated Hospital of Guilin Medical University and with those of the 1964 Helsinki Declaration and its later amendments for ethical research involving human subjects.
Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
Total RNA was isolated using TRIzol reagent (Invitrogen, USA). RNA was reverse transcribed into cDNA using reverse transcriptase (Thermo Fisher, USA). SYBR Green Supermix (Bio-Rad, USA) was ultilized for qRT-PCR analysis on the Bio-Rad CFX96 system (Bio-Rad, USA). Primers used in the qRT-PCR were listed as follows. NFAT5: forward: 5ʹ-CTCCTCAGATCCAGTTGGTTCA-3′, reverse: 5′-GCTGCATGTCTGGTTGGTTTAT-3′; GAPDH: forward: 5ʹ-CTCTGCTCCTCCTGTTCGAC-3′, reverse: 5ʹ-ACCAAATCCGTTGACTCCGA-3′.
Cell Lines and Transfection
MEC-1 cell lines purchased from the American Type Culture Collection (ATCC, Manassas, VA) and B cells isolated from CLL patients were cultured with RPMI-1640 supplied with 20% fetal bovine serum and penicillin/streptomycin. For NFAT5 knockdown, NFAT5 shRNA or control shRNA, pcDNA3.1-AQP5, and pcDNA3.1-STUB1 (Origene, Rockville, MD, USA) and empty vector were transfected into cells using Lipofectamine 2000 (Thermo Fisher Scientific), respectively. NFAT5, AQP5, and STUB1 expression was measured 48 h after transfection.
Immunoblot
The total proteins were collected with RIPA lysis buffer and separated by SDS-PAGE. After transferring proteins onto PVDF membranes, membranes were blocked and subsequently incubated using the specific antibodies against NFAT5 (1:500 dilution, sc-398171, Santa Cruz Biotechnology, Dallas, TX, USA), β-actin (1:5000 dilution, ab8226, Abcam, Cambridge, MA, USA), BCL-2 (1:500 dilution, ab182858, Abcam), cleaved caspase-3 (1:1000 dilution, ab32042, Abcam), p38 (1:1000 dilution, ab170099, Abcam) and p-p38 (1:500 dilution, ab178867, Abcam). Then the membranes were subjected to HRP-conjugated secondary antibodies for 1 h. Signals were visualized by an ECL kit.
Flow Cytometry Analysis
For flow cytometry detection, cultured cells were collected by trypsinization and washed with PBS. Then cells underwent annexin V/propidium iodide (PI) staining (Dojindo, Kumamoto, Japan) following the guidelines of the manufacture.
BrdU Incorporation Assays
Cells were fixed in 4% paraformaldehyde and permeabilized in PBST. 10 µM BrdU labeling solution was added to cells for 1 h at 37 °C in a CO2 incubator. Then the BrdU labeling solution was removed from the cells and the cells were washed in PBS for three more times.
Statistics
The correlations between clinical features of patients and NFAT5 expression was analyzed through χ2 analysis. Student’s t test was used for statistical analysis. p < 0.05 was considered as significantly different. * p < 0.05, **p < 0.01.
Results
NFAT5 Expression Was Upregulated in CLL Patients
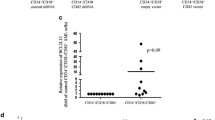
To measure the expression level of NFAT5 in CLL patients, we collected 40 CLL blood samples, 20 healthy donor samples and B cells from CLL patients. Through qRT-PCR and Immunoblot assays, we found the increased levels of NFAT5 in CLL patient compared with healthy donors (Fig. 1a–c). According to the relative expression level of NFAT5, patients were divided into NFAT5 high group and NFAT5 low group based on the median NFAT5 level. As shown in Table 1, high expression of NFAT5 in CLL was associated with β2-MG (p = 0.025), Binent stage (p = 0.015) and Rai risk stage (p = 0.024). There was no association between the expression level of NFAT5 and age (p = 0.736), high leukocytosis (p = 0.288), IgVH (p = 0.110), ZAP-70 (p = 0.490), CD38 (p = 0.091) or FISH cytogenetics (p = 0.525).

NFAT5 was upregulated in CLL patients. a–c Quantitative PCR (a) and Immunoblot assay (b, c) were performed to measure mRNA and protein levels of NFAT5 in CLL patients and healthy donors. Results are presented as mean ± SD, ***p < 0.001
NFAT5 Knockdown Inhibited the Proliferation of MEC-1 and B Cells from CLL Patients
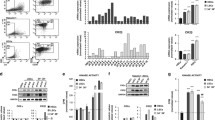
Since we found that NFAT5 level was increased in CLL patients and correlated with risk stage of CLL, we next aim to explore the potential role of NFAT5 in CLL. We depleted NFAT5 by transfection of NFAT5 shRNA. qRT-PCR and Immunoblot assays were conducted to evaluate the knockdown efficiency. NFAT5 was reduced in NFAT5 shRNA transfected cells as expected (Fig. 2a, b). Subsequently, BrdU assay was used to determine the effect of NFAT5 silencing on the proliferation of MEC-1 and B cells from CLL patients (Fig. 2c). The proliferation rates of MEC-1 and B cell from CLL patients in the NFAT5 shRNA group was significantly lower compared with those in the control shRNA groups (Fig. 2c). Taken together, NFAT5 knockdown inhibited the proliferation of MEC-1 cells and B cells from CLL patients.

NFAT5 knockdown inhibited the proliferation of MEC-1 and B cell from CLL patients (CLL-B). a Quantitative PCR assays were performed to measure the mRNA levels of NFAT5 in MEC-1 and B cell from CLL patients upon transfection of indicated shRNA. b Immunoblot assay was conducted to measure the expression of NFAT5 in MEC-1 and CLL-B cells upon transfection of indicated shRNA. c BrdU incorporation assay was performed to evaluate the proliferation ability of control or NFAT5-depleted MEC-1 or CLL-B cells. Results are presented as mean ± SD, **p < 0.01
NFAT5 Gene Silencing Promoted Apoptosis in MEC-1 Cells and B Cells of CLL Patients
NFAT5’s function in apoptosis was evaluated by flow cytometry assays. We noticed NFAT5 knockdown increased apoptosis both in MEC-1 cells and B cells of CLL patients (Fig. 3a). Furthermore, apoptosis-related protein levels were monitored as well. As shown in Fig. 3b, BCL-2 was reduced, and cleaved caspase-3 was increased in NFAT5-depleted MEC-1 cells and B cells of CLL patients. These results indicated that NFAT5 gene silencing increased apoptosis in MEC-1 cells and B cells of CLL patients.

NFAT5 gene silencing facilitated apoptosis in MEC-1 cells and B cells of CLL patients. a Flow cytometry assay was performed to evaluate apoptosis of NFAT5 in MEC-1 and CLL-B cells upon indicated transfection. b Immunoblot was performed to measure the protein levels of BCL-2 and cleaved caspase-3 in MEC-1 and CLL-B cells upon the indicated transfection. Results are presented as mean ± SD, **p < 0.01
NFAT5 Regulated AQP5 and p38 MAPK Activation
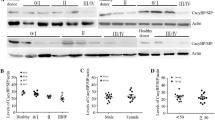
AQP5 was reported to be overexpressed in CML cells and play a role in promoting cell proliferation and inhibiting apoptosis (Chae et al. 2008). We found decreased AQP5 expression in MEC-1 and CLL-B cells following NFAT5 shRNA transfection (Fig. 4a). We also assessed the impact of NFAT5 knockdown on AQP5 level. As determined by Immunoblot, NFAT5 depletion resulted in decreased AQP5 level. NFAT5 depletion also reduced phosphorylated p38 protein level without affecting total p38 protein level (Fig. 4b). These results indicated that NFAT5 regulated AQP5 level and promoted p38 activation simultaneously.

NFAT5 regulated AQP5 and p38 MAPK activation. a The mRNA level of AQP5 in control or NFAT5-depleted MEC-1 cells and CLL-B cells was measured by QPCR. b The expression levels of AQP5, p38 and p-p38 in control or NFAT5-depleted MEC-1 cells and CLL-B cells were detected through Immunoblot assays. Results are presented as mean ± SD, **p < 0.01
AQP5 Overexpression Reversed the Inhibitory Effect of NFAT5 Depletion on Cell Proliferation
Since AQP5 was regulated by NFAT5, we wonder whether NFAT5’s effect on apoptosis and p38 activation could be abrogated by AQP5 overexpression. As shown in Fig. 5a, reduced BCL-2 and increased cleaved caspase-3 caused by NFAT5 depletion could be reversed by AQP5 overexpression. Inhibition of p38 activation caused by NFAT5 knockdown was reversed by AQP5 overexpression (Fig. 5a). Furthermore, the reduced cell proliferation by NFAT5 knockdown was suppressed by AQP5 overexpression (Fig. 5b). The increased apoptosis resulted from NFAT5 depletion was also suppressed by AQP5 overexpression (Fig. 5c). Taken together, NFAT5 facilitated malignant cell survival by upregulating AQP5.

AQP5 overexpression reversed the inhibitory effect of NFAT5 depletion on cell proliferation. a The expression levels of AQP5, p38, p-p38, BCL-2 and cleaved caspase-3 in control, NFAT5-depleted, AQP5 overexpressed, and NFAT5-depleted plus AQP5 overexpressed MEC-1 cells and CLL-B cells were determined through Immunoblot assays. b BrdU incorporation assay was performed to evaluate the proliferation ability of control, NFAT5-depleted, AQP5 overexpressed, and NFAT5-depleted plus AQP5 overexpressed MEC-1 cells and CLL-B cells. c Flow cytometry assay was performed to evaluate apoptosis in control, NFAT5-depleted, AQP5 overexpressed, and NFAT5-depleted plus AQP5 overexpressed MEC-1 cells and CLL-B cells. Results are presented as mean ± SD, *p < 0.05, ** p < 0.01
STUB1 Regulated NFAT5 Stability
In light of the critical function of NFAT5 in cell survival, we screened for genes that could regulate NFAT5 expression. Bioinformatic analysis indicated STUB1 as a potential binding protein of NFAT5 (http://ubibrowser.ncpsb.org/ubibrowser/home/index) (Fig. 6a). Then we explored whether STUB1 affects the expression of NFAT5. As shown in Fig. 6b, NFAT5 mRNA level remained unchanged upon STUB1 overexpression. However, STUB1 overexpression inhibited NFAT5 protein level in MEC-1 cells (Fig. 6c). Pulldown assay indicated the direct binding interaction between STUB1 and NFAT5 (Fig. 6d). In addition, STUB1 overexpression further inhibited NFAT5 expression under cycloheximide (CHX) treatment (Fig. 6e). Our data indicated STUB1 directly bound to NFAT5 and promoted its degradation.

STUB1 regulated NFAT5 degradation. a Bioinformatic analysis of NFAT5 binding protein. b, c The level of NFAT5 in control or STUB1 overexpressed MEC-1 cells as measured through QPCR and Immunoblot assays. d Pulldown assay showed direct binding between NFAT5 and STUB1. e Immunoblot assay indicated inhibited expression of NFAT5 in STUB1 overexpressed cells. Results are presented as mean ± SD, **p < 0.01
Discussion
CLL is mainly caused by lymphocytic tumor-like proliferation (Hosnijeh et al. 2020). Due to blocked apoptosis and prolonged survival time of small monoclonal lymphocytes, a large number of them gather in bone marrow, blood, lymph nodes and other organs, and the normal hematopoietic function of bone marrow is therefore inhibited, eventually leading to bone marrow failure, anemia, and ischemia (Gopalakrishnan et al. 2020). CLL develops slowly and the average survival time from diagnosis is about 4–6 years. To combat this disease, targeted therapy is a promising strategy, and more therapeutic targets needs to be developed (Hashemi et al. 2020; Karabon et al. 2020). Herein, we noticed a transcription factor, NFAT5, was highly expressed in human CLL cells. Our data further confirmed that NFAT5 affected the proliferation and apoptosis of CLL cells in vitro. Therefore, we concluded that NFAT5 could serve as a promising therapeutic target for CLL treatment.
NFAT5 was widely involved in the progression and metastasis of multiple types of tumors (Germann et al. 2012; Guo and Jin 2015; Wang et al. 2013). It was reported that NFAT5 contributed to the glycolytic phenotype rewiring and pancreatic cancer progression via activating the expression of PGK1 (Jiang et al. 2019). Additionally, a miRNA, miR-194, suppressed high glucose-induced non-small cell lung cancer cell (NSCLC) progression via targeting NFAT5 (Meng et al. 2019). Another study indicated that NFAT5 contributed to the metastasis of murine melanoma (Kim et al. 2018). In addition, NFAT5 was also involved in regulating the progression of a variety of tumors, including colorectal cancer and breast cancer (Amara et al. 2016; Guo and Jin 2015). We here found the key role of NFAT5 in the progression of CLL. Through series of in vitro assays, we found that NFAT5 promoted CLL progression via affecting cell proliferation and apoptosis. These studies, together with our findings, suggest targeting NFAT5 would be a promising anti-tumor strategy.
Furthermore, we found that NFAT5 regulated AQP5 expression and the phosphorylation of p38 MAPK in CLL cells. Our data suggested that the NFAT5/AQP5/p38 MAPK axis acted a critical role in CLL progression. AQP5 was involved in the progression of leukemia. Previous study showed that AQP5 was overexpressed in chronic myelogenous leukemia cells and played a role in promoting cell proliferation and inhibiting apoptosis, consistent with our study (Trama et al. 2000). MAPK pathway was also critical in the progression and metastasis of CLL. The inhibition of MEK1/2, a member in the MAPK pathway, affected the actions of Venetoclax and ABT-737 under conditions that mimic the CLL tumor microenvironment (Zhou et al. 2020). Recent findings have also shown aberrant activation of MAPK signaling and its interactions with other pathways in the pathogenesis of CLL (Zhou et al. 2011). The inhibitory effect of NFAT5 on cell proliferation could be partially reversed by AQP5 overexpression, suggesting the existence of other mechanisms mediating the regulation of NFAT5. We found that NFAT5 participated in the progression of CLL partially via MAPK pathway, and further confirmed the key effects of MAPK pathway on CLL. Notably, the results show that AQP5 overexpression does not completely reverse the effect of NFAT5 knockdown, suggesting that NFAT5 has a regulatory mechanism involving factors other than AQP5. Interestingly, previous studies provided the evidence that increased NFAT5 expression stimulated transcription of Hsp70 in preeclamptic placentas (Park et al, 2014). NFAT5 also mediated oral cancer cell proliferation by inducing the EGFR translocation through increasing the expression of DPAGT1, an essential enzyme for catalyzing the first committed step of N-linked protein glycosylation (Yoshimoto et al. 2020).
Notably, NFAT5 serves as a transcription factor and affects different cellular functions. NFAT5 protected against hypertonicity through structuring of its intrinsically disordered domain (Ito et al. 2007). We would examine the role of the disordered domain of NFAT5 in CLL progression in future studies. Additionally, Titanium dioxide nanoparticles induced RAW264.7 macrophage apoptosis through miR-29b-3p/NFAT5 axis, which suggested that NFAT5 affected inflammatory responses and might therefore affect CLL (Xu et al. 2020a).
In conclusion, we found that NFAT5 was highly expressed in CLL cells. Our data further confirmed that NFAT5 promoted proliferation and suppressed apoptosis of CLL cells in vitro. Further, we found that NFAT5 facilitated malignant cell survival and p38 MAPK activation by upregulating AQP5, and therefore promoted the progression of CLL. We also found that STUB1 directly bound to NFAT5 and promoted its degradation. We therefore propose that NFAT5 could serve as a promising therapeutic target for CLL treatment.
Data Availability
All data generated or analyzed during this study are included in this published article.
References
Amara S, Alotaibi D, Tiriveedhi V (2016) NFAT5/STAT3 interaction mediates synergism of high salt with IL-17 towards induction of VEGF-A expression in breast cancer cells. Oncol Lett 12:933–943. https://doi.org/10.3892/ol.2016.4713
Barthel R et al (2020) CD74 is dispensable for development of chronic lymphocytic leukemia in Emicro-TCL1 transgenic mice. Leuk Lymphoma. https://doi.org/10.1080/10428194.2020.1791851
Chae YK et al (2008) Human AQP5 plays a role in the progression of chronic myelogenous leukemia (CML). PLoS ONE 3:e2594. https://doi.org/10.1371/journal.pone.0002594
Fernandez-Castillo N et al (2015) Transcriptomic and genetic studies identify NFAT5 as a candidate gene for cocaine dependence. Transl Psychiatry 5:e667. https://doi.org/10.1038/tp.2015.158
Germann S et al (2012) Dual role of the ddx5/ddx17 RNA helicases in the control of the pro-migratory NFAT5 transcription factor. Oncogene 31:4536–4549. https://doi.org/10.1038/onc.2011.618
Gopalakrishnan S et al (2020) Integrated mechanistic model of minimal residual disease kinetics with venetoclax therapy in chronic lymphocytic leukemia. Clin Pharmacol Ther. https://doi.org/10.1002/cpt.2005
Guo K, Jin F (2015) NFAT5 promotes proliferation and migration of lung adenocarcinoma cells in part through regulating AQP5 expression. Biochem Biophys Res Commun 465:644–649. https://doi.org/10.1016/j.bbrc.2015.08.078
Halterman JA, Kwon HM, Leitinger N, Wamhoff BR (2012) NFAT5 expression in bone marrow-derived cells enhances atherosclerosis and drives macrophage migration. Front Physiol 3:313. https://doi.org/10.3389/fphys.2012.00313
Hashemi M, Mohammadipour M, Rostami S, Soltanpour MS (2020) Promoter DNA methylation frequency and clinicopathological role of miR-129–2 gene in patients with chronic lymphocytic leukemia. Oman Med J 35:e151. https://doi.org/10.5001/omj.2020.71
Hosnijeh FS et al (2020) Proteomic markers with prognostic impact on outcome of chronic lymphocytic leukemia patients under chemo-immunotherapy; results from the HOVON 109 study. Exp Hematol. https://doi.org/10.1016/j.exphem.2020.08.002
Ito T, Fujio Y, Takahashi K, Azuma J (2007) Degradation of NFAT5, a transcriptional regulator of osmotic stress-related genes, is a critical event for doxorubicin-induced cytotoxicity in cardiac myocytes. J Biol Chem 282:1152–1160. https://doi.org/10.1074/jbc.M609547200
Jiang Y et al (2019) Transcription factor NFAT5 contributes to the glycolytic phenotype rewiring and pancreatic cancer progression via transcription of PGK1. Cell Death Dis 10:948. https://doi.org/10.1038/s41419-019-2072-5
Karabon L et al (2020) Abnormal expression of BTLA and CTLA-4 immune checkpoint molecules in chronic lymphocytic leukemia patients. J Immunol Res 2020:6545921. https://doi.org/10.1155/2020/6545921
Kim DH, Kim KS, Ramakrishna S (2018) NFAT5 promotes in vivo development of murine melanoma metastasis. Biochem Biophys Res Commun 505:748–754. https://doi.org/10.1016/j.bbrc.2018.09.171
Koch M et al (2020) Meta-analysis reveals significant sex differences in chronic lymphocytic leukemia progression in the emicro-TCL1 transgenic mouse model. Cancers. https://doi.org/10.3390/cancers12071980
Kumar R, DuMond JF, Khan SH, Thompson EB, He Y, Burg MB, Ferraris JD (2020) NFAT5, which protects against hypertonicity, is activated by that stress via structuring of its intrinsically disordered domain. Proc Natl Acad Sci USA. https://doi.org/10.1073/pnas.1911680117
Kuper C, Beck FX, Neuhofer W (2014) NFAT5-mediated expression of S100A4 contributes to proliferation and migration of renal carcinoma cells. Front Physiol 5:293. https://doi.org/10.3389/fphys.2014.00293
Lee JH, Kim JW, Im YS, Seong GJ, Lee HK (2011) Cyclosporine a induces nerve growth factor expression via activation of MAPK p38 and NFAT5. Cornea 30(Suppl 1):S19–S24. https://doi.org/10.1097/ICO.0b013e3182281028
Meng X, Li Z, Zhou S, Xiao S, Yu P (2019) miR-194 suppresses high glucose-induced non-small cell lung cancer cell progression by targeting NFAT5. Thorac Cancer 10:1051–1059. https://doi.org/10.1111/1759-7714.13038
Park J et al (2014) Increased NFAT5 expression stimulates transcription of Hsp70 in preeclamptic placentas. Placenta 35:109–116. https://doi.org/10.1016/j.placenta.2013.12.005
Qin X, Wang Y, Li J, Xiao Y, Liu Z (2017) NFAT5 inhibits invasion and promotes apoptosis in hepatocellular carcinoma associated with osmolality. Neoplasma 64:502–510. https://doi.org/10.4149/neo_2017_403
Remo A et al (2015) Systems biology analysis reveals NFAT5 as a novel biomarker and master regulator of inflammatory breast cancer. J Transl Med 13:138. https://doi.org/10.1186/s12967-015-0492-2
Sedlarikova L, Petrackova A, Papajik T, Turcsanyi P, Kriegova E (2020) Resistance-associated mutations in chronic lymphocytic leukemia patients treated with novel agents. Front Oncol 10:894. https://doi.org/10.3389/fonc.2020.00894
Trama J, Lu Q, Hawley RG, Ho SN (2000) The NFAT-related protein NFATL1 (TonEBP/NFAT5) is induced upon T cell activation in a calcineurin-dependent manner. J Immunol 165:4884–4894. https://doi.org/10.4049/jimmunol.165.9.4884
Villanueva S et al (2012) NFAT5 is activated by hypoxia: role in ischemia and reperfusion in the rat kidney. PLoS ONE 7:e39665. https://doi.org/10.1371/journal.pone.0039665
Wang Q, Zhou Y, Rychahou P, Liu C, Weiss HL, Evers BM (2013) NFAT5 represses canonical Wnt signaling via inhibition of beta-catenin acetylation and participates in regulating intestinal cell differentiation. Cell Death Dis 4:e671. https://doi.org/10.1038/cddis.2013.202
Xu S, Sui J, Fu Y, Wu W, Liu T, Yang S, Liang G (2020a) Titanium dioxide nanoparticles induced the apoptosis of RAW264.7 macrophages through miR-29b-3p/NFAT5 pathway. Environ Sci Pollut Res Int 27:26153–26162. https://doi.org/10.1007/s11356-020-08952-5
Xu Z, Sun Y, Wei Z, Jiang J, Xu J, Liu P (2020b) Suppression of CXCL-1 could restore necroptotic pathway in chronic lymphocytic leukemia. OncoTargets Ther 13:6917–6925. https://doi.org/10.2147/OTT.S256993
Yoshimoto S, Morita H, Matsuda M, Katakura Y, Hirata M, Hashimoto S (2020) NFAT5 promotes oral squamous cell carcinoma progression in a hyperosmotic environment. Lab Investig. https://doi.org/10.1038/s41374-020-00486-1
Zhou X, Izumi Y, Burg MB, Ferraris JD (2011) Rac1/osmosensing scaffold for MEKK3 contributes via phospholipase C-gamma1 to activation of the osmoprotective transcription factor NFAT5. Proc Natl Acad Sci USA 108:12155–12160. https://doi.org/10.1073/pnas.1108107108
Zhou X et al (2020) Anti-breast cancer effect of 2-dodecyl-6-Methoxycyclohexa-2,5-diene-1,4-dione in vivo and in vitro through MAPK signaling pathway. Drug Design Devel Ther 14:2667–2684. https://doi.org/10.2147/DDDT.S237699
Author information
Authors and Affiliations
Contributions
BLC and YL designed the study, supervised data collection, SX and YN analyzed the data, interpreted the data, JZ prepare the manuscript for publication and reviewed the draft of the manuscript. All authors have read and approved the manuscript.
Corresponding author
Ethics declarations
Conflict of Interest
The authors state that there are no conflicts of interest to disclose.
Ethical Approval
Ethical approval was obtained from the Ethics Committee of Affiliated Hospital of Guilin Medical University.
Informed Consent
Written informed consent was obtained from legally authorized representative(s) for anonymized patient information to be published in this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Chen, B.L., Li, Y., Xu, S. et al. NFAT5 Regulated by STUB1, Facilitates Malignant Cell Survival and p38 MAPK Activation by Upregulating AQP5 in Chronic Lymphocytic Leukemia. Biochem Genet 59, 870–883 (2021). https://doi.org/10.1007/s10528-021-10040-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10528-021-10040-3