
Abstract
Protein kinase CK2 sustains acute myeloid leukemia cell growth, but its role in leukemia stem cells is largely unknown. Here, we discovered that the CK2 catalytic α and regulatory β subunits are consistently expressed in leukemia stem cells isolated from acute myeloid leukemia patients and cell lines. CK2 inactivation with the selective inhibitor CX-4945 or RNA interference induced an accumulation of leukemia stem cells in the late S–G2–M phases of the cell cycle and triggered late-onset apoptosis. As a result, leukemia stem cells displayed an increased sensitivity to the chemotherapeutic agent doxorubicin. From a molecular standpoint, CK2 blockade was associated with a downmodulation of the stem cell-regulating protein BMI-1 and a marked impairment of AKT, nuclear factor-κB (NF-κB) and signal transducer and activator of transcription 3 (STAT3) activation, whereas FOXO3a nuclear activity was induced. Notably, combined CK2 and either NF-κB or STAT3 inhibition resulted in a superior cytotoxic effect on leukemia stem cells. This study suggests that CK2 blockade could be a rational approach to minimize the persistence of residual leukemia cells.
Similar content being viewed by others


Introduction
Acute myeloid leukemia (AML) represents a therapeutic challenge because of the high relapse rate.1, 2 AML is hierarchically organized in a minority of leukemia stem cells (LSCs) and the remaining bulk of more differentiated AML blasts.3, 4 Similar to normal hematopoietic stem cells (HSCs), LSCs display a low proliferation rate, being thus inherently resistant to commonly employed cell cycle-dependent chemotherapeutic agents. Therefore, the identification of molecular mechanisms that regulate LSC proliferation and survival could help develop LSC-specific targeted therapies.5
CK2 is a serine/threonine kinase formed by two catalytic (α or α') subunits and one regulatory (β) subunit that assemble in a tetramer.6 Recently, this kinase has gained growing importance in the pathogenesis of a variety of blood tumors. A ‘non oncogene addiction’ mechanism, through which CK2 regulates cell growth enhancing the oncogenic potential of critical signaling cascades, has been proposed.7, 8 For instance, CK2 has a prominent role in nuclear factor-κB (NF-κB), signal transducer and activator of transcription 3 (STAT3) and AKT pathways. It regulates the degradation of IκBα by direct phosphorylation in the C-terminal PEST domain,9 boosts the activation of IKKα and IKKβ kinases and phosphorylates NF-κB p65 (RelA) directly on S529, likely enhancing its transcriptional activity.10 Similarly, CK2 regulates the activation of JAK1 (Janus kinase 1) and JAK2 (Janus kinase 2) upstream of STAT1, STAT5 and STAT3 and it likely controls the phosphorylation of this latter protein on S727.11 CK2 also inhibits PTEN (phosphatase and tensin homolog)12 and triggers AKT phosphorylation on S129.13 All these pathways play a pivotal role in AML/LSC biology, regulating AML maintenance, LSC renewal and proliferation.14, 15, 16, 17, 18 To note, a peculiar scenario marks the AKT/FOXO pathway in LSCs. Although FOXO transcription factors have an antiproliferative function that is inhibited by AKT-mediated phosphorylation,19 in leukemogenesis the role of AKT could be paradoxically antagonistic to the maintenance of LSCs. In fact, either AKT activation or deletions of FOXO1/3/4 were described to impair leukemia cell growth in a mouse model of MLL-AF9 allele-driven AML.20 Similarly, it has been shown that imatinib-induced downregulation of BCR-ABL in chronic myelogenous leukemia leads to a reduction of AKT activation and subsequent stimulation of FOXO3a that in turn sustains leukemia-initiating cell proliferation.21 Thus, the consequences of AKT inhibition on LSC expansion have to be carefully evaluated.
Nevertheless, despite others and our group demonstrating that CK2 may play a role in AML blasts,22, 23 its function in LSC-associated pathways has been poorly investigated.24 In the present study, employing LSCs obtained from AML patients and AML cell lines Kasumi-1 and KG1a, we analyzed CK2 expression levels and the consequences of its inactivation on AML/LSC growth and on NF-κB, STAT3 and AKT/FOXO signaling pathways. CK2 resulted to be critical for the sustenance of these signaling cascades and LSC survival, regulating the balance between apoptosis and cell cycle and, consequently, self-renewal and sensitivity to a chemotherapeutic agent like doxorubicin. Moreover, CK2 blockade increased the efficacy of NF-κB and STAT3 inhibitors, profoundly affecting LSC viability.
Materials and methods
Patients and cell lines
Peripheral blood and bone marrow specimens were collected from 22 patients with a newly diagnosed AML after informed consent. Procedures followed the rules indicated in the Declaration of Helsinki. The project outline and consent procedures were submitted and approved by the Ethic Committee of the Padova University Hospital (protocol number 2612P). Only samples with >60% of CD34+ blasts cells were used. Mononuclear cells were isolated through Ficoll-Hypaque (GE Healthcare, Milan, Italy) centrifugation and cell viability was assessed by Trypan blue exclusion assay. Healthy adult bone marrow CD34+ cells used as normal controls for western blot (WB) and kinase assay were obtained from Lonza Ltd (Walkersville, MD, USA). AML Kasumi-1 cell line was purchased from DSMZ (Braunschweig, Germany). KG1a cell line was a kind gift of Dr Ornella Parolini (E Menni Research Center (CREM), Brescia, Italy).
Kinase activity in cell lysates
CK2 activity was determined using ‘Casein Kinase 2 Assay Kit’ (Millipore, Temecula, CA, USA) on 2.5 μg of whole-cell lysate following the manufacturer’s instruction. CK2 activity was in a linear range according to the lysate protein amount used for the assay. The synthetic peptide used as substrate and provided by the kit has the following sequence: RRRDDDSDDD.
Lentiviral transduction
Kasumi-1 cells were transduced with IPTG (isopropyl-β-D-thiogalactoside)-inducible lentiviral particles carrying CK2α-directed small hairpin RNA (shRNA; pLKO_IPTG_3XLacO, Sigma, Milan, Italy), coupled to a puromycin resistance cassette, according to the manufacturer’s instructions. Cells were infected with a multiplicity of infection of 10. Transfected cells were selected adding puromycin (0.5 μg/ml) to the culture medium. Selected Kasumi-1 clones were then induced with 500 μM of IPTG every 3 days for a total induction period of 2 weeks, the lapse of time in which we obtained the best results in terms of CK2α knockdown efficacy. Three independent CK2α shRNA sequences were tested and finally the vector clone TRCN0000320858 with the shRNA sequence 5′-CAATCCCGGCTGCTGCATTTA-3′ was chosen.
Statistical analysis
Data were evaluated for their statistical significance with the two-tail paired Student’s t-test. Values were considered statistically significant at P-values below *0.05, **0.01 and ***0.001.
See Supplementary Information for other methods.
Results
CK2 expression in AML LSCs
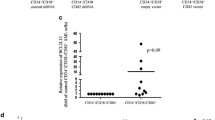
Analysis of mRNA expression of the CK2 subunits in the LSC-enriched fraction (Lin− CD34+ CD38−) from 18 primary AML samples and two AML cell lines, Kasumi-1 and KG1a, (Figure 1a shows the fluorescence-activated cell sorting (FACS) strategy; see Table 1 for clinical and pathological features of the patients) showed a substantial overexpression of CK2α mRNA in 15/18 samples compared with a pool of normal CD34+ HSCs, in most cases higher than twofold (Figure 1b, top graph). CK2β mRNA was expressed at high levels in 13/18 patients (Figure 1b, bottom graph). In AML cell lines, quantitative real-time PCR clearly demonstrated an overexpression of CK2α in both subpopulations of the two cell lines in the study (Figure 1c, top graph), whereas CK2β was overexpressed only in KG1a cells (Figure 1c, bottom graph). In addition, WB and densitometric analyses, performed in the LSC-enriched population and in the Lin− CD34+ CD38+ non-stem cell fraction from three patients, proved a consistent expression of CK2α and CK2β in both populations (Figure 1d). However, CK2 kinase activity showed a remarkable variability among patients: it was found high in 2/4 samples (patients 12 and 20), and similar to the control HSC in two other lysates (14 and 18) (Figure 1e). Interestingly, WB and densitometric analyses showed a clear upregulation of both subunits in Kasumi-1 and KG1a subpopulations (Figure 1f), consistent with the high kinase activity observed for these cells (Figure 1g).

CK2α and CK2β expression in LSCs purified from AML patients and cell lines. (a) Representative dot plots of one AML patient sample (pt 18), and cell lines Kasumi-1 and KG1a stained with anti-Lineage (anti-CD2, CD3, CD14, CD15, CD16, CD19, CD20, Glycophorin A), anti-CD34 and anti-CD38 antibodies. The gate was set on Lineage-negative (Lin−) cells (left panel) and the analysis of CD34 and CD38 expression inside the Lin− population was then performed (right panel). LSCs (34+ 38−) and non-LSCs (34+ 38+) were selected through FACS. (b) CK2α and CK2β mRNA expression was determined by qRT-PCR in sorted LSC samples from patients and (c) cell lines; the internal calibrator was represented by cDNA derived from a pool of HSCs purified from the bone marrow of healthy donors; GAPDH was the reference gene; n=3, ***P<0.001. (d) WB and corresponding densitometric analysis of CK2α and CK2β protein levels in three AML patients comparing LSC and non-LSC populations with healthy HSCs. GAPDH was used as loading control. (e) CK2 kinase activity assay in LSC and CD34+CD38+ AML patient cells, measured against a specific synthetic peptide: data are reported as normalized counts per minute (CPM). (f) WB and corresponding densitometric analysis of CK2α and CK2β protein levels in Kasumi-1 and KG1a cell lines comparing LSC and non-LSC populations with healthy HSCs. GAPDH was used as loading control. The threshold set in the densitometric analysis (dot line) corresponds to normalized LSCs protein expression. (g) CK2 kinase activity assay in both populations of AML cell lines measured against a specific synthetic peptide: data are reported as CPM; n=3, ***P<0.001 compared with healthy HSCs; ##P<0.01 compared with LSC counterpart. pt, patient; qRT-PCR, quantitative real-time PCR; 34+ 38+, Lin−/CD34+/CD38+ cells.
CK2 blockade with the highly selective ATP-competitive inhibitor CX-4945 is cytotoxic to the AML LSC population
To test the role of CK2 in LSC-enriched versus non-LSC-enriched AML fractions, in a first set of experiments, we employed the ATP-competitive CK2 inhibitor CX-4945.25, 26 Freshly purified cells from AML patients were treated for 18 h and FACS analysis of Annexin V-stained LSCs and CD34+ CD38+ cells was then performed (Figure 2a). The concentrations of CX-4945 (5 μM) were previously optimized (data not shown). CX-4945 caused apoptosis in LSCs of all the patients studied, even if to different extents. On the non-LSC fraction the effects of CX-4945 were similar (Figure 2b). To note, in two of the four patients tested, patients 12 and 14, who displayed an adverse and an intermediate-I ELN (European LeukemiaNet) genetic risk, respectively (see Table 1), cell death was much higher for LSCs than for the CD34+ CD38+ population (Figure 2b). However, patients 11 and 17 also had adverse risk, indicating that the sensitivity to CK2 inhibition could not be influenced by the ELN genetic risk. The effects of CK2 blockade were then tested on the AML Kasumi-1 and KG1a cells: roughly 40% of Lin− Kasumi-1 cells display a LSC phenotype and 60% are CD34+ CD38+ and for KG1a Lin− cells the percentages are ∼80% LSCs and 20% CD34+ CD38+ (Figure 1a).27 CK2 blockade for 18 h with CX-4945 caused substantial apoptosis of LSC-enriched and CD34+ CD38+ cells (Figures 2c and e) that was even more pronounced after 36 h of treatment (Figures 2d and f). At early time points, LSCs appeared to be more sensitive to the treatment compared with the non-LSC counterpart, even at lower concentrations of CX-4945, reinforcing the results obtained in AML patient samples. Apoptosis was also confirmed by WB analysis showing an increase of PARP (poly (ADP-ribose) polymerase) cleavage that was markedly higher after addition of CX-4945 (5 μM) for both 18 and 36 h (Figures 2g and h). Thus, this result highlights the efficacy of CX-4945 in promoting apoptosis of the whole leukemic population, affecting both LSCs and CD34+ CD38+ cells. As some patients displayed similar CK2 protein levels and kinase activity (Figures 1d and e) when compared with normal HSCs, we wanted to exclude that CX-4945 also had an effect on healthy HSCs. To this aim, we used healthy HSCs from two adult donors (for more details see Supplementary Information). Cells were treated with CX-4945 5 μM for 18 h and viability was assessed through Annexin V staining restricting the analysis in the gate of Lin−CD34+ cells (Supplementary Figures 1a and 1b). In both samples the inhibition of CK2 did not cause any significant change in HSC viability.

Effects of CK2 inhibition on LSC and CD34+ CD38+ cell survival. (a) Representative dot plot graphs of one primary AML sample untreated (UN) (upper panel) or treated with CX-4945 5 μM (lower panel): Annexin V (AnnV) positivity was determined in both CD38+ and CD38− cells in the gate of CD34+ inside the Lin− population. (b) Histograms representing the quantification of AnnV-positive cells in LSCs (upper panel) and CD34+ CD38+ subfraction (lower panel) from four AML patients untreated or treated with CX-4945 5 μM; data are reported as normalized values over the untreated samples. (c) Histogram plots of AnnV staining and FACS analysis of purified populations from Kasumi-1 and KG1a cell lines untreated or exposed to different concentrations of CX-4945 for 18 h and (d) 36 h. (e) Graph summarizing total dead cells (apoptotic, AnnV+, and necrotic, PI+ cells) within the LSC and CD34+ CD38+ subfractions (normalized data over untreated samples) upon treatment with CX-4945 for 18 h and (f) 36 h. (g) WB analysis of PARP cleavage upon treatment with CX-4945 for 18 h and (h) 36 h. Figures (c) to (h) represent three independent experiments. The values are shown as mean values±s.d. *P<0.05, **P<0.01 and ***P<0.001. In WB, GAPDH was used as loading control.
CK2 inactivation affects cell cycle progression of LSCs and non-LSCs
CK2 controls the cell cycle, sustaining cell proliferation.28, 29, 30 Therefore, we sought to check what could be the effects of CK2 inhibition on LSC and non-LSC cell cycle progression. To this aim, LSC-enriched and CD34+ CD38+ populations, purified from Kasumi-1 or KG1a cell lines, were treated for 18 and 36 h with CX-4945 (5 μM) and cell cycle was analyzed either with propidium iodide or with 5-bromodeoxyuridine (BrdU) staining. At later time points (36 h) cell cycle analysis was also assessed with Ki-67 staining that allows the identification of quiescent cells in the G0 phase (Ki67 negative). Surprisingly, CK2 blockade was associated with the accumulation of cells in S and M phases in both LSCs and CD34+ CD38+ populations, reducing the amount of cells in quiescent G0 and G1 phases (Figure 3a). At 18 h, inhibition of CK2 also caused an accumulation of cells in sub-G0/G1 phase indicating DNA fragmentation likely associated with incipient apoptosis. The increased number of BrdU-positive cells also suggested the accumulation of cycling cells (Figure 3b). After 36 h, a similar behavior could be observed, although the amount of cells in S and M phases was lower, and the frequency of cells in sub-G0/G1 was higher as compared with 18 h (Figure 3c). The analysis performed with Ki-67/propidium iodide staining confirmed the decline in cell proliferation at this later time point (Figure 3d). Thus, these results clearly show that a blockade of CK2 prompts leukemia cells to accumulate in the late phases of the cell cycle and, later on, to undergo apoptosis.

Effects of CK2 inhibition on cell proliferation in AML cell lines. (a) Evaluation of cell cycle progression by propidium iodide (PI) staining and flow cytometry of Kasumi-1 and KG1a cells upon treatment with CX-4945 for 18 h: (a, left panel) Representative peaks of PI incorporation and (a, right panel) corresponding histograms summarizing the average percentages of cells in the different cell cycle phases (sub-G0/G1, G1, S, G2/M) (mean values±s.d.; n=6; *P<0.05, **P<0.01 and ***P<0.001). (b) BrdU and PI staining of Kasumi-1 cells treated with CX-4945 5 μM for 18 h: dot plots are shown on the left, and graphs representing BrdU-positive cells on the right (mean±s.d.; n=6; *P<0.05. (c) PI staining and FACS analysis of Kasumi-1 and KG1a cells after exposition to CX-4945 for 36 h. (c, left panel) Representative peaks of PI intensity and (c, right panel) the histogram showing the average of percentages (mean values±s.d.; n=6; *P<0.05, **P<0.01 and ***P<0.001). (d) Staining of Kasumi-1 cells with anti Ki-67 antibody and PI and FACS analysis after 36 h of CK2 blockade with CX-4945 5 μM: on the left, the contour plots show the three main cell cycle phases identifiable with this approach: in Q3 panel sub-G0 and G0 (Ki-67-/PIlow) phase; in Q1 panel G1 (Ki-67+/PIlow) phase; and in Q2 panel S-G2-M (Ki67+/PIhigh) phase. On the right the corresponding histograms of cell phase distribution are shown (mean values±s.d.; n=3, *P<0.05 and **P<0.01). Data are representative of three independent experiments.
CK2 blockade turns off NF-κB, STAT3 and AKT, reduces BMI-1 expression and promotes accumulation of FOXO3a
To gain insights on how CK2 blockade can elicit cell cycle progression while triggering apoptosis in LSC-enriched and Lin− CD34+ CD38+ cells, we investigated the NF-κB, STAT3 and AKT/FOXO pathways. As shown in Supplementary Figure 2a, in primary AML blasts from patients (n=4) (top panels) and in the cell lines Kasumi-1 and KG1a (bottom panels) we observed AKT phosphorylation in S473 in LSCs and non-LSCs counterpart. The AKT-dependent inhibitory phosphorylation of FOXO3a in S253 was also detected. To note, the FOXO3-regulated target FOXO1 protein was strongly expressed in Kasumi-1 cells, whereas it was found much less abundant in KG1a cells, indicating a variable pattern. Moreover, NF-κB p65 and STAT3 were found phosphorylated in S529 and S727, respectively, indicating their activation. To note, the CD34+ CD38+ AML cell population isolated from patients displayed higher levels of all these phosphorylated proteins compared with the LSC fraction. We next evaluated the mRNA levels of Polycomb family protein BMI-1, which is essential for LSC maintenance.31, 32 BMI-1 mRNA was considerably overexpressed in 15 of 18 LSC patient samples, with a range of increase between 2-fold and 10-fold compared with normal HSCs (Supplementary Figure 1b, upper-left panel). Intriguingly, BMI-1 was found equally expressed in Kasumi-1 relative to HSCs and overexpressed in both populations of KG1a (Supplementary Figure 2b, upper-right panel). Furthermore, expression levels of FOXO3a transcription factor, target of the PI3K/AKT pathway, were analyzed. FOXO3a mRNA was overexpressed in 9 out of 18 LSC-enriched samples, but it was downmodulated in both cell lines (Supplementary Figure 2b, middle panels). We also looked at the mRNA levels of the FOXO3a transcriptional target FOXO1,33 which was downmodulated in both cell lines and substantially expressed in only 6 of 18 patient samples and without any obvious correlation with FOXO3a levels (Supplementary Figure 2b, lower panels). The significance of these findings remains to be elucidated. Next, LSCs and CD34+ CD38+ cells sorted from Kasumi-1 and KG1a cell lines (Figure 4) were treated for 9 or 18 h with CX-4945. As shown in Figure 4a, a dose-dependent reduction in AKT phosphorylation in S473 was observed in both populations, already at 9 h after treatment. CK2 inhibition caused a slight decrease in AKT expression that was more evident in KG1a cell line. The AKT target FOXO3a was found highly phosphorylated on S253 in untreated samples in both LSC-enriched and CD34+ CD38+ cells. Remarkably, CK2 inhibition caused a clear reduction of FOXO3a S253 phosphorylation. Furthermore, after CK2 blockade, an increase in the total amount of FOXO3a could be detected. Likewise, in CK2-inhibited cells the FOXO3a target FOXO1 was increased (Figure 4a), with a similar dose-dependent trend of accumulation preceded by a parallel increase of its mRNA levels within 18 h of CK2 inhibition (Figure 4b). We also studied NF-κB p65 and STAT3 activating phosphorylation after 18 h of treatment with CX-4945 (Figure 4c). NF-κB p65 phosphorylation in S529 was markedly reduced in both LSCs and non-LSCs (Figure 4c). The blockade of CK2 caused a similar reduction of STAT3 phosphorylation on S727 (Figure 4c), confirming our previous findings.23 Similar molecular investigations were performed on LSCs purified from one AML patient (patient 22) after exposure to CX-4945 for 18 h. WB analysis also confirmed in this sample the reduction of AKT, FOXO3a, STAT3 and NF-κB p65 phosphorylation upon CK2 blockade (Supplementary Figure 3a) as well as the increase in FOXO3a and FOXO1 mRNA levels (Supplementary Figure 3b).

Effects of CK2 inactivation on leukemia-associated signaling pathways. (a) WB analysis of AKT, FOXO3a and FOXO1 levels in Kasumi-1 (left panel) and KG1a (right panel) sorted LSCs and CD34+ CD38+ population after 9 or 18 h of CK2 inhibition with 2.5 or 5 μM CX-4945. (b) Quantitative real-time PCR (qRT-PCR) of FOXO1, FOXO3a and BMI-1 after 18 h of treatment with CX-4945 in Kasumi-1 and KG1a LSCs. Data are representative of three experiments (mean values±s.d.; *P<0.05, **P<0.01 and ***P<0.001). (c) Protein expression of NF-κB (p65), STAT3 and CD44 in LSCs and non-LSCs of Kasumi-1(left panel) or KG1a (right panel) upon 18 h of exposure to CX-4945. (d) Immunofluorescence analysis of Kasumi-1 (left) and KG1a (right) LSCs stained with anti-FOXO3a antibody after 18 h of treatment with CX-4945 5 μM. Blue: DAPI (4',6-diamidino-2-phenylindole); green: FOXO3a. Images were collected by means of confocal microscope acquiring 10 slices along the z-stack for a total thickness of 10 μM and are represented as the maximum projection. (e) WB of nuclear extracts purified from Kasumi-1 and KG1a LSCs after exposure to CX-4945 for 18 h. GAPDH and PARP were used to assess the purity of the nuclear lysate, and β-actin was used as loading control. (f) WB of BMI-1 in LSC cell lysates from both cell lines after 18 or 36 h of CK2 chemical inhibition. (g) WB showing the expression of CK2α, AKT, FOXO3a, NF-κB (p65) and STAT3 in anti-CK2α iIPTG-inducible shRNA-bearing Kasumi-1 LSCs after treatment with 500 μM of IPTG (see Materials and methods). (h) Analysis of CK2α, BMI-1, FOXO1 and FOXO3a gene expression in the IPTG-induced Kasumi-1 clones. In WB, GAPDH was used as loading control. The graphs related to gene expression report mean values±s.d.; n=3; *P<0.05 and **P<0.01; data were normalized over GAPDH and samples not induced with IPTG.
Intriguingly, CD44 protein, whose expression has been linked to NF-κB and STAT3 activity in AML,34, 35, 36 was also downregulated upon CK2 blockade after 18 h of treatment (Figure 4c). Moreover, confocal immunofluorescence analysis demonstrated in both cell lines that although at baseline conditions FOXO3a was more abundant in the cytoplasm, after treatment with CX-4945 this factor shuttled into the nucleus (Figure 4d). WB of nuclear extracts confirmed the redistribution of FOXO3a and the absence of activated AKT in the nuclear compartment, indicating a reduction of inhibitory mechanism against FOXO3a (Figure 4e). Of note, BMI-1 underwent a clear downmodulation after 36 h of CK2 inhibition (Figure 4f). Most importantly, the changes in the levels of phospho-AKT S473, phospho-FOXO3a S253, phospho-NF-κB p65 S529 and phospho-STAT3 S727 were also detected in IPTG-induced Kasumi-1 clones carrying CK2α-specific lentiviral shRNA, excluding in this way the possibility that they could have been caused from off-target effects of CX-4945 (Figure 4g). Similarly, inhibition with CX-4945 or CK2α RNA silencing determined a reduction of BMI-1 mRNA levels, and an increase of FOXO3a and FOXO1 expression (Figures 4b and h). In these experiments, CK2α silencing was confirmed both at mRNA and protein levels with a reduction of CK2α levels of as high as 50% (Figure 4g). To note, IPTG did not induce toxic effects or nonspecific modulation of these molecules in LSCs not infected with lentiviral particles (data not shown).
CK2 blockade enhances the chemosensitivity of LSCs
LSCs are thought to barely respond to chemotherapy because of their low replication rate. Given that CK2 inhibition prompted leukemia cells toward cell cycling, we assayed LSC chemosensitivity upon CK2 block. To this aim, we pretreated Kasumi-1 LSCs with CX-4945 with or without doxorubicin for 18 h. Doxorubicin produced a moderate cytotoxic effects on Kasumi-1 cells, especially in the non-stem cell fraction. However, the combination of doxorubicin with CX-4945 resulted in a higher rate of apoptosis as compared with the single treatments, in particular of LSCs (Supplementary Figures 4a and b). WB analysis demonstrated a decrease of pro-caspase 3 and a downfall of BCL-XL expression (Supplementary Figure 4c). To validate these results, we repeated the same treatment in IPTG-inducible Kasumi-1 clones. After induction with IPTG, cells were treated with doxorubicin for 18 h. Remarkably, analysis of apoptosis revealed a statistically significant higher amount of apoptotic cells in the combination treatment that was more pronounced in the LSC fraction (Supplementary Figures 4d and 4e). Morphological analysis of Kasumi-1 cells also showed clear signs of apoptosis (Supplementary Figure 4f). These results confirmed the trend observed with CX-4945, although to a lower extent, likely because of residual CK2α activity.
CK2 blockade boosts the effects of both NF-κB and STAT3 inhibitors on LSCs
The data shown above suggest that CK2 could regulate AKT, STAT3 and NF-κB in LSCs. Thus, we analyzed whether CK2 blockade could boost the effects of subtoxic concentrations of STAT3 or NF-κB inhibitors. STATTIC is a commercially available STAT3 inhibitor that selectively inhibits the activation, dimerization and nuclear translocation of STAT3,37 whereas BAY 11-7082 is a IκBα kinase inhibitor that represses NF-κB signaling. To this aim, AML LSCs and CD34+ CD38+ fractions isolated from patients and from Kasumi-1 and KG1a cell lines were treated for 18 h with CX-4945 in the presence of BAY 11-7082 (1 or 4 μM) or STATTIC (2.5 μM). A mild increase of cell death was observed after exposure to low concentrations of the single agents BAY or STATTIC. Remarkably, the combination of either one of the two compounds with CX-4945 strongly enhanced apoptosis in both leukemia cellular subfractions of AML cell lines and in patient LSCs (Figures 5a–c). Similarly, morphological examination of Kasumi-1 LSCs revealed cellular alterations such as vacuolization, nuclear condensation and membrane blebbing, especially in the combined treatments (Figure 5d), and WB analysis demonstrated a stronger increase of PARP cleavage in the combined treatments of both cell lines (Figures 5e and f). The clear inhibitory effects of BAY11-7082 and STATTIC on NF-κB p65 and STAT3, respectively, were confirmed by the downregulation of the phosphorylation levels in the respective activatory sites (Figures 5g and h). Strikingly, the combination of CX-4945 and STATTIC also caused a clear reduction of pNF-κB p65 S529, the serine targeted by CK2 itself. Similarly, the combination with BAY11-7082 negatively influenced the phosphorylation of STAT3. Whether this could be because of a crosstalk between the two pathways or off-target effects remains to be investigated.38

Effects of CK2 blockade combined with STAT3 or NF-κB inhibition. Samples from four AML patients and sorted populations (LSCs and CD34+38+) from Kasumi-1 or KG1a cell lines were treated with CX-4945 (5 μM) in combination with subtoxic concentrations of BAY 11-7082 (1 or 4 μM) or STATTIC (2.5 μM). (a) Graph summarizing Annexin V-positive cells in patient samples treated with CX-4945, BAY, STATTIC or combinations of two of these. Annexin V-positive cells were evaluated in the gates of Lin-CD34+CD38- or Lin-CD34+CD38+ populations. (b) Morphological analysis of Kasumi-1 LSCs after cytospin and May Grünwald-Giemsa staining. Black arrows highlight signs of cell apoptosis/necrosis. (c) Graph summarizing the percentages of Annexin V- and propidium iodide (PI)-positive cells in Kasumi-1 LSCs and non-LSC counterpart (mean values±s.d.; n=3, *P<0.05, **P<0.01 and ***P<0.001; *: the value is significant when compared with untreated samples; #: when compared with CX-4945 alone; +: when compared with BAY; or °: when compared with STATTIC alone). (d) WB showing PARP levels. (e) WB analysis of the expression and phosphorylation of NF-κB (p65) and STAT3 in LSCs. In all WB, GAPDH was used as a loading control. (f) Graph summarizing the percentages of Annexin V- and PI-positive cells in KG1a LSCs and non-LSC counterpart (mean values±s.d.; n=3). (g) WB showing PARP levels. (h) WB analysis of the expression and phosphorylation of NF-κB (p65) and STAT3 in LSCs. In all WB, GAPDH was used as a loading control.
Discussion
As demonstrated in previous studies, AML is initiated and sustained by a numerically rare subset of LSCs characterized by the immature CD34+ CD38− phenotype.3, 4 LSCs are less sensitive to chemotherapy and are considered as a reservoir for disease relapses.2
In the present work we have shown a key role for protein kinase CK2 in promoting LSC survival through the modulation of the STAT3, NF-κB and AKT/FOXO signaling pathways. We have also shown that CK2 inactivation makes LSCs more sensitive to the chemotherapeutic drug doxorubicin.
Our work is the first to indicate consistent expression levels of CK2 (of both the α catalytic and β regulatory subunits) in a series of LSC samples isolated from 18 AML patients and the upregulation of this kinase in the CD34+ CD38− subfraction of Kasumi-1 and KG1a AML cell lines. In LSCs, we analyzed the Polycomb family member BMI-1 that is essential for self-renewal/maintenance of HSCs and LSCs,31 as its expression is controlled by the Wnt and Hedgehog pathways that are in turn positively modulated by CK2-dependent phosphorylation of β-catenin, Smoothened, Gli and Dvl.39 BMI-1 resulted to be highly expressed in the majority of the LSC samples purified from AML patients, although there was not a clear correlation between CK2 and BMI-1 mRNA levels, indicating possible different regulatory fates in LSCs. Experiments aimed at inactivating CK2 pointed out the addiction of both LSCs and CD34+ CD38+ cells to the activity of this kinase. The increase in apoptosis observed in both counterparts, upon CK2 blockade, suggests that the chemical inhibitor CX-4945 could be an effective therapeutic approach to affect the whole malignant population. To note, CK2 inhibition does not seem to affect normal HSC viability, despite the similar CK2 protein levels and kinase activity observed in some cases between HSCs and AML leukemic samples from patients. As CK2 is not an oncogene but supports the oncogenic potential of key signaling pathways and prompts the expansion of mutated cell clones,7, 8 its expression and activity levels taken alone are not sufficient to explain a malignant phenotype. Unlike HSCs, malignant cells become more dependent on CK2 through a non-oncogene addiction mechanism and this condition likely makes them more sensitive even to a mild decrease of CK2 activity. From a molecular standpoint CK2 blockade experiments unraveled a function for this kinase in STAT3, NF-κB (p65/RelA), AKT and BMI-1 pathways in LSCs. These molecules were found activated in both CD34+ CD38− and CD34+ CD38+ patient AML cells. Bona fide tumor suppressor FOXO3a, lying downstream of AKT, was reported to be highly expressed in LSCs,18, 40 and the presence of abundant levels of inhibitory phosphorylation in S253 indicates that FOXO3a is often kept inactive and delocalized in the cytoplasm.41 The observation that CK2 inhibition, either with CX-4945 or by RNA interference, produced a downregulation of AKT and FOXO3a phosphorylation causing an elevation of FOXO3a protein levels in LSCs and in the CD34+ CD38+ fraction suggests a prominent role for this kinase in this signaling axis. To note, the increased FOXO3a expression, upon CK2 blockade, was more evident in LSCs as compared with the CD34+ CD38+ counterpart, together with a marked accumulation of this factor in the nuclear compartment. As growing evidence has suggested that FOXO3a may behave as a promoter of leukemia expansion by regulating stem cell maintenance, as shown in CML and AML models,20, 21 the inhibition of CK2 could appear counterproductive in AML therapy. However, against this possibility is the observation that CK2 inhibition with CX-4945 or RNA interference was clearly cytotoxic to LSCs. Moreover, in AML cell lines, the higher CK2 kinase activity observed may indicate a prominent role for this kinase in AML LSCs. Kim et al.22 had shown how an increase in CK2 expression led to a decrease in the portion of cells at the G0–G1 phase in AML non-stem cell lines. Intriguingly, the effects of CK2 inhibition on the cell cycle were somewhat unexpected. We observed a clear increase of the amount of Kasumi-1 cells in the S phase, as early as 18 h after CK2 inhibition (robustly demonstrated with three different techniques), whereas an accumulation of cells in subG0/G1 phases occurred at later time points, denoting an increase of apoptosis. Contrasting results were obtained in the LSC fraction of KG1a. Even though we also detected for these cells a reduction of G0/G1 phases (as previously observed in Kasumi-1), CX-4945 induced a G2/M arrest. Thus, CK2 might function in a dual mode and its cell cycle regulation effect might be cell-type dependent as previously hypothesized by Siddiqui-Jain et al.42 In LSCs in particular, CK2 likely buffers down cell cycle while sustaining their survival. Noteworthy, the clear downmodulation of BMI-1 in LSCs after treatment with CX-4945 or shRNA indicates the loss of self-renewing cells that are otherwise strictly reliant to a quiescent condition. How CK2 controls BMI-1 expression remains unknown; however, as mentioned above, the possibility of the existence of a CK2/Wnt/BMI-1 or CK2/Hedgehog/BMI-1 pathway should be investigated. We speculate that upon CK2 blockade, FOXO3a might be one of the players involved in the increase of cell cycle progression. Indeed, it was reported that FOXO3a was linked to the maintenance and expansion of leukemia-initiating cells.20, 21 Moreover, it was demonstrated that FOXO3a, in physiological conditions, stabilizes Cdt1 protein that is important to favor cell cycle transition from G1 to S phase.43 From a therapeutic perspective, accumulation of cells in S–G2–M upon blockade/knockdown of CK2 could render the leukemia cell DNA more accessible to chemotherapeutic agents such as doxorubicin.44 Finally, our results demonstrate that targeting CK2 can cooperate with potential novel therapies, such as inhibitors of the STAT3 and NF-κB pathways. CD44 is a STAT3/NF-κB target that positively controls multidrug resistance.34, 36 Upon CK2 inhibition, we observed a downmodulation of this molecule, indicating another possible way through which the kinase targeting can render LSCs more sensitive to chemotherapeutic agents. Moreover, the same downregulation of CD44 could also contribute to the progressive loss of LSC pool, as clearly demonstrated in a previous work.36 Based on our data, a model of CK2 activity in LSC is proposed in Supplementary Figure 5. In conclusion, our study set the rationale for testing CX-4945 or other CK2 inhibitors in combination treatments with conventional and/or novel drugs in AML.
References
Dohner H, Weisdorf DJ, Bloomfield CD . Acute myeloid leukemia. N Engl J Med 2015; 373: 1136–1152.
Badar T, Ravandi F . Relapsed acute myeloid leukemia: need for innovative treatment strategies to improve outcome. Clin Lymph Myel Leuk 2015; 15: S104–S108.
Bonnet D, Dick JE . Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat Med 3: 730–737.
Lapidot T, Sirard C, Vormoor J, Murdoch B, Hoang T, Caceres-Cortes J et al. A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature 1994; 367: 645–648.
Reinisch A, Chan SM, Thomas D, Majeti R . Biology and clinical relevance of acute myeloid leukemia stem cells. Semin Hematol 2015; 52: 150–164.
Litchfield DW . Protein kinase CK2: structure, regulation and role in cellular decisions of life and death. Biochem J 2003; 369 (Pt 1): 1–15.
Piazza F, Manni S, Ruzzene M, Pinna LA, Gurrieri C, Semenzato G . Protein kinase CK2 in hematologic malignancies: reliance on a pivotal cell survival regulator by oncogenic signaling pathways. Leukemia 2012; 26: 1174–1179.
Mandato E, Manni S, Zaffino F, Semenzato G, Piazza F . Targeting CK2-driven non-oncogene addiction in B-cell tumors. Oncogene 2016; e-pub ahead of print 4 April 2016 doi:10.1038/onc.2016.86.
Kato Jr T, Delhase M, Hoffmann A, Karin M . CK2 is a C-terminal IκB kinase responsible for NF-κB activation during the UV response. Mol Cell 2003; 12: 829–839.
Wang D, Westerheide SD, Hanson JL, Baldwin AS . Tumor necrosis factor α-induced phosphorylation of RelA/p65 on Ser529 is controlled by casein kinase II. J Biol Chem 2000; 275: 32592–32597.
Zheng Y, Qin H, Frank SJ, Deng L, Litchfield DW, Tefferi A et al. A CK2-dependent mechanism for activation of the JAK-STAT signaling pathway. Blood 2011; 118: 156–166.
Torres J, Pulido R . The tumor suppressor PTEN is phosphorylated by the protein kinase CK2 at its C terminus. Implications for PTEN stability to proteasome-mediated degradation. J Biol Chem 2001; 276: 993–998.
Di Maira G, Salvi M, Arrigoni G, Marin O, Sarno S, Brustolon F et al. Protein kinase CK2 phosphorylates and upregulates Akt/PKB. Cell Death Differ 2005; 12: 668–677.
Hong S-H, Yang S-J, Kim T-M, Shim J-S, Lee H-S, Lee G-Y et al. Molecular integration of HoxB4 and STAT3 for self-renewal of hematopoietic stem cells: a model of molecular convergence for stemness. Stem Cells 2014; 32: 1313–1322.
Chung Y-J, Park B-B, Kang Y-J, Kim T-m, Eaves CJ, Oh I-H . Unique effects of Stat3 on the early phase of hematopoietic stem cell regeneration. Blood 2006; 108: 1208–1215.
Stein SJ, Baldwin AS . Deletion of the NF-κB subunit p65/RelA in the hematopoietic compartment leads to defects in hematopoietic stem cell function. Blood 2013; 121: 5015–5024.
Martelli AM, Evangelisti C, Chiarini F, Grimaldi C, Cappellini A, Ognibene A et al. The emerging role of the phosphatidylinositol 3-kinase/Akt/mammalian target of rapamycin signaling network in normal myelopoiesis and leukemogenesis. Biochim Biophys Acta 2010; 1803: 991–1002.
Miyamoto K, Araki KY, Naka K, Arai F, Takubo K, Yamazaki S et al. Foxo3a is essential for maintenance of the hematopoietic stem cell pool. Cell Stem Cell 2007; 1: 101–112.
Tzivion G, Dobson M, Ramakrishnan G . FoxO transcription factors; regulation by AKT and 14-3-3 proteins. Biochim Biophys Acta 2011; 1813: 1938–1945.
Sykes SM, Lane SW, Bullinger L, Kalaitzidis D, Yusuf R, Saez B et al. AKT/FOXO signaling enforces reversible differentiation blockade in myeloid leukemias. Cell 2011; 146: 697–708.
Naka K, Hoshii T, Muraguchi T, Tadokoro Y, Ooshio T, Kondo Y et al. TGF-beta-FOXO signalling maintains leukaemia-initiating cells in chronic myeloid leukaemia. Nature 2010; 463: 676–680.
Kim JS, Eom JI, Cheong JW, Choi AJ, Lee JK, Yang WI et al. Protein kinase CK2alpha as an unfavorable prognostic marker and novel therapeutic target in acute myeloid leukemia. Clin Cancer Res 2007; 13: 1019–1028.
Quotti Tubi L, Gurrieri C, Brancalion A, Bonaldi L, Bertorelle R, Manni S et al. Inhibition of protein kinase CK2 with the clinical-grade small ATP-competitive compound CX-4945 or by RNA interference unveils its role in acute myeloid leukemia cell survival, p53-dependent apoptosis and daunorubicin-induced cytotoxicity. J Hematol Oncol 2013; 6: 78.
Cheong J-W, Min YH, Eom JI, Kim SJ, Jeung HK, Kim JS . Inhibition of CK2α and PI3K/Akt synergistically induces apoptosis of CD34+CD38− leukaemia cells while sparing haematopoietic stem cells. Anticancer Res 2010; 30: 4625–4634.
Chon HJ, Bae KJ, Lee Y, Kim J . The casein kinase 2 inhibitor, CX-4945, as an anti-cancer drug in treatment of human hematological malignancies. Front Pharmacol 2015; 6: 70.
Pierre F, Chua PC, O'Brien SE, Siddiqui-Jain A, Bourbon P, Haddach M et al. Pre-clinical characterization of CX-4945, a potent and selective small molecule inhibitor of CK2 for the treatment of cancer. Mol Cell Biochem 2011; 356: 37–43.
Pedranzini L, Mottadelli F, Ronzoni S, Rossella F, Ferracin M, Magnani I et al. Differential cytogenomics and miRNA signature of the acute myeloid leukaemia Kasumi-1 cell line CD34+38-compartment. Leuk Res 2010; 34: 1287–1295.
Yde CW, Olsen BB, Meek D, Watanabe N, Guerra B . The regulatory beta-subunit of protein kinase CK2 regulates cell-cycle progression at the onset of mitosis. Oncogene 2008; 27: 4986–4997.
Guerra B, Issinger OG . Protein kinase CK2 and its role in cellular proliferation, development and pathology. Electrophoresis 1999; 20: 391–408.
Li D, Dobrowolska G, Aicher LD, Chen M, Wright JH, Drueckes P et al. Expression of the casein kinase 2 subunits in Chinese hamster ovary and 3T3 L1 cells provides information on the role of the enzyme in cell proliferation and the cell cycle. J Biol Chem 1999; 274: 32988–32996.
Liu S, Dontu G, Mantle ID, Patel S, Ahn NS, Jackson KW et al. Hedgehog signaling and Bmi-1 regulate self-renewal of normal and malignant human mammary stem cells. Cancer Res 2006; 66: 6063–6071.
Dimri GP, Martinez JL, Jacobs JJ, Keblusek P, Itahana K, Van Lohuizen M et al. The Bmi-1 oncogene induces telomerase activity and immortalizes human mammary epithelial cells. Cancer Res 2002; 62: 4736–4745.
Essaghir A, Dif N, Marbehant CY, Coffer PJ, Demoulin J-B . The transcription of FOXO genes is stimulated by FOXO3 and repressed by growth factors. J Biol Chem 2009; 284: 10334–10342.
Liu CM, Chang CH, Yu CH, Hsu CC, Huang LL . Hyaluronan substratum induces multidrug resistance in human mesenchymal stem cells via CD44 signaling. Cell Tissue Res 2009; 336: 465–475.
Zoller M . CD44: can a cancer-initiating cell profit from an abundantly expressed molecule? Nat Rev Cancer 2011; 11: 254–267.
Jin L, Hope KJ, Zhai Q, Smadja-Joffe F, Dick JE . Targeting of CD44 eradicates human acute myeloid leukemic stem cells. Nat Med 2006; 12: 1167–1174.
Schust J, Sperl B, Hollis A, Mayer TU, Berg T . Stattic: a small-molecule inhibitor of STAT3 activation and dimerization. Chem Biol 2006; 13: 1235–1242.
Logue JS, Morrison DK . Complexity in the signaling network: insights from the use of targeted inhibitors in cancer therapy. Genes Dev 2012; 26: 641–650.
Jia H, Liu Y, Xia R, Tong C, Yue T, Jiang J et al. Casein kinase 2 promotes Hedgehog signaling by regulating both smoothened and Cubitus interruptus. J Biol Chem 2010; 285: 37218–37226.
Stahl M, Dijkers PF, Kops GJ, Lens SM, Coffer PJ, Burgering BM et al. The forkhead transcription factor FoxO regulates transcription of p27Kip1 and Bim in response to IL-2. J Immunol 2002; 168: 5024–5031.
Kornblau SM, Singh N, Qiu Y, Chen W, Zhang N, Coombes KR . Highly phosphorylated FOXO3A is an adverse prognostic factor in acute myeloid leukemia. Clin Cancer Res 2010; 16: 1865–1874.
Siddiqui-Jain A, Drygin D, Streiner N, Chua P, Pierre F, O'Brien SE et al. CX-4945, an orally bioavailable selective inhibitor of protein kinase CK2, inhibits prosurvival and angiogenic signaling and exhibits antitumor efficacy. Cancer Res 2010; 70: 10288–10298.
Zhang Y, Xing Y, Zhang L, Mei Y, Yamamoto K, Mak TW et al. Regulation of cell cycle progression by forkhead transcription factor FOXO3 through its binding partner DNA replication factor Cdt1. Proc Natl Acad Sci USA 2012; 109: 5717–5722.
Santini V, Nooter K, Delwel R, Lowenberg B . Susceptibility of acute myeloid leukemia (AML) cells from clinically resistant and sensitive patients to daunomycin (DNR): assessment in vitro after stimulation with colony stimulating factors (CSFs). Leuk Res 1990; 14: 377–380.
Acknowledgements
We thank patients and their families for donating samples. We thank Dr A Cabrelle (Venetian Institute of Molecular Medicine (VIMM)) for helping with FACS experiments. This work was supported by grants from Associazione Italiana per la Ricerca sul Cancro (AIRC; no. 14481), from the Italian Ministry of Education, University and Research (FIRB (Futuro in Ricerca) no. RBFR086EW9_001), from the University of Padova (Progetti di Ricerca di Ateneo no. CPDA114940/11) to FP and from AIRC IG to GS. SM is supported by a 2014 fellowship of the ‘Fondazione Umberto Veronesi’, Milan, Italy.
Author contributions
LQT and SCN performed research, analyzed data and wrote the manuscript. AB, EDB, SM, EM, FZ, PM, MC and KG helped in experiments and partly analyzed data; LT, GB, RZ and CG contributed patient samples and critical advices during elaboration of data; GS contributed patients and provided funding; FP conceived and designed the study, provided funding, contributed patient samples, supervised research and data analysis and wrote/edited the manuscript.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on the Leukemia website
Rights and permissions
About this article

Cite this article
Quotti Tubi, L., Canovas Nunes, S., Brancalion, A. et al. Protein kinase CK2 regulates AKT, NF-κB and STAT3 activation, stem cell viability and proliferation in acute myeloid leukemia. Leukemia 31, 292–300 (2017). https://doi.org/10.1038/leu.2016.209
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/leu.2016.209
- Springer Nature Limited
This article is cited by
-
Cancer stem cells: advances in knowledge and implications for cancer therapy
Signal Transduction and Targeted Therapy (2024)
-
Gene expression profiling unveils the temporal dynamics of CIGB-300-regulated transcriptome in AML cell lines
BMC Genomics (2023)
-
Low doses of IFN-γ maintain self-renewal of leukemia stem cells in acute myeloid leukemia
Oncogene (2023)
-
Protein kinase CK2 – diverse roles in cancer cell biology and therapeutic promise
Molecular and Cellular Biochemistry (2023)
-
Baicalein attenuates cardiac hypertrophy in mice via suppressing oxidative stress and activating autophagy in cardiomyocytes
Acta Pharmacologica Sinica (2021)