
Abstract
Phosphatase and tensin homolog (PTEN), a tumor suppressor gene, by negatively regulating the PI3K-Akt signaling pathway, participates in multiple biological processes such as cell proliferation, apoptosis, differentiation, and migration. Recent studies show that selective deletion of PTEN in pancreatic β-cells leads to resistance to streptozotocin (STZ)-induced diabetes, but the mechanism is unclear. One major mechanism underlying STZ toxicity is cytokine-mediated β-cell destruction in which oxidative stress plays a key role. The present study investigated the role of PTEN in cytokine-induced β-cell apoptosis, and further explored whether oxidative stress, particularly peroxynitrite formation, could regulate PTEN-Akt pathway. Incubation of βTC-6 cells with cytokine mixture (IL-1β, TNF-α, and IFN-γ) or exogenous peroxynitrite significantly increased apoptotic cell percentage, elevated PTEN and p-PTEN levels, and inhibited Akt activation. Transfection with PTEN-specific siRNA protected βTC-6 cells from cytokine or peroxynitrite-mediated cell apoptosis and partially reversed Akt inhibition. Furthermore, nitrotyrosine formation, an indicator of peroxynitrite production, was significantly elevated after cytokine treatment. Preventing peroxynitrite formation by administrating NAC/l-NMMA, or scavenging peroxynitrite directly by UA, attenuated cytokine-induced PTEN upregulation, Akt inhibition, and β-cell apoptosis. These findings suggest that peroxynitrite-mediated PTEN upregulation plays an important role in cytokine-induced pancreatic β-cell apoptosis.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Phosphatase and tensin homolog (PTEN), originally identified as a tumor suppressor gene frequently mutated in a large percentage of human cancers, is the central negative regulator of the PI3K-Akt signal transduction cascade [1]. Reduced PTEN function results in marked activation of Akt survival pathways, leading to hyperplasia, inhibition of apoptosis, and thereby contributing to tumor formation [2]. Recent studies have given insight to the broader role of PTEN beyond tumor suppression. In major insulin target organs (principally the liver, muscle, and fat), PTEN deficiency affects glucose metabolism and protects mice from developing diabetes [3–5]. Importantly, pancreas-specific PTEN deletion leads to resistance to streptozotocin (STZ)-induced diabetes and decreased β-cell apoptosis [6, 7], but the underlying mechanisms are still unclear. As a major mechanism of STZ toxicity is believed to be cytokine-mediated [8, 9], and evidence supports that cytokines cause β-cell loss by inducing apoptosis [10–12], we hypothesize that PTEN may play a role in cytokine-induced β-cell apoptosis.
It has been reported that generation of high levels of reactive oxygen/nitrogen species plays a key role in cytokine-induced β-cell apoptosis [13, 14]. Peroxynitrite, the reaction product of nitric oxide (NO) and superoxide, is a highly reactive radical species with increased toxic potential compared with NO or superoxide alone [15]. Scavenging peroxynitrite has been shown to prevent diabetes development in NOD mice, and significantly decreases β-cell destruction when incubated with cytokines [16]. Expression of calbindin-D28k can protect βTC-3 cells against cytokine-mediated apoptosis by inhibiting peroxynitrite formation [17]. These findings suggest that peroxynitrite is a key mediator of β-cell apoptosis induced by cytokines.
Furthermore, several studies have demonstrated that oxidative stress is an important regulator of PTEN expression/activation [18, 19]. It has been specifically reported that peroxynitrite inhibits PTEN, activating the anti-apoptotic PI3K-Akt pathway in primary neurons, and consequently providing neuroprotection [20]. Studies using human umbilical vein endothelial cells have shown that hyperglycemia triggers apoptosis by inhibiting Akt signaling via peroxynitrite-mediated PTEN activation [21]. We currently investigate the role of PTEN in cytokine-induced apoptosis, and explore the potential regulatory effect of peroxynitrite upon the PTEN-Akt pathway in β-cells.
Materials and methods
Reagents
N-Acetyl-l-cysteine (NAC), NG-Methyl-l-arginine acetate salt (l-NMMA), Uric acid (UA), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) and Hoechst 33342 were purchased from Sigma-Aldrich (St. Louis, MO). Peroxynitrite was obtained from Calbiochem (San Diego, CA). Dulbecco’s modified Eagle’s medium (DMEM) and fetal calf serum (FCS) were purchased from Invitrogen (Carlsbad, CA). Murine IL-1β, murine TNFα, and murine IFNγ were purchased from Peprotech (Rocky Hill, NJ). Antibodies against Akt, phospho-Akt-Ser473, PTEN, phospho-PTEN-Ser380/Thr382/383, and caspase-3; PTEN small interference RNA (siRNA) and control siRNA were all from Cell Signaling Technology (Beverly, MA). siRNA delivery agent DharmaFECT 2 was obtained from Dharmacon (Lafayette, CO). Nitrotyrosine Assay Kit (Chemiluminescence Detection) was from Upstate (Temecula, CA). β-actin antibody and HRP-conjugated secondary antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA) and CaspaseTM assay system (colorimetric) was purchased from Promega (Madison, WI).
Cell culture and treatment
Mouse βTC-6 cells were grown in DMEM medium supplemented with 10% heat-inactivated FCS, penicillin (100 IU/ml), streptomycin (100 μg/ml), and 2 mM l-glutamine in the presence of 5% CO2/95% air with 100% relative humidified atmosphere at 37°C. Medium was changed every 2–3 days. Cells were passaged at 80% confluence. Unless otherwise stated, cells in logarithmic growth phase were treated with cytokine mixture for 24 h. The following cytokine concentrations were used based on dose–response experiments (data not shown): recombinant murine IL-1β at 2 ng/ml, recombinant murine TNF-α at 5 ng/ml, and recombinant murine IFN-γ at 25 ng/ml. Cells were preincubated for 1 h with pharmacologic inhibitors NAC (1 mM), l-NMMA (1 mM), or UA (0.5 mM) prior to cytokine exposure. During time course analysis of protein levels, cells were treated at time zero, and harvested at the indicated time points.
Cells were treated with peroxynitrite as described previously [22]. After PBS wash, and equilibration (pH 7.4) for 5 min, cells were exposed to 5 μM peroxynitrite (diluted in 10 mM NaOH) in PBS for 1 h. Subsequently, PBS was removed and replaced by fresh serum-free medium and incubated for another 6 h.
Cell viability assay
The viability of βTC-6 cells was determined by a colorimetric MTT assay. Cells were cultured in 96-well plates at a density of 2 × 105/ml. After designed treatment, 20 μl MTT (5 mg/ml) was added and cells were incubated for 4 more hours. Medium was removed, and the formazan crystals were dissolved in 150 μl DMSO with shaking for 10 min. The absorbance at 570 nm was read using a microplate reader (Bio-Rad, USA). Cell viability was expressed as a percentage to untreated control.
Hoechst 33342 staining
Morphologic evidence of apoptosis was assessed by nuclear staining with Hoechst 33342. After above-mentioned treatment, βTC-6 cells, seeded on coverslides in 24-well plates, were fixed with 4% paraformaldehyde for 30 min, washed with PBS, and stained with Hoechst 33342 (0.5 μg/ml) for 10 min at room temperature in the dark. Samples were observed under a fluorescence microscope (Olympus, Japan). Cells intensely fluorescing and/or with condensed nuclei were considered apoptotic. Apoptotic and total cells were counted in four random fields of four different slides. The number of apoptotic cells was expressed as a percentage to total cells.
Measurement of caspase-3 activity
Caspase-3 activity was determined using a colorimetric caspase-3 assay kit (CaspACETM Assay System, Colorimetric) per manufacturer’s protocol. Briefly, treated cells were lysed in lysis buffer and centrifuged. The supernatant was collected and protein concentrations were determined by the Bradford method. Samples were mixed with Ac-DEVD-pNA, a caspase-3 substrate, and incubated for 4 h at 37°C. Enzyme-catalyzed release of pNA was monitored at 405 nm by means of a microplate reader (Bio-Rad, USA), and the caspase-3 activity was calculated per manufacturer’s instruction.
siRNA gene silencing of PTEN
Mouse βTC-6 cells were seeded in 6-well plates and incubated overnight. PTEN siRNA or control siRNA was mixed with DharmaFECT 2 transfection reagent per manufacturer’s instructions. siRNA-DharmaFECT complexes were added to the cells, which had been washed twice with serum free medium. After 6 h incubation, the transfection medium was replaced by full culture medium. The control siRNA contained inert genetic sequences non-degrading of cellular material.
Nitrotyrosine assay
After treatment, cells were washed with cold PBS, lysed on ice, and centrifuged for 15 min at 12,000 g at 4°C. The supernatant was collected and protein concentrations were determined by the bicinchoninic acid method. Nitrotyrosine levels were determined via a modified ELISA procedure [23, 24]. In short, after blocking the nonspecific binding sites, a nitrated protein solution was prepared and diluted for use as a standard. These standard samples, along with cell samples (50 μl/well), were applied to the ELISA plate and allowed to incubated for 1 h with a rabbit polyclonal anti-nitrotyrosine primary antibody (1:100) at 37°C. Samples were then incubated with a HRP-conjugated secondary antibody (1:5,000) for 1 h. After washing, samples were incubated with freshly prepared LumiGLO® Chemiluminescent Substrate (75 μl/well) for 10 min. Luminescence was measured with a microplate reader (Bio-Rad, USA). The nitrotyrosine content in cell samples was calculated using standard curves generated from nitrated BSA containing quantified nitrotyrosine amounts.
Western blotting
After treatment with cytokine combination for 4 h, cells were washed with cold PBS and lysed in lysis buffer (20 mM Tris–HCl of pH 7.5, 150 mM NaCl, 1 mM Na2EDTA, 1 mM EGTA, 1% Triton, 2.5 mM sodium pyrophosphate, 1 mM beta-glycerophosphate, 1 mM Na3VO4, and 1 μg/ml leupeptin) with PMSF on ice. Proteins (~35 μg) from each sample were separated by electrophoresis on a SDS-PAGE gel and transferred onto a nitrocellulose membrane using semi-dry blotting apparatus (Bio-Rad, USA). The membrane was blocked for 1 h with 5% non-fat milk in TBST (Tris Buffered Saline-0.1% Tween 20), and then probed with primary antibodies (PTEN, 1:1000; p-PTEN, 1:1000; Akt, 1:1000; p-Akt, 1:1000; caspase-3, 1:1000; β-actin, 1:1000) overnight at 4°C. After incubation with HRP-conjugated secondary antibodies (1:5,000), specific bands were visualized with the ECL detection system.
To detect reduced and oxidized forms of PTEN, protein samples were fractionated by SDS-PAGE under non-reducing conditions, and subjected to PTEN immunoblot assay.
Statistical analysis
All experiments were repeated at least three times. All values in the text and figures are presented as mean ± SEM. All data were subjected to one-way ANOVA followed by Bonferroni correction for post hoc t test. P values less than 0.05 were considered statistically significant.
Results
IL-1β, TNF-α, and IFN-γ induce apoptosis of βTC-6 cells
Exposure to the cytokine mixture of IL-1β (2 ng/ml), TNFα (5 ng/ml), and IFNγ (25 ng/ml) for 24 h significantly decreased viability of mouse pancreatic βTC-6 cells, as determined by MTT assay (Fig. 1a). To observe the cytokine induced-apoptosis of βTC-6 cells, we performed Hoechst staining (Fig. 1b, c) and measured caspase-3 activity (Fig. 1c, d). The round nuclei of normal cells demonstrated homogeneous and diffused staining with regular contours. After cytokine treatment, there was significantly increased apoptotic cells, which exhibited an asymmetric and bright blue fluorescence. Caspase-3 activation was used as a key marker of apoptosis. Elevated levels of cleaved caspase-3 and caspase-3 activity were observed after cytokine exposure. Each cytokine alone had no notable effect on cell apoptosis at the dose and time course regiment investigated.

Cytokine mixture induced apoptosis of βTC-6 cells. Cells were treated with the combination of IL-1β (2 ng/ml), TNF-α (5 ng/ml), and IFN-γ (25 ng/ml) or each alone for 24 h. a Cell viability was measured using MTT assay and expressed as a percentage to untreated control (n = 5). b Morphology of untreated or cytokine-treated βTC-6 cells was visualized by Hoechst 33342 staining (Bar = 50 μm). c The apoptotic cells were assessed by Hoechst 33342 staining (n = 4) and caspase-3 activity (n = 3). d Caspase-3 expression was detected by Western blotting. Results are shown as mean ± SEM. Con indicates control; and CTK cytokine mixture. * P < 0.05 vs. control
Cytokines induce the upregulation of PTEN and p-PTEN
To investigate whether PTEN was targeted by cytokines, we examined the protein levels of total PTEN and phosphorylated PTEN (S380/T382/383). As shown in Fig. 2a, a time course analysis revealed that the level of total PTEN increased approximately 2.4-fold after 4 h cytokine exposure. Similarly, cytokines up-regulated p-PTEN levels, peaking at 1 h (~2.0-fold vs. control, P < 0.05). A slight decline of p-PTEN levels was observed at 4 h (~1.6-fold vs. control, P < 0.05), plateauing for 24 h. The time-matched controls (0, 4 h) had no significant difference in protein levels.

a Cytokines increased the protein levels of PTEN and p-PTEN. Cells were treated with or without the combination of IL-1β (2 ng/ml), TNF-α (5 ng/ml), and IFN-γ (25 ng/ml) for the indicated times. b PTEN siRNA decreased basal expression of PTEN/p-PTEN and inhibited cytokine-induced elevation of PTEN/p-PTEN. Cells were transfected with either control siRNA (100 nM) or PTEN siRNA (100 nM) for 48 h and then treated with or without the combination of cytokines for 4 h. Results are shown as mean ± SEM. Con indicates control; CTK cytokine mixture; s-Con control siRNA; and s-PTEN PTEN siRNA. * P < 0.05 vs. control. # P < 0.05 s-PTEN/CTK vs. s-Con/CTK
Knockdown of PTEN decreases cell apoptosis induced by cytokines
To determine whether PTEN upregulation was responsible for cytokine-induced apoptosis, βTC-6 cells were transfected with PTEN siRNA or control siRNA, and apoptosis initiation was monitored. Transfection efficiency was estimated between 70 and 80%. PTEN siRNA suppressed the basal expression of PTEN and p-PTEN. In parallel, PTEN siRNA attenuated the elevation of PTEN and p-PTEN induced by cytokines (Fig. 2b). Importantly, βTC-6 cells transfected with PTEN siRNA, but not control siRNA, were resistant to cytokine toxicity. Cells transfected with control siRNA had significantly decreased viability after cytokine treatment, similar to non-transfected cells. However, the viability of cells transfected with PTEN siRNA was increased compared to control after cytokine treatment (Fig. 3a). Transfection with PTEN siRNA resulted in significantly reduced pro-apoptotic effects of cytokines (Fig. 3b, c).

PTEN siRNA decreased cytokine-induced apoptosis in βTC-6 cells. Cells were transfected with either control siRNA (100 nM) or PTEN siRNA (100 nM) for 24 h and treated with or without the combination of cytokines for another 24 h. a Cell viability was measured using MTT assay and expressed as a percentage to untreated control (n = 5). b Caspase-3 expression was detected by Western blotting. c The apoptotic cells were assessed by Hoechst 33342 staining (n = 4) and caspase-3 activity (n = 3). Results are shown as mean ± SEM. Con indicates control; CTK cytokine mixture; S-Con control siRNA; and S-PTEN PTEN siRNA. * P < 0.05 vs. Con. # P < 0.05 s-PTEN/CTK vs. s-Con/CTK
Knockdown of PTEN reverses cytokine-induced Akt inhibition
Akt is known to play a key role in β-cell growth and survival, and we investigated the effects of cytokines on total Akt and Akt-Ser473 phosphorylation in cultured βTC-6 cells. As shown in Fig. 4a, exposure of βTC-6 cells to cytokines for 1 h led to significant inhibition of Akt-Ser473 phosphorylation, a result even more prominent after 4 h of treatment. Total Akt levels did not change during cytokine stimulation.

a Cytokines decreased Akt phosphorylation in βTC-6 cells. Cells were treated with or without the combination of IL-1β (2 ng/ml), TNF-α (5 ng/ml), and IFN-γ (25 ng/ml) for the indicated times. b PTEN siRNA attenuated cytokine-induced Akt inhibition. Cells were transfected with either control siRNA (100 nM) or PTEN siRNA (100 nM) for 48 h, and then treated with or without the combination of cytokines for 4 h. Results are shown as mean ± SEM. Con indicates control; CTK cytokine mixture; s-Con control siRNA; and s-PTEN PTEN siRNA. * P < 0.05 vs. Con. # P < 0.05 s-PTEN/CTK vs. s-Con/CTK
It has been well established that PTEN is a lipid phosphatase specifically dephosphorylating plasma membrane lipid PIP3. Conversion of PIP3 to PIP2 enables PTEN to functionally antagonize PI3K, thereby inactivating downstream protein kinases, most notably Akt [25]. Conversely, PTEN inhibition, or mutation, impairs PIP3 inactivation, thus enhancing the Akt-mediated anti-apoptotic pathway [20]. We assessed the levels of phosphorylated Akt in βTC-6 cells transfected with or without PTEN siRNA, to determine whether PTEN elevation was responsible for the cytokine-induced Akt inhibition. As shown in Fig. 4b, siRNA-induced reduction of PTEN reversed cytokine-induced Akt inhibition.
Cytokines cause the upregulation of PTEN via peroxynitrite
Several studies have shown that peroxynitrite is a key mediator of cytokine-induced β-cell destruction, but it is still unknown whether increased peroxynitrite formation will contribute to the upregulation of PTEN induced by cytokines. To detect the role of peroxynitrite in the signaling pathway, βTC-6 cells transfected with or without PTEN siRNA were directly treated with exogenous peroxynitrite (5 μM) for 1 h. Treatment with peroxynitrite induced apoptosis of βTC-6 cells and, similar to cytokines, increased the levels of PTEN and p-PTEN, and inhibited Akt activation. Moreover, cells transfected with PTEN siRNA partially reversed peroxynitrite-induced cell apoptosis and Akt suppression (Fig. 5).

Knockdown of PTEN partially reversed peroxynitrite-induced cell apoptosis and Akt inhibition. a The apoptotic cells were assessed by Hoechst 33342 staining (n = 4) and caspase-3 activity (n = 3). b Caspase-3 expression was detected by Western blotting. c The protein levels of PTEN/p-PTEN and Akt/p-Akt were detected by Western blotting. Cells were transfected with either control siRNA (100 nM) or PTEN siRNA (100 nM) for 48 h and treated with or without peroxynitrite (5 μM) for 1 h. Subsequently, PBS was replaced by serum-free medium and incubated for another 6 h. Results are shown as mean ± SEM. Con indicates control; PN peroxynitrite; s-Con control siRNA; and s-PTEN PTEN siRNA. * P < 0.05 vs. Con. # P < 0.05 s-PTEN/PN vs. s-Con/PN
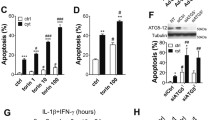
Next, we measured nitrotyrosine production in βTC-6 cells with or without cytokine exposure, a footprint of increased peroxynitrite presence. Consistent with previous studies in pancreatic islets and in other β-cell lines [17, 26, 27], the mixture of cytokines significantly increased the formation of nitrotyrosine (Fig. 6a). While it is known that PTEN activity can be regulated by oxidation, cytokine treatment in our study for a duration of 4 or 24 h did not result in PTEN oxidation (Fig. 6b). To further confirm the role of peroxynitrite in cytokine-induced PTEN upregulation, pharmacologic inhibitors, including NAC (an O •−2 scavenger), l-NMMA (a NOS inhibitor), or UA (a peroxynitrite scavenger), were applied prior to cytokine administration. Both inhibition of peroxynitrite formation by administrating NAC or l-NMMA and scavenging peroxynitrite directly by UA attenuated cytokine-induced nitrotyrosine production (Fig. 6a) and apoptosis (Fig. 7). Simultaneously, these inhibitors significantly decreased the protein levels of PTEN and p-PTEN, and increased Akt phosphorylation compared to cells exposed to cytokines alone (Fig. 8). In addition, NAC, l-NMMA, and UA alone had no effect on baseline cell viability and PTEN level (data not shown). Taken together, these data suggested that peroxynitrite-mediated PTEN upregulation played a potential role in the cytokine-induced βTC-6 cell apoptosis.

a NAC, l-NMMA, or UA decreased cytokine-induced nitrotyrosine formation. Exposure of βTC-6 cells with cytokines for 24 h with or without the preincubation with NAC (1 mM), l-NMMA (1 mM), or UA (0.5 mM) for 1 h. Nitrotyrosine formation was detected by ELISA (n = 3). Results are shown as mean ± SEM. Con indicates control; CTK cytokine mixture; and UA uric acid. * P < 0.05 vs. Con. # P < 0.05 vs. CTK. b Cytokines did not cause PTEN oxidation in βTC-6 cells. The reduced and oxidized forms of PTEN were fractionated by nonreducing SDS-PAGE and identified by immunoblot assay. Cells were treated with H2O2 (500 μM) for 5 min as a control for oxidized PTEN. 2-mercaptoethanol (2-ME) was added to an aliquot of cell lysate as a control for reduced PTEN

NAC, l-NMMA, or UA decreased cytokine-induced βTC-6 cell apoptosis. Exposure of βTC-6 cells with cytokines for 24 h with or without the preincubation with NAC (1 mM), L-NMMA (1 mM), or UA (0.5 mM) for 1 h. a Cell viability was measured using MTT assay and expressed as a percentage to untreated control (n = 5). b The apoptotic cells were assessed by Hoechst 33342 staining (n = 4) and caspase-3 activity (n = 3). c Caspase-3 expression was detected by Western blotting. Results are shown as mean ± SEM. Con indicates control; CTK cytokine mixture; and UA uric acid. * P < 0.05 vs. Con. # P < 0.05 vs. CTK

NAC, l-NMMA, or UA attenuated cytokine-induced PTEN/p-PTEN elevation and Akt inhibition. Exposure of βTC-6 cells with cytokines for 4 h with or without the preincubation with NAC (1 mM), l-NMMA (1 mM), or UA (0.5 mM) for 1 h. Results are shown as mean ± SEM. Con indicates control; CTK cytokine mixture; and UA uric acid. * P < 0.05 vs. Con. # P < 0.05 vs. CTK
Discussion
In type 1 diabetes, insulin-producing β-cells are selectively destroyed by the cellular autoimmune response. Proinflammatory cytokines (such as IL-1β, TNF-α, and IFN-γ) released during this autoimmune response are regarded as important mediators of β-cell apoptosis [28]. Recently, inflammatory mediators have been increasingly implicated in type 2 diabetes development [29–31]. These data strongly suggest that cytokine signaling blockade may be a potential method for preventing diabetic β-cell loss. The present study is the first to demonstrate that peroxynitrite-mediated the upregulation of PTEN and p-PTEN via inhibition of Akt activation, plays an important role in cytokine induced β-cell apoptosis.
In our experiment, the combination of IL-1β, TNF-α, and IFN-γ significantly decreased β-cells viability, consistent with previous studies [11, 32]. Additionally, because cytokines likely act synergistically during in vivo pancreatic immune infiltration inducing β-cell damage, we tailored our study to focus on the effect of these three typical inflammatory cytokines in combination. To investigate the mechanisms responsible for cytokine-induced β-cell injury, we evaluated the morphologic and biochemical apoptotic hallmarks by Hoechst staining and caspase-3 activity assays. The results confirmed that apoptosis was the primary mechanism of cell death by cytokine exposure. At the concentration- and time- dose utilized in our study, each cytokine alone demonstrated no detrimental effect on cellular viability. While some studies have demonstrated that IL-1β alone was sufficient to cause β-cell death or the impairment of cell function [33, 34], others report that massive induction of β-cells apoptosis usually requires a combination of IL-1β plus TNF-α and/or IFN-γ [10, 11, 35].
The PI3K/Akt pathway is critically involved in regulating cell survival [36, 37]. It has been previously demonstrated that cytokine-induced apoptosis was significantly increased in β-cells infected with kinase-dead Akt [38], while cytokine-induced decrease of bcl-2 promoter activity and increase of β-cell apoptosis were partially restored when cells were cotransfected with a constitutively active form of Akt [28]. PTEN, a member of the protein tyrosine phosphatase family, by negatively modulating the PI3K-Akt signaling pathway, participates in multiple biological processes such as cell proliferation, differentiation, apoptosis, and migration [39–41]. In the present study, exposure of βTC-6 cells to cytokine mixture significantly elevated PTEN level, inhibited Akt-Ser473 phosphorylation, and increased the percentage of apoptotic cells. Moreover, knockdown of PTEN with PTEN-specific siRNA partially reversed cytokine-induced Akt inhibition and cell apoptosis. These results imply that PTEN is involved in the signaling pathway of cytokine-induced β-cell apoptosis. More importantly, the cell apoptosis and Akt inactivation are PTEN-dependent.
The activity and half-life of PTEN are reciprocally regulated by phosphorylation. Several studies have demonstrated that, after phosphorylation, reduced PTEN activity can facilitate cellular protection via activators of the PI3K/Akt prosurvival kinase pathway [42]. Other studies have indicated that phosphorylation of PTEN increases its stability, while dephosphorylation leads to its rapid degradation [43, 44]. Our experimental data showed that PTEN was phosphorylated under basal conditions. Moreover, the level of p-PTEN was significantly increased during cytokine-induced apoptosis, which was accompanied by the PTEN level elevation. These findings are consistent with the idea that phosphorylation of PTEN may increase the protein’s stability. As total PTEN level remained elevated for 4 h with slight decline of PTEN phosphorylation, PTEN expression may be also regulated by other mechanisms (such as transcriptional regulation and protein–protein interactions) requiring further investigation for elucidation.
The production of reactive oxygen/nitrogen species is a key regulator of PTEN function [18, 19, 21]. It has been reported that peroxynitrite (100 μM) could rapidly (5 or 15 min) oxidize PTEN along with Akt phosphorylation [20]. But, in our experiment, cytokine exposure did not cause PTEN oxidation. A possible explanation for the inconsistent result may be that peroxynitrite production post cytokine treatment was lower than the amount requisite for PTEN oxidation. Furthermore, the stimulatory effect of endogenous peroxynitrite may differ from exogenous sources.
While it has been established that peroxynitrite is a primary mediator of cytokine toxicity upon β-cell viability [16, 17, 26], whether cytokine-induced PTEN upregulation is caused by peroxynitrite formation remains unclear. In the present study, exogenous peroxynitrite treatment increased both PTEN and p-PTEN levels, and simultaneously inhibited Akt activation. Moreover, cells transfected with PTEN siRNA exhibited attenuated peroxynitrite-induced cell apoptosis and Akt inhibition compared to control. Assays for nitrotyrosine formation and pharmacologic inhibitors further confirmed peroxynitrite mediated PTEN-dependent Akt inhibition. We demonstrated that the elevated nitrotyrosine formation, up-regulated levels of PTEN and p-PTEN, Akt activation inhibition, and increased β-cell apoptosis induced by cytokines were all partially reversed by either peroxynitrite formation inhibition (NAC and l-NMMA) or direct scavenging peroxynitrite (UA). Taken together, our data demonstrated that cytokines induce PTEN-dependent Akt inhibition via peroxynitrite in βTC-6 cells apoptosis.
In conclusion, PTEN-dependent Akt inhibition is involved in the apoptosis induced by cytokine mixture in βTC-6 cells. PTEN-specific siRNA can partially attenuate cytokine toxicity. These findings suggest that PTEN may be a potential target for prevention of cytokine induced β-cell loss in diabetes. Moreover, our results provide a link between oxidative stress and β-cell destruction, as PTEN upregulation was mediated by peroxynitrite, a relationship that still needs in vivo clarification.
References
Baker SJ (2007) PTEN enters the nuclear age. Cell 128:25–28
Tell G, Pines A, Arturi F et al (2004) Control of phosphatase and tensin homolog (PTEN) gene expression in normal and neoplastic thyroid cells. Endocrinology 145:4660–4666
Wijesekara N, Konrad D, Eweida M et al (2005) Muscle-specific Pten deletion protects against insulin resistance and diabetes. Mol Cell Biol 25:1135–1145
Stiles B, Wang Y, Stahl A et al (2004) Liver-specific deletion of negative regulator Pten results in fatty liver and insulin hypersensitivity [corrected]. Proc Natl Acad Sci USA 101:2082–2087
Kurlawalla-Martinez C, Stiles B, Wang Y, Devaskar SU, Kahn BB, Wu H (2005) Insulin hypersensitivity and resistance to streptozotocin-induced diabetes in mice lacking PTEN in adipose tissue. Mol Cell Biol 25:2498–2510
Tong Z, Fan Y, Zhang W et al (2009) Pancreas-specific Pten deficiency causes partial resistance to diabetes and elevated hepatic AKT signaling. Cell Res 19:710–719
Stiles BL, Kuralwalla-Martinez C, Guo W et al (2006) Selective deletion of Pten in pancreatic beta cells leads to increased islet mass and resistance to STZ-induced diabetes. Mol Cell Biol 26:2772–2781
Amirshahrokhi K, Dehpour AR, Hadjati J, Sotoudeh M, Ghazi-Khansari M (2008) Methadone ameliorates multiple-low-dose streptozotocin-induced type 1 diabetes in mice. Toxicol Appl Pharmacol 232:119–124
Fukuda K, Tesch GH, Nikolic-Paterson DJ (2008) c-Jun amino terminal kinase 1 deficient mice are protected from streptozotocin-induced islet injury. Biochem Biophys Res Commun 366:710–716
Holohan C, Szegezdi E, Ritter T, O’Brien T, Samali A (2008) Cytokine-induced beta-cell apoptosis is NO-dependent, mitochondria-mediated and inhibited by BCL-XL. J Cell Mol Med 12:591–606
Abdelli S, Puyal J, Bielmann C et al (2009) JNK3 is abundant in insulin-secreting cells and protects against cytokine-induced apoptosis. Diabetologia 52:1871–1880
Ortis F, Pirot P, Naamane N et al (2008) Induction of nuclear factor-kappa B and its downstream genes by TNF-alpha and IL-1 beta has a pro-apoptotic role in pancreatic beta cells. Diabetologia 51:1213–1225
Chambers KT, Unverferth JA, Weber SM, Wek RC, Urano F, Corbett JA (2008) The role of nitric oxide and the unfolded protein response in cytokine-induced beta-cell death. Diabetes 57:124–132
Barbu A, Welsh N, Saldeen J (2002) Cytokine-induced apoptosis and necrosis are preceded by disruption of the mitochondrial membrane potential (Deltapsi(m)) in pancreatic RINm5F cells: prevention by Bcl-2. Mol Cell Endocrinol 190:75–82
Suarez-Pinzon WL, Szabo C, Rabinovitch A (1997) Development of autoimmune diabetes in NOD mice is associated with the formation of peroxynitrite in pancreatic islet beta-cells. Diabetes 46:907–911
Suarez-Pinzon WL, Mabley JG, Strynadka K, Power RF, Szabo C, Rabinovitch A (2001) An inhibitor of inducible nitric oxide synthase and scavenger of peroxynitrite prevents diabetes development in NOD mice. J Autoimmun 16:449–455
Rabinovitch A, Suarez-Pinzon WL, Sooy K, Strynadka K, Christakos S (2001) Expression of calbindin-D (28 k) in a pancreatic islet beta-cell line protects against cytokine-induced apoptosis and necrosis. Endocrinology 142:3649–3655
Lee SR, Yang KS, Kwon J, Lee C, Jeong W, Rhee SG (2002) Reversible inactivation of the tumor suppressor PTEN by H2O2. J Biol Chem 277:20336–20342
Fantini D, Vascotto C, Deganuto M et al (2008) APE1/Ref-1 regulates PTEN expression mediated by Egr-1. Free Radic Res 42:20–29
Delgado-Esteban M, Martin-Zanca D, Andres-Martin L, Almeida A, Bolanos JP (2007) Inhibition of PTEN by peroxynitrite activates the phosphoinositide-3-kinase/Akt neuroprotective signaling pathway. J Neurochem 102:194–205
Song P, Wu Y, Xu J et al (2007) Reactive nitrogen species induced by hyperglycemia suppresses Akt signaling and triggers apoptosis by upregulating phosphatase PTEN (phosphatase and tensin homologue deleted on chromosome 10) in an LKB1-dependent manner. Circulation 116:1585–1595
Brito PM, Simoes NF, Almeida LM, Dinis TC (2008) Resveratrol disrupts peroxynitrite-triggered mitochondrial apoptotic pathway: a role for Bcl-2. Apoptosis 13:1043–1053
Tanaka S, Choe N, Hemenway DR, Zhu S, Matalon S, Kagan E (1998) Asbestos inhalation induces reactive nitrogen species and nitrotyrosine formation in the lungs and pleura of the rat. J Clin Investig 102:445–454
Ma XL, Gao F, Nelson AH et al (2001) Oxidative inactivation of nitric oxide and endothelial dysfunction in stroke-prone spontaneous hypertensive rats. J Pharmacol Exp Ther 298:879–885
Zhu Y, Hoell P, Ahlemeyer B, Krieglstein J (2006) PTEN: a crucial mediator of mitochondria-dependent apoptosis. Apoptosis 11:197–207
Mathews CE, Suarez-Pinzon WL, Baust JJ, Strynadka K, Leiter EH, Rabinovitch A (2005) Mechanisms underlying resistance of pancreatic islets from ALR/Lt mice to cytokine-induced destruction. J Immunol 175:1248–1256
Lakey JR, Suarez-Pinzon WL, Strynadka K et al (2001) Peroxynitrite is a mediator of cytokine-induced destruction of human pancreatic islet beta cells. Lab Investig 81:1683–1692
Jambal P, Masterson S, Nesterova A et al (2003) Cytokine-mediated down-regulation of the transcription factor cAMP-response element-binding protein in pancreatic beta-cells. J Biol Chem 278:23055–23065
Malozowski S, Sahlroot JT (2007) Interleukin-1-receptor antagonist in type 2 diabetes mellitus. N Engl J Med 357:302–303 (author reply 303)
Donath MY, Schumann DM, Faulenbach M, Ellingsgaard H, Perren A, Ehses JA (2008) Islet inflammation in type 2 diabetes: from metabolic stress to therapy. Diabetes Care 31(Suppl 2):S161–S164
Ryan A, Murphy M, Godson C, Hickey FB (2009) Diabetes mellitus and apoptosis: inflammatory cells. Apoptosis 14:1435–1450
Gurgul E, Lortz S, Tiedge M, Jorns A, Lenzen S (2004) Mitochondrial catalase overexpression protects insulin-producing cells against toxicity of reactive oxygen species and proinflammatory cytokines. Diabetes 53:2271–2280
Larsen CM, Dossing MG, Papa S, Franzoso G, Billestrup N, Mandrup-Poulsen T (2006) Growth arrest- and DNA-damage-inducible 45 beta gene inhibits c-Jun N-terminal kinase and extracellular signal-regulated kinase and decreases IL-1 beta-induced apoptosis in insulin-producing INS-1E cells. Diabetologia 49:980–989
Hammar E, Parnaud G, Bosco D et al (2004) Extracellular matrix protects pancreatic beta-cells against apoptosis: role of short- and long-term signaling pathways. Diabetes 53:2034–2041
Azevedo-Martins AK, Lortz S, Lenzen S, Curi R, Eizirik DL, Tiedge M (2003) Improvement of the mitochondrial antioxidant defense status prevents cytokine-induced nuclear factor-kappa B activation in insulin-producing cells. Diabetes 52:93–101
Lee SC, Pervaiz S (2007) Apoptosis in the pathophysiology of diabetes mellitus. Int J Biochem Cell Biol 39:497–504
Flacke JP, Kumar S, Kostin S, Reusch HP, Ladilov Y (2009) Acidic preconditioning protects endothelial cells against apoptosis through p38- and Akt-dependent Bcl-xL overexpression. Apoptosis 14:90–96
Li L, El-Kholy W, Rhodes CJ, Brubaker PL (2005) Glucagon-like peptide-1 protects beta cells from cytokine-induced apoptosis and necrosis: role of protein kinase B. Diabetologia 48:1339–1349
Tamguney T, Stokoe D (2007) New insights into PTEN. J Cell Sci 120:4071–4079
Sugatani T, Alvarez U, Hruska KA (2003) PTEN regulates RANKL- and osteopontin-stimulated signal transduction during osteoclast differentiation and cell motility. J Biol Chem 278:5001–5008
Stambolic V, Suzuki A, de la Pompa JL et al (1998) Negative regulation of PKB/Akt-dependent cell survival by the tumor suppressor PTEN. Cell 95:29–39
Das S, Dixon JE, Cho W (2003) Membrane-binding and activation mechanism of PTEN. Proc Natl Acad Sci USA 100:7491–7496
Cai Z, Semenza GL (2005) PTEN activity is modulated during ischemia and reperfusion: involvement in the induction and decay of preconditioning. Circ Res 97:1351–1359
Vazquez F, Ramaswamy S, Nakamura N, Sellers WR (2000) Phosphorylation of the PTEN tail regulates protein stability and function. Mol Cell Biol 20:5010–5018
Acknowledgments
This study was supported by the National Natural Science Foundation of China (No. 30870948). We gratefully acknowledge Xinliang Ma (Department of Emergency Medicine, Thomas Jefferson University, USA), Feng Gao (Department of Physiology in our university), Hua Han (Department of Medical Genetics and Developmental Biology in our university) and Xiaoying Li (Institute of Endocrinology and Metabolism, Shanghai Jiaotong University School of Medicine, China) for their comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Rongrong Hou, Jing Zhang authors contributed equally to this work.
Rights and permissions
About this article
Cite this article
Hou, R., Zhang, J., Yin, T. et al. Upregulation of PTEN by peroxynitrite contributes to cytokine-induced apoptosis in pancreatic β-cells. Apoptosis 15, 877–886 (2010). https://doi.org/10.1007/s10495-010-0510-z
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10495-010-0510-z