
Abstract
The intertidal yeast communities inhabiting various environments in the territories of the White Sea Biological Station “Kartesh” (WSBS ZIN RAS) and the N.A. Pertsov White Sea Biological Station (WSBS MSU) were studied. A total of 31 yeast species were isolated using a conventional plating technique and identified using molecular methods. The yeast community of the White Sea intertidal zone consists of members that are typical for marine substrates, ubiquist species that are common in water and in low-temperature terrestrial environments, and a group of species that was isolated from marine substrates for the first time. The most diverse yeast communities formed on the surface of marine algae and in silt. Metschnikowia zobellii, which is a typical inhabitant of northern seas, was the most abundant yeast on algae from both biological stations. A new basidiomycetous yeast species, which was described in this work as Glaciozyma litorale sp. nov., dominated in the silt samples. The type strain of this new species is K94bT (=KBP 4246T = VKPM Y-3850T = PYCC 6252T = CBS 12957T = DSM 28204T); MycoBank registration number is MB 805475.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Yeasts are typically considered as inhabitants of sugar-rich substrates, but they also comprise a considerable proportion of the microbial population of plants, soils, freshwater and seawater (Fonseca and Inacio 2006; Botha 2006; Nagahama 2006). These types of habitats have not been studied thoroughly for various reasons. Recent reviews noted that marine yeasts are one of the most poorly studied microbial communities (Nagahama 2006; Kutty and Philip 2008), although fungi with yeast lifestyles are the dominant and most successful fungal form in seas, particularly deep seas (Bass et al. 2007).
Microbiological studies of marine yeasts began more than a century ago. One of the first studies in this field was a description of North Sea microbial communities that was published in 1894 (Fischer 1894). This study clearly showed that yeasts could be isolated from seawater. The first study of marine yeasts was performed in Russia by B.L. Isachenko, who isolated yeasts from seawater from the Barents Sea in 1914 (Isachenko 1914). In 1921–1922, G.A. Nadson first isolated yeasts from the surface of marine algae and demonstrated that the quantity of yeasts on algae was significantly higher than the quantity in the seawater (Nadson and Burgvits 1931). The most intensive studies of marine yeasts occurred in the 1940–1970s, after it became clear that yeasts are an essential component of the seawater microbial community (ZoBell 1946; Johnson and Sparrow 1961; van Uden and Castelo-Branco 1963; Fell and van Uden 1963; Goto et al. 1974; Fell 1976). During that period, the yeasts of the Black, Okhotsk, Caspian and Aral Seas, as well as the Pacific and Atlantic Oceans, were investigated in Russia and the former USSR (Kriss et al. 1952; Rukina and Novozhilova 1952; Kriss and Novozhilova 1954; Novozhilova 1955, 1979). Unfortunately, current studies of marine yeasts in Russia are nonexistent, despite some records of “huge amounts of yeasts” in seawater and other marine environments (Bubnova et al. 2008; Konovalova and Bubnova 2011).
The following conclusions can be drawn from the available data related to marine yeasts: (1) yeasts can be isolated from all layers of the water column, as deep as at 11,000 m; (2) the quantity and diversity of yeasts is larger in coastal areas; (3) yeasts may be used as a food source for sea animals; (4) yeasts form specific associations, including parasitic associations, with marine plants, algae and animals; (5) both autochthonous and allochthonous species of yeasts, including ubiquist species, can be found in seawater; (6) the major genera of yeasts that are isolated from seawater are Candida, Cryptococcus, Debaryomyces and Rhodotorula (Nagahama 2006; Kutty and Philip 2008).
The continuation of studies of the yeast community in northern seas is currently of interest to scientists. Although studies of the biology of marine yeasts started in northern seas, these seas have received much less scientific attention than southern and tropical seas. This is particularly true for Russian northern seas, where the most recent studies of yeasts occurred in the middle of the twentieth century. In the last 20 years, the taxonomy of yeasts has changed substantially; many new species have been described, and strains that were isolated decades ago have not been preserved. Consequently, a revisions of the marine yeasts from northern seas is an essential aim of research related to the biodiversity of yeasts.
In seas, coastal shallow-water biocenoses are more biodiverse than their deep-water counterparts. Therefore, the microbial community, including yeasts, that is associated with the intertidal biota could be more abundant and prolific, which has been demonstrated previously for southern seas (Seshadri and Sieburth 1971; de Araujo et al. 1995). Yeasts are generally more concentrated in intertidal sediment habitats than in water (Hagler et al. 1993; Soares et al. 1997). Thereby, analysis of the intertidal yeasts communities is applicable to an ecological research.
The aim of this work is to complete the marine research conducted in the northern part of Russia by providing a detailed overview of the intertidal yeast communities at The White Sea Biological Station “Kartesh” and the N.A. Pertsov White Sea Biological Station. The results of this study increase our knowledge about the association of yeasts with some typical intertidal substrates and samples of coastal seawater from the White Sea. We also report the isolation of novel yeast species, one of them is described in this paper as Glaciozyma litorale sp. nov.
Materials and methods
Study sites and sampling
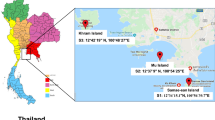
This study was performed at The White Sea Biological Station “Kartesh” (WSBS ZIN RAS) (approximate coordinates: 66.3394° N, 33.6596° E) in June 2011 and at the N.A. Pertsov White Sea Biological Station (WSBS MSU) (approximate coordinates: 66.5485° N, 33.1188° E) in August 2011. The White Sea belongs to the marginal seas of the Arctic Ocean. The biological stations are located on the shore of the Kandalaksha Gulf. This area is characterised by low environmental pollution and human land management. The majority of the Kandalaksha Gulf is included in The Kandalaksha State Nature Reserve. Average water temperatures in the gulf in January are between −0.5 and −0.7 °C, and ice covers the sea from early December to early May. The surface of the seawater warms up to 14–15 °C in August. The salinity of the water in the Kandalaksha Gulf does not exceed 20 ‰.
The investigation primarily focused on the algae Ascophyllum nodosum and Fucus vesiculosus. Samples (approx. 100 g per sample) were obtained from the intertidal zone at low tide. The algal samples from WSBS ZIN RAS were represented by a mixture of probes from the brown algae A. nodosum and F. vesiculosus. In addition, other types of substrates from the White Sea intertidal zone were analysed: algae (Odonthalia dentate and Saccharina latissima) collected at WSBS MSU and a sponge (Halichondria panicea) collected at WSBS ZIN RAS, as well as samples of silt, seawater and coastal sand. Algae and sponges collected for research were fresh and healthy. Seawater collection was performed directly in the field by filtering 10 or 20 ml of water through 0.22 µm membrane filters (Millipore), with subsequent fixation by air-drying. A total of 4–5 replicates were collected for each type of sample. After collection, the samples were transferred to sterile plastic bags, kept cool, and transported to the laboratory for analysis with in 2 days. The moisture content of the algae, sponges, silts and coastal sands were measured gravimetrically and calculated as the proportion of the original wet weight to the dry weight. These values were used to calculate the number of yeast cells (CFU) in each type of sample per its dry weight.
Isolation and identification of cultures
The quantity and taxonomic composition of yeasts were analysed by plating the samples on glucose-peptone-yeast extract (GPY) agar supplemented with chloramphenicol. After cutting the algal and sponge samples, three sub-samples (1 g each) were taken from each sample containing surface and internal parts of the algae and sponges, placed in a 50 ml plastic tube, suspended at a dilution of 1:30 (w/v) in sterile demineralised water, and vortexed for 20 min. The membrane filter was cut in twain, and each half was placed in a 50 ml plastic tube and suspended at a dilution of 1:10 (w/v). An aliquot of 0.1 ml was used for inoculation. Each sub-sample was plated in triplicate. To thoroughly investigate marine yeasts, some samples were also plated on other types of media: malt agar (MA), containing 3 % malt extract, which is a traditional medium for the isolation of fungi, dilute (10 %) GPY agar supplemented with 20 g/l of sea salt, which is an oligotrophic and salt-containing medium, and Luria–Bertani agar (LB), which is a medium for the isolation of proteolytic microorganisms, such as mycoparasites of sea animals. The plates were incubated at 10 °C for 1 month. Yeast colonies were differentiated into macromorphological types using a dissection microscope and counted. A total of 2–3 representatives of every colony type per sample were brought into pure cultures.
PCR-fingerprinting with microsatellite oligonucleotides (GTG)5 was used to group the pure cultures. Strains showing identical electrophoretic profiles were considered conspecific, and only 1–2 representatives of these strains were chosen for further identification using DNA barcoding. Yeast strains were identified by PCR amplification and sequencing of rDNA regions [i.e., the internal transcribed spacer (ITS) region and the D1/D2 domains of the large subunit (LSU) of rDNA]. The employed protocols for DNA extraction, PCR-fingerprinting, amplification of rDNA regions, purification and sequencing were previously described by Libkind et al. (2003), Kachalkin (2010) and Glushakova et al. (2010). To identify species, the nucleotide sequences were compared with sequences deposited in the NCBI (www.ncbi.nih.gov) and CBS (www.cbs.knaw.nl) databases. Nucleotide sequences from isolated species were deposited in GenBank under the accession numbers presented in Table 1.
Some isolated yeasts were deposited in The Yeast Collection of the Department of Soil Sciences at Lomonosov Moscow State University (KBP), The Russian National Collection of Industrial Microorganisms (VKPM), The Portuguese Yeast Culture Collection (PYCC), Leibniz Institute DSMZ–German Collection of Microorganisms and Cell Cultures (DSM) and The Centraalbureau voor Schimmelcultures (CBS).
Novel yeast species
A total of four strains representing the novel species of the genus Glaciozyma were studied. Phenotypic characterisation, including assimilation tests, of all isolates was performed as previously described by Kurtzman et al. (2011) at 10 °C. Mating tests and the production of hyphae and teliospores were performed on potato-dextrose agar (PDA), cornmeal agar (CMA), diluted (10 %) GPY agar, and water agar (WA). The germination of the teliospores was induced by incubating blocks of agar containing the teliospores in dH2O at 10 °C for 2–4 weeks and subsequently transferring the teliospores to WA or GPY agar.
Phylogenetic analysis was performed based on the nucleotide sequences of the internal transcribed spacers (ITS1–ITS2 regions), including the 5.8S, and the D1/D2 domains of the LSU regions of rDNA. The data were processed and analysed using DNA Baser Sequence Assembler (http://www.dnabaser.com), MAFFT 6 (Katoh et al. 2009) and MEGA 4 (Tamura et al. 2007) software.
Results
On average, approximately 72 colonies were obtained per plate in this study. This number allows for reliable counting and differentiation of colonies and for adequate isolation of representative strains for identification.
In total, 157 yeast strains were isolated during this study: 118 strains isolated on GPY agar and 39 strains isolated as a result of additional analysis of some samples using MA and oligotrophic media. A total of 24 different yeast species were isolated on GPY agar, including four undescribed species (Table 1). Using additional media, we were able to enrich the taxonomical list with seven more species: Candida vartiovaarae, Cryptococcus albidus, Debaryomyces hansenii, Rhodotorula laryngis, Rh. mucilaginosa, Rhodotorula sp. (Erythrobasidiales), and Sporidiobolus salmonicolor. Rhodotorula sp. (Erythrobasidiales) is undescribed yet species. Thus, altogether we isolated 31 yeast species, which belong to the following four lineages of fungi: Agaricomycotina (12), Pucciniomycotina (10), Saccharomycotina (8) and Pezizomycotina (1).
The quantity of yeasts in the White Sea intertidal zone varied based on substrate type; the highest number of cells was detected on algae, and the lowest number of cells was detected in seawater (Table 2). The highest number of yeasts among different algae was detected on the brown algae S. latissima [6.48 log10 (CFU/g, dry weight)]. Yeast counts did not differ significantly between brown and red algae, as A. nodosum and O. dentate were found to possess the same quantity of yeasts.
The most abundant yeast species isolated on different types of substrates from the White Sea intertidal zone differ considerably (Table 2). The novel species of the genus Glaciozyma were widespread in silt, Cryptococcus cf. oeirensis was widespread in seawater and Metschnikowia zobellii was widespread on algae. The yeasts isolated from coastal sand and sponges formed polydominant communities, without a single dominant yeast species characterizing many other substrates.
A comparison of the yeast taxonomic structure for substrates from the intertidal zones of the two biological stations revealed only three shared species: Aureobasidium pullulans, Cystofilobasidium sp. and M. zobellii. Algae and silt samples had the highest species richness among the different types of substrates; seawater had the lowest species richness (Table 2). The total numbers of species isolated from mixed samples of algae from WSBS ZIN RAS and from WSBS MSU were relatively close, and M. zobellii was the most abundant yeast species inhabiting the algae of the White Sea intertidal zone (Table 2).
Discussion
Glucose-peptone-yeast extract agar was used as the principal medium for yeast isolation and for the comparison of different substrates from the intertidal zone, as it has proven to be reliable in long-term studies of the yeast ecology and taxonomic composition of different soil and plant substrates (Chernov et al. 2013). GPY agar supports the isolation of species for which various selective media are frequently required (Yurkov et al. 2011). The results presented here demonstrate that many yeast species that are isolated on GPY agar can also grow on other types of media, including oligotrophic medium containing salts (Table 1). However, species isolated only on additional types of media have previously been isolated or able to grow on GPY; therefore, the increase in the number of isolated species was likely influenced by the additional analyses of some samples. It should be noted that GPY, which has high levels of peptone, is frequently used to isolate proteolytic yeast species; but results given in Table 1 showed isolation of various yeast species from different ecological groups. LB agar, as analog of GPY medium with salt and without glucose compounds, was used as an additional medium in this work for comparison of results and for selective isolation of proteolytic yeasts that could create close associations with marine animals.
A comparison of the results obtained from plating the samples on different types of media indicates that GPY agar and dilute GPY agar with sea salt are best suited for marine yeasts, according to the number of isolated species. This observation should be verified in future studies. LB agar did not support the isolation of mycoparasites of sea animals, such as some species of Metschnikowia.
Outside the intertidal zone and a sufficient distance from land, the average quantity of yeasts in seawater rarely exceeds 10–100 cells/l (Nagahama 2006). However, in coastal zones, in zoo- and phytoplankton-rich waters, and in the waters and soil of mangrove bromeliads, the quantity of yeasts can reach 103–105 cells/l (Fell and van Uden 1963; Novozhilova 1979; Hagler et al. 1993). The high quantity of yeasts in the seawater of White Sea intertidal zone is consistent with these previously published results. Our results confirm a previous observation of yeast quantity in the intertidal silt samples near 103 cells/g (Soares et al. 1997). The highest quantity of yeasts in the White Sea intertidal zone was observed for algae, particularly for the brown algae S. latissima (Table 2). Previous investigations of this algal species and of F. vesiculosus demonstrated that the quantity of yeasts on these algae was significantly higher than the quantity in the seawater (Nadson and Burgvits 1931). However, early investigations employed the elective cultivation method followed by inoculation, making quantitative estimations impossible.
The taxonomic structure of yeast communities from the White Sea intertidal zone includes species that are typical of marine habitats, as well as some species that are only occasionally isolated from seawater and some species which have not been previously found in marine habitats. Metschnikowia bicuspidata, M. zobellii and S. salmonicolor, which were isolated during this survey, are prevalent yeast species in marine environments (Novozhilova 1979; Nagahama 2006). Our results confirm previous reports that M. zobellii is the most abundant yeast species on marine algae (van Uden and Castelo-Branco 1961; Novozhilova 1979). This species was previously isolated from the alga A. nodosum in Norway, according to data from the CBS collection (strain CBS 7704). It should be noted that three closely related species of the genus Metschnikowia (M. australis, M. bicuspidata and M. zobellii) are prevalent in marine environments. However, some differences exist between these species. M. zobellii has been isolated from waters in the northern hemisphere, while M. australis is primarily found in the southern Antarctic region (Vaca et al. 2013). However, despite the frequent isolation of Metschnikowia species from seas and oceans, these species are not true autochthonous species of marine environments, as is often noted (Butinar et al. 2011). It is a well-known fact that M. bicuspidata and M. zobellii are mycoparasites of arthropods that inhabit seas (i.e., Daphnia magna), saline water (i.e., Artemia salina) and freshwater (i.e., Daphnia dentifera and Macrobrachium rosenbergii) (van Uden and Castelo-Branco 1961; Chen et al. 2007; Duffy and Sivars-Becker 2007). S. salmonicolor is also a prevalent yeast in marine environments and is sometimes isolated from soils and plants (Sampaio 2011).
A significant number of yeasts isolated from the White Sea intertidal zone are not substrate-confined. However, with the exclusion of the species discussed above, the observed yeast community could be characterised as more typical of low-temperature habitats. A. pullulans, Candida sake, Cryptococcus adeliensis, Cr. victoriae, Leucosporidium scottii, Mrakia frigida, and R. laryngis are typical inhabitants of soils and plants in northern regions of Russia, as well as glaciers and soils of highlands and Arctic and Antarctic territories (Babjeva and Reshetova 1998; Branda et al. 2010). Other species have also been previously noted in lists of yeasts from permanently or temporarily cold environments (Butinar et al. 2007; Chernov et al. 2013). An important feature of the White Sea intertidal yeast community is that about one-third of the isolated species are ascomycetes, which are not abundant in soils and plants of northern regions or in other cold terrestrial environments (Branda et al. 2010). In our opinion, the most likely cause for the broad existence of this group in the samples is a wide distribution of mucus-covered algae in the intertidal zone. Algal mucus, which is rich in polysaccharides (e.g., agarose, fucoidan, and carrageenan) and contains mannose, galactose, glucose, xylose, and rhamnose as minor monosaccharides (Zhang et al. 2009), is a suitable substrate for the growth of yeast communities with a high proportion of ascomycetes. This theory was supported by comparing yeast abundance in different types of samples; five of the seven ascomycetes were isolated from algae (Table 2).
Another group of yeast species that was isolated and identified during this study had not been previously found in marine habitats. First, we report the second isolation of the species Leucosporidium drummii from coastal sand, but this habitat can be partly assigned as a marine habitat. The newly reported yeasts from marine habitats are Leucosporidiella fragaria, which is a minor component of soil and plant environments, and five other species representing undescribed species. The yeast Cystofilobasidium sp. (strains KBP 4391 = VKPM Y-3909 and KBP 4384 = VKPM Y-3908), which is closely related to Cystofilobasidium bisporidii, is a new undescribed yet species isolated from two biological stations. The conspecific strain Cystofilobasidium sp. EX-F 1547 (DQ640493) was previously found in Arctic glaciers. Saprochaete sp. (strain KBP 4390 = VKPM Y-3915) is an undescribed yet species that was isolated as minor component of the yeast community on O. dentate. This species is genetically most closely related to Saprochaete suaveolens (former syn. Geotrichum fragrans) and has the conspecific strain Geotrichum sp. NCAIM Y.01895 (GQ258823). Cryptococcus sp. (strain 93b) from the Cr. victoriae clade is an undescribed yet species with a small number of conspecific strains, such as Cryptococcus sp. CBS 11772, which was isolated from mud sedge in Russia, and other strains from different natural habitats and from industry. During the current study, this species was isolated from silt. Rhodotorula sp. (strain 31-1), which is closely related to R. laryngis, is a new undescribed yet species that was isolated from water. Other conspecific strains of Rhodotorula sp. were isolated from Antarctic (strain CBS 8923), Alpine (strains DBVPG 5748 and DBVPG 10053) and Tibetan (JQ768910) glaciers and from swamp plants in the Moscow region in the winter–spring season. Finally, four strains of a new yeast species in the genera Glaciozyma were isolated as dominants in the yeast community in silt from the White Sea intertidal zone. This genus, which includes three psychrophilic species, was proposed and described in 2011 (Turchetti et al. 2011). All strains of these species were isolated from seawater, Antarctic soil or Alpine glaciers and are unable to maintain growth at temperatures greater than 17–20 °C.
The high diversity of yeasts in the White Sea intertidal zone supported the isolation of five undescribed yet or new species, which represent 16 % of the species identified in the current study. Thus, this study aims to support the continued study of yeast biology in the White Sea and other northern seas of Russia.
Novel yeast species description
In our study of yeast taxonomic composition at The White Sea Biological Station “Kartesh” (WSBS ZIN RAS), four strains of a new yeast species were isolated. These strains belong to a new species of the genus Glaciozyma and were isolated from silt, the alga A. nodosum and coastal sand. The genus Glaciozyma (Camptobasidiaceae, Kriegeriales, Microbotryomycetes, Pucciniomycotina) contains three species: G. antarctica, G. martinii and G. watsonii. rDNA-based phylogenetic analysis of two strains from the White Sea intertidal zone indicated that these strains belong to a single species and that these strains are sufficiently different from other species of the genus Glaciozyma to comprise a new species (Fig. 1). The nearest sequence match among the currently recognised species was obtained with G. martinii, which showed an approx. 11 % difference (i.e., 13 nucleotide substitutions and 54 gaps) in the ITS1–5.8S–ITS2 region and an approx. 3 % difference (i.e., 16 nucleotide substitutions) in the D1/D2 domains of the LSU rDNA. The remaining two isolated strains have rDNA sequences identical to the strains represented in the analysis.

Maximum parsimony tree based on combined ITS1–5.8S–ITS2 region and D1/D2 domains of the LSU rDNA sequences for Glaciozyma litorale sp. nov. The numbers given on branches are frequencies (>50 %) with which a given branch appeared in 1,000 bootstrap replications. The scale bar indicates the number of nucleotide substitutions. The tree is rooted with Rhodotorula minuta CBS 319T
Varying results of physiological tests between species and between strains are a common trait of the genus Glaciozyma (Turchetti et al. 2011). The d-galactose (i.e., no growth) and salicin (i.e., growth) physiological tests are good markers for the distinction of the new species from the other species of the genus Glaciozyma. Variable results were obtained from five physiological tests for strains isolated from the White Sea intertidal zone. Such results could be caused by some genomic variability of strains, as demonstrated by PCR fingerprinting with microsatellite (Fig. S1 in Supplementary material).
After 3–7 days of incubation on PDA, all of the strains of the new species began formation of pseudomycelium containing lageniform cells and short segments of true hyphae. The new species formed structures similar to teliospores, developing from lageniform cells, with intercalar, lateral or terminal positions on pseudomycelium and on true hyphae (Fig. 2c–e). Teliospores formed with the same morphology after mating tests and after cultivation of single strains. Clamp connections were not observed. Similarly, the three other species of the genus Glaciozyma are characterised by the ability to produce teliospores with or without clamp connections (Turchetti et al. 2011). The new species is morphologically similar to G. antarctica (formerly Leucosporidium antarcticum), which forms teliospores without clamp connections. The germination of the teliospores of the new species was induced by a 3-week incubation in dH2O at 10 °C, with subsequent transfer to GPY agar or WA. Selective germination started after 2 days of cultivation on GPY agar (Fig. 2e) and after 15 days of cultivation on WA (Fig. 2f–h). Teliospore germination produced only yeast cells or pseudomycelium. Similarly, teliospore germination of the species G. antarctica and G. watsonii produces pseudomycelium without structures that could be considered basidia.

Light microscopic images illustrating the different stages of Glaciozyma litorale sp. nov. a, b Budding cells; c, d, e hyphae, pseudohyphae and teliospores; e–h germinated teliospores. Bars 10 μm
Description of Glaciozyma litorale sp. nov. Kachalkin
Glaciozyma litorale (li.to.rále. L. neut. adj. litorale of the shore, dweller of the intertidal zone).
On PDA, after 1 month at 10 °C streak culture is white, flat, butyrous glistening, homogeneous consistency, the surface is undulate with the sinuous margin that fringing with filaments into medium. On GPY agar streak culture is yellowish-white. After growth on PDA and GPY agar for 3 days at 10 °C, cells are short ovoidal to elongate 4–6 × 8–25 μm and proliferate by polar budding (Fig. 2a); short pseudohyphae are frequently present. After growth on PDA for 14 days at 10 °C, cells are mostly ovoidal with 2–4 lipid bodies (Fig. 2b); pseudo- and true hyphae are present (Fig. 2c–d). Hyphae are 1.5–6 μm in diameter; clamp connections were not observed. Teliospores formations from pseudohyphae cells started on PDA after 3 days. After true hyphae formations teliospores appear on it. Teliospores are spherical, 8.5–11 μm in diameter, terminal, lateral or intercalary, often formed short chains of 2–4 spores (Fig. 2c–e). Teliospores germinate selectively by forming pseudohyphae (Fig. 2e–h).
Glucose is not fermented. Assimilates the following carbon compounds: d-glucose, cellobiose (slow and variable), trehalose (slow and variable), ethanol (slow), glycerol (slow), d-mannitol, salicin, dl-lactic acid (slow and variable), succinic acid (slow and variable), citric acid (slow and variable), saccharic acid (slow), mucic acid (slow), 2-ketogluconic acid (slow). No growth occurs on d-galactose, l-sorbose, sucrose, maltose, lactose, melibiose, raffinose, melezitose, inulin, starch, d-xylose, l-arabinose, d-arabinose, d-ribose, l-rhamnose, d-glucosamine, erythritol, ribitol, dulcitol (galactitol), d-sorbitol (d-glucitol), methyl-α-d-glucoside, myo-inositol, 5-ketogluconic acid, arbutin, d-glucuronate, protocatechuic acid, gentisic acid, gallic acid, p-hydroxybenzoic acid, m-hydroxybenzoic acid, salicylic acid, ferulic acid, vanillic acid, veratric acid, l-tartaric acid. Potassium nitrate is not assimilated. Growth on YM agar with 10 % sodium chloride is negative. Growth in 50 % glucose/yeast extract (0.5 %) is negative. Maximal growth temperature: 20–21 °C. Positive for urease activity. Diazonium blue B reaction is positive. Starch-like compounds are not produced.
The type strain is K94b (=KBP 4246T = VKPM Y-3850T = PYCC 6252T = CBS 12957T = DSM 28204T) isolated from silty-sandy substrate of intertidal zone of the White Sea (Russia). The sequence of the ITS (ITS1–5.8S–ITS2) region and D1/D2 domains of 26S (LSU) rDNA was deposited in GenBank under the accession number HF934009. The new species was registered in MycoBank under MB 805475.
References
Babjeva I, Reshetova I (1998) Yeast resources in natural habitats at polar circle latitude. Food Technol Biotechnol 36:1–5
Bass D, Howe A, Brown N, Barton H et al (2007) Yeast forms dominate fungal diversity in the deep oceans. Proc Biol Sci 274:3069–3077
Botha A (2006) Yeasts in soil. In: Rosa CA, Peter G (eds) Biodiversity and ecophysiology of yeasts. The yeast handbook. Springer-Verlag, Berlin, pp 221–240
Branda E, Turchetti B, Diolaiuti G, Pecci M et al (2010) Yeast and yeast-like diversity in the southernmost glacier of Europe (Calderone Glacier, Apennines, Italy). FEMS Microbiol Ecol 72:354–369
Bubnova EN, Kireev YV, Konovalova OP, Porhunova NN (2008) Fungi of the White Sea. In: The conference dedicated to 70th anniversary of the N.A. Pertsov White Sea Biological Station. Grif I K, Tula, pp 217–223
Butinar L, Spencer-Martins I, Gunde-Cimerman N (2007) Yeasts in high Arctic glaciers: the discovery of a new habitat for eukaryotic microorganisms. Antonie Van Leeuwenhoek 91:277–289
Butinar L, Strmole T, Gunde-Cimerman N (2011) Relative incidence of ascomycetous yeasts in arctic coastal environments. Microb Ecol 61(4):832–843
Chen SC, Chen YC, Kwang J, Manopo I et al (2007) Metschnikowia bicuspidata dominates in Taiwanese cold-weather yeast infections of Macrobrachium rosenbergii. Dis Aquat Organ 75:191–199
Chernov IY, Glushakova AM, Kachalkin AV (2013) Annotated list of yeasts from Moscow region. Mycol Phytopathol 47:103–115
de Almeida JM (2005) Yeast community survey in the Tagus estuary. FEMS Microbiol Ecol 53(2):295–303
de Araujo FV, Soares CA, Hagler AN, Mendonça-Hagler LC (1995) Ascomycetous yeast communities of marine invertebrates in a southeast Brazilian mangrove ecosystem. Antonie Van Leeuwenhoek 68(2):91–99
Duffy MA, Sivars-Becker L (2007) Rapid evolution and ecological host-parasite dynamics. Ecol Lett 10:44–53
Fell JW (1976) Yeasts in oceanic regions. In: Jones EBG (ed) Recent advances in aquatic mycology. Elek Science, London, pp 93–124
Fell JW, van Uden N (1963) Yeasts in marine environments. In: Oppenheimer CH (ed) Symposium on marine microbiology. Charles C. Thomas, Springfield, pp 329–340
Fischer B (1894) Die Bakterien des Meeres nach den Untersuchungen der Plankton-Expedition unter gleichzeitinger Berucksichtigung einiger alterer und neurer Untersuchungen. Ergebnisse der Plankton-Expedition der Humboldt-Stiftung, bd. 4
Fonseca A, Inacio J (2006) Phylloplane yeasts. In: Rosa CA, Peter G (eds) Biodiversity and ecophysiology of yeasts. The yeast handbook. Springer-Verlag, Berlin, pp 263–303
Glushakova A, Maximova I, Kachalkin A, Yurkov A (2010) Ogataea cecidiorum sp. nov., a methanol-assimilating yeast isolated from galls on willow leaves. Antonie Van Leeuwenhoek 98:93–101
Goto S, Yamasato K, Iizaka H (1974) Identification of yeasts isolated from the Pacific Ocean. J Gen Appl Microbiol 20:309–316
Hagler AN, Rosa CA, Morais PB, Mendonca-Hagler LC et al (1993) Yeasts and coliform bacteria of water accumulated in bromeliads of mangrove and sand dune ecosystems of southeast Brazil. Can J Microbiol 39:973–977
Isachenko BL (1914) Studies of bacteria of the Arctic Ocean. In: Murmansk Scientific Fishery Expedition, 1906. Petrograd.
Johnson TW, Sparrow FK (1961) Fungi in oceans and estuaries. J. Cramer, Weinheim
Kachalkin AV (2010) New data on the distribution of certain psychrophilic yeasts in Moscow oblast. Microbiology 79:840–844
Katoh K, Asimenos G, Toh H (2009) Multiple alignment of DNA sequences with MAFFT. Bioinformatics for DNA sequence analysis. Methods Mol Biol 537:39–64
Konovalova OP, Bubnova EN (2011) Fungi on brown seaweeds Ascophyllum nodosum and Pelvetia canaliculata in the Kandalaksha bay of White sea. Mycol Phytopathol 45:240–248
Kriss AE, Novozhilova MN (1954) Are yeasts inhabitants of seas and oceans? Microbiology 23:669–683
Kriss AE, Rukina EA, Tikhonenko AS (1952) A distribution of yeasts in the sea. Zh Obshch Biol 13:242–252
Kurtzman CP, Fell JW, Boekhout T, Robert V (2011) Methods for isolation, phenotypic characterization and maintenance of yeasts. In: Kurtzman CP, Fell JW, Boekhout T (eds) The yeasts, a taxonomic study, vol 1, 5th edn. Elsevier, Amsterdam, pp 87–111
Kutty SN, Philip R (2008) Marine yeasts—a review. Yeast 25:465–483
Libkind D, Brizzio S, Ruffini A, Gadanho M et al (2003) Molecular characterization of carotenogenic yeasts from aquatic environments in Patagonia, Argentina. Antonie Van Leeuwenhoek 84(4):313–322
Nadson GA, Burgvits GK (1931) Yeast of the Arctic ocean. Dokl Akad Nauk SSSR 4:103–110
Nagahama T (2006) Yeast biodiversity in freshwater, marine and deep-sea environments. In: Rosa CA, Peter G (eds) Biodiversity and ecophysiology of yeasts. The yeast handbook. Springer-Verlag, Berlin, pp 241–263
Novozhilova MI (1955) The quantitative characteristics, species composition and distribution of yeast like organisms in the Black Sea, the Sea of Okhotsk and the Pacific Ocean. Tr Inst Mikrobiol Akad Nauk SSSR 4:155–195
Novozhilova MI (1979) Asporogenous yeasts and their roles in waters. Nauka, Alma-Ata
Rukina YA, Novozhilova MI (1952) Species composition of yeast organisms isolated from various depths of the Black Sea. Tr Inst Mikrobiol Akad Nauk SSSR 2:150–156
Sampaio JP (2011) Sporidiobolus Nyland (1949). In: Kurtzman CP, Fell JW, Boekhout T (eds) The yeasts, a taxonomic study, vol 3, 5th edn. Elsevier, Amsterdam, pp 1549–1561
Seshadri R, Sieburth JM (1971) Cultural estimation of yeasts on seaweeds. Appl Microbiol 22(4):507–512
Soares CA, Maury M, Pagnocca FC, Araujo FV et al (1997) Ascomycetous yeasts from tropical intertidal dark mud of southeast Brazilian estuaries. J Gen Appl Microbiol 43(5):265–272
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol Biol Evol 24:1596–1599
Turchetti B, Thomas Hall SR, Connell LB, Branda E et al (2011) Psychrophilic yeasts from Antarctica and European glaciers: description of Glaciozyma gen. nov., Glaciozyma martinii sp. nov. and Glaciozyma watsonii sp. nov. Extremophiles 15:573–586
Vaca I, Faúndez C, Maza F, Paillavil B et al (2013) Cultivable psychrotolerant yeasts associated with Antarctic marine sponges. World J Microbiol Biotechnol 29:183–189
van Uden N, Castelo-Branco R (1961) Metschnikowia zobellii sp. nov. and M. krissii sp. nov., two yeasts from the Pacific Ocean pathogenic for Daphnia magna. J Gen Microbiol 26:141–148
van Uden N, Castelo-Branco R (1963) Distribution and population densities of yeast species in Pacific water, air, animals, and kelp off southern California. Limnol Oceanogr 8(3):323–329
Yurkov AM, Kemler M, Begerow D (2011) Species accumulation curves and incidence-based species richness estimators to appraise the diversity of cultivable yeasts from beech forest soils. PLoS ONE 6(8):e23671
Zhang JJ, Zhang QB, Wang J, Shi XL, Zhang ZS (2009) Analysis of monosaccharide compositions in fucoidan by precolumn derivation HPLC. Chin J Oceanol Limnol 27:578–582
ZoBell CE (1946) Marine microbiology. Waltham Mass, USA
Acknowledgments
The author is grateful to O. E. Marfenina (Lomonosov Moscow State University, Russia), M. A. Velikanova and T. V. Artyshkina (Research Institute for Genetics and Selection of Industrial Microorganisms, Russia), who contributed samples for this study. The author wishes to thank M. M. Vustin (Research Institute for Genetics and Selection of Industrial Microorganisms, Russia) and A. M. Yurkov (Leibniz-Institute DSMZ, Germany) for valuable suggestions on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Kachalkin, A.V. Yeasts of the White Sea intertidal zone and description of Glaciozyma litorale sp. nov.. Antonie van Leeuwenhoek 105, 1073–1083 (2014). https://doi.org/10.1007/s10482-014-0165-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10482-014-0165-9