
Abstract
The growth rate of forest trees indicates the rate of wood production and overall tree health. Tree growth also indicates resource distribution and the effect of ecological variables. Silvicultural treatments may improve the growth and resilience of trees growing in dehesas (Mediterranean open-woodland forests). These ecosystems are expected to experience increased temperature and decreased precipitation due to global climate change; hence their management is a key factor contributing to the adaption, and hence the conservation of these systems. In this paper, we analyzed the effect of traditional silvicultural treatments on the intra-annual stem growth of holm oak (Quercus ilex L.) in three dehesa plots in SW Spain: (i) soil treatments (ploughing, or ploughing + fertilization with calcium superphosphate + sowing with the legume yellow lupin, Lupinus luteus) and (ii) pruning intensity (heavy, moderate, or light). The soil treatments had no significant effects on growth, but pruning significantly affected growth patterns. Heavy pruning slightly reduced growth during the spring, and moderate and heavy pruning increased the normal stem contractions (due to water stress during drought) and expansions (due to rehydration after drought) in sites with poorly developed soils or other stress causes. Hence, heavy pruning could affect the vigor and vegetative status of trees in areas where tree survival is already compromised. Light pruning did not affect tree growth, so this treatment may be acceptable if the extraction of firewood or biomass is one of the management objectives.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Growth is an important biological parameter of forestry species because it indicates the amount of wood production and overall tree health. Growth is also an indicator of resource distribution, because it influences phenological processes and indicates response to ecological variables, such as climate and soil (Fritts 1976). Therefore, knowledge of tree growth processes and their ecological determinants is a key issue for planning sustainable management programs in the presence of global climate change.
Holm oak (Quercus ilex L.) is a widespread species in the Mediterranean Basin and covers more than 6.5 million ha in total (Quézel and Médail 2003), including 3 million ha in the Iberian Peninsula (Bravo et al. 2008). It is one of the dominant species in “dehesas”, traditional agroforestry systems composed of an open woodland forest (10–60 trees ha–1) and an herbaceous layer (Cubera and Moreno 2007a). Dehesas are widely distributed in Spain and cover about 3–3.5 million ha (San Miguel 1994). Previous studies have shown that the stem growth of holm oak is strongly driven by climate, especially water availability (Campelo et al. 2009; Gea-Izquierdo et al. 2011; Martín et al. 2014). Improvement of soil and stand characteristics by certain management practices such as ploughing, fertilization or pruning, could increase the growth and resilience of holm oak in Mediterranean open woodland forests, which are expected to experience increased temperature and decreased precipitation in the coming years (IPCC 2007). However, it is unclear whether the traditional management practices applied in Mediterranean open woodland forests enhance or reduce tree growth.
Ploughing is one of the most common soil treatments traditionally performed in dehesas. Ploughing improves water infiltration and movement of organic matter and also removes woody shrubs and parched herbaceous plants, allowing the development of new pasture (Eichhorn et al. 2006). Nevertheless, in the Mediterranean area, Ploughing also increases the vulnerability of soils to erosion and negatively influences tree regeneration because it removes oak seedlings and the shrub species that can protect them (Pulido and Díaz 2005). Fertilization is also commonly applied in dehesas to enhance pasture and crops production. N and P are the most limiting nutrients in Mediterranean ecosystems (Gallardo et al. 2009) so their addition is widely used in the Mediterranean area to enhance pasture production. On the other hand, because of soil acidity, dehesas respond positively to Ca fertilization (San Miguel 1994). Sowing of leguminous species, such as yellow lupin (Lupinus luteus) is also frequently applied because these species can be harvested as livestock feed (Serrano et al. 2010) and they fix atmospheric N2. However, previous studies in diverse forests and open woodlands throughout the world have showed mixed and contradictory results of the response of tree growth to fertilization and atmospheric nitrogen fixation (e.g., McMaster et al. 1982; Cartan-Son et al. 1992; Scowcroft et al. 2007; Markewitz et al. 2012).
Pruning is a widespread multipurpose silvicultural practice applied to a large variety of tree species in distant regions (e.g., Pinkard and Beadle 1998; Balandier et al. 2000; Gyenge et al. 2010). Holm oak trees have traditionally been subjected to crown pruning for collection of fuelwood and improvement of acorn production (Alejano et al. 2011). Nevertheless, several researchers have questioned the benefits of pruning on acorn production (e.g., Cañellas et al. 2007; Alejano et al. 2008, 2011). This objection, along with the decreasing market value of fuelwood and the possibly negative influence of pruning on the spread of phytopathogenic agents involved in the “oak decline” of SW Spain (see Navarro 2011), have motivated reconsideration of the usefulness of pruning in holm oak. On the other hand, pruned trees are more sensitivity to water stress in dry regions (Gyenge et al. 2009). Hence, pruning could increase the vulnerability of holm oak to climate change, leading to a higher tree dieback.
Currently, the main management challenges of Mediterranean forest systems, and especially dehesas, is the survival and maintenance of productivity in the presence of oak decline, climate change and misuse of resources (Navarro 2011). In a former study (Martín et al. 2014), we studied the patterns of intra-annual stem growth of holm oak in open woodland forest and dense Mediterranean forests of SW Spain, and assessed the effects of climate, competition, tree size and microecological factors on stem increments. Now, the aim of the present study is to answer the following research questions:
-
(i)
Do the traditional soil management practices performed in open woodlands have a beneficial effect on the intra-annual stem growth of holm oak in this region?
-
(ii)
Do the pruning treatments have a detrimental effect on stem growth?
Our results will increase the basic scientific knowledge of Mediterranean open woodland forests and will therefore contribute to the development of methods for sustainable management based on established scientific criteria.
Materials and methods
Field plots
This study was performed in three experimental plots in the Huelva province of SW Spain (Table 1).
The Huerto Ramirez (HR) plot was in an open woodland of holm oak where sheep and Iberian pig were raised. Its soils had different degrees of development from acrisols, alisols and lixisols, to regosols and cambisols (IUSS Working Group WRB 2007). There was a sparse understory of mainly rock-rose (Cistus ladanifer) and curly rockrose (Cistus crispus) and an abundant herbaceous layer of mainly grasses. The San Bartolomé (SB) plot was in an open woodland of holm oak where bulls were raised. Its soils were endoleptic regosols (episkeletic) or endoleptic luvisols (dystric) in depositional or concave areas (IUSS Working Group WRB 2007). There was a very scarce understory due to frequent tillage, and an abundant herbaceous layer of mainly grasses. The Calañas (CA) plot was in an open woodland of holm oak with a highly developed understory layer composed of rock-rose and Montpelier cistus (Cistus monspeliensis) that was mainly used for hunting and extensive sheep herding. Its soils had a low profile development and were classified as cambisols (dystric, chromic) and leptic regosols (IUSS Working Group WRB 2007). In CA symptoms of oak decline were detected in three trees during the study and dendrometer data of these trees were removed.
Age of trees was very difficult to assess because it requires cutting trees to obtain cross sections for accurately estimate this data (Gea-Izquierdo et al. 2011). Even though we don’t have information for assessing the age structure of the plots, measurements in 30 sections in CA plot gave a result of an average age of 95 ± 12 years (oldest tree 113 years) and the measurement in 12 sections in HR plot gave a result of an average age of 110 ± 31 (oldest tree 149 years) (Natalini et al. 2013).
The climate of all three plots is Mediterranean, with highly variable temperature and rainfall within and among years. The mean annual temperature in the study period (2003–2011) was 17.1 °C in HR, 17.3 °C in SB and 17.7 °C in CA. Mean annual precipitation was 578 mm in HR, 655 mm in SB and 623 mm in CA (data from the Andalusian agroclimatic weather stations network). There were no large monthly variations in temperature across years, but there were large monthly and annual changes in precipitation. In particular, the annual precipitation in CA was only 306 mm during 2005, but was 1019 mm in SB during 2010. A more detailed description of the climate of this location is available in Martín et al. (2014).
Soil treatments
The HR plot was fenced and divided into nine subplots, with a mean area of 3340 ± 305 m2 and including 20–25 trees per subplot. Two soil treatments were applied randomly to three subplots each during autumn of 2005 and repeated during autumn of 2008: (i) ploughing (PL) or (ii) ploughing + fertilization + yellow lupin sowing (PFS). The other three subplots were used as controls. All PL was done with a disk harrow on a farm tractor. For the PFS treatment, PL was performed as in the PL treatment, and the soil was then fertilized with calcium superphosphate (P2O5 18 %; CaO 28 %; SO3 25 %) and sowed with yellow lupin using a mechanical seeder. Calcium superphosphate increases soil P without lowering pH, because CaO reduces the acidifying effect of phosphate. Yellow lupin provides soil N without the need for artificial fertilization. This species fixes atmospheric N2, is well adapted to local conditions, and is traditionally grown in this area. A total of 300 kg ha−1 of fertilizer (54 kg ha−1 of P2O5, 84 kg ha−1 of CaO, 75 kg ha−1 of SO3) and 60 kg ha−1 of seeds were used on each treatment date (100 kg of fertilizer and 20 kg of seeds in each PFS subplot, approximately). These particular treatments are traditional soil practices that have been frequently used in the dehesas of SW Spain. Control plots were progressively covered by an understory of shrubs, mainly rock-rose.
Pruning treatments
Trees in the CA and SB plots were subjected to traditional pruning at three different intensities (light, moderate, or heavy) or were left unpruned (controls). Pruning was performed in January 2001 in the CA plot and in February 2003 in the SB plot. All trees had been pruned every 6–7 years before we began our study. Pruning treatments were randomly assigned to trees at both sites. The pruning intensity was established as follows: light pruning involved removing sucker and dead branches only; heavy pruning, much stronger, removed up to 1/3 of the crown volume cutting branches thinner than 15 cm diameter and coincided with the usual practice in the area; and moderate pruning was in between the previous two. In order to ensure homogeneity in treatment intensity, each worker was assigned a single treatment type and supervised by a member of the research group. Once pruning was completed, pruning intensity was estimated based on the dry weight in kg of pruned branches (DW) and tree diameter in cm (D), as DW/D. A DW/D of 0.8 kg cm −1 or less was considered light pruning, a DW/D greater than 1.7 kg cm−1 was considered heavy pruning, and intermediate DW/D values were considered moderate pruning. Details of the estimation of pruning intensities are available in Alejano et al. (2008).
Measurement of stem growth
A total of 119 aluminum band dendrometers (system developed by the University of Huelva) were installed at breast height (1.30 m), with care taken to avoid stem deformities. Details of band dendrometers theory and construction are available in Keeland and Young (2014). Trees were randomly sampled within each pruning or soil treatment. There were 55 trees in HR (18 control, 19 PL and 18 PFS), 32 trees in SB, and 32 trees in CA (8 control and 8 by pruning intensity in each plot). Measurements of every band dendrometer were recorded each month with a digital caliper (0.01 mm accuracy) from 2003–2011 in SB, from 2003–2006 in CA, and from 2006–2011 in HR. Because there were differences in measurement dates and in the number of days per month, average daily increments for each tree between the first day and the last day of each month were calculated. Girth increment data were not transformed into diameter increment because holm oak is a species with high within-tree variability in stem growth and then the stems were not enough cylindrical to assume diameter transformation. Hence, girth increment data of entire cross-sections were used instead of diameter increment. At the beginning of the study, the topographic location of each of the 119 trees was measured using a total station (Sokkia 3B).
Data analysis
We used two different models to analyze the effect of different pruning treatments on growth in the CA and SB plots and the effect of different soil treatments on growth in the HR plot. Each model was a linear mixed model in which we considered tree as a random effect and the following fixed effects: month, year, treatment (pruning or soil treatment), plot (only in the pruning experiment), and all interactions. Thus, the initial structure of the soil treatment model was:
where y ijlm is the girth increase (mm day−1) of tree i in month j of year l under soil treatment m; μ is the general mean; b i is a tree random effect with i = 1, 2,…, 55 under the hypothesis b i ∼ N(0, G); αj is a month fixed-effect with j = 1, 2,…,12; γ l is a year fixed-effect with l = 1, 2,…, 6; τ m is a treatment fixed-effect with m = 1, 2, 3; (α|γ|τ) jlm is all possible double and triple interactions between fixed effects; and e ijlm is the residual error under the hypothesis e ijlm ∼ N(0, R).
The initial structure of the pruning treatment model is:
here y iklmt is the girth increase (mm day−1) of tree i at plot k in the month l of year m under pruning treatment t; μ is the general mean; b i(k) is a tree random effect within each plot with i = 1, 2,…, 32 and k = 1, 2 under the hypothesis b i(k) ∼ N(0, G); α κ is a plot fixed-effect with k = 1, 2; γ l is a month fixed-effect with l = 1, 2,…, 12; τ m is a year fixed-effect with m = 1, 2,…, 8; λ t is the treatment fixed effect with t = 1, 2, 3, 4; \(\left(\vert \alpha \vert \gamma \vert \tau \vert \lambda\right)_{klmt}\) is all possible double, triple and quadruple interactions between fixed effects; and e iklmt is residual error under the hypothesis e iklmt ∼ N(0, R).
The following procedure was used to select the best model structure:
-
1.
The models were adjusted by consideration of tree random effect, the presence of temporal correlations between observations of different months for each tree and year, and the presence of heterogeneous variances in different months of the year. Hence, G was initially considered as a diagonal matrix and R as a block diagonal matrix, with each block corresponding to a 12 × 12 submatrix of observations for one year in each tree. We considered the following alternatives for the structure of blocks in the R matrix: autoregressive order 1, autoregressive heterogeneous, Toeplitz up to five bands, heterogeneous Toeplitz up to five bands, unstructured up to five bands, Huynh–Feldt, compound symmetry, compound symmetry heterogeneous, dependent covariance, and first-order factor analytic (Littell et al. 2006). Variance components for each structure were estimated by restricted maximum likelihood (REML) (Patterson and Thompson 1971) and model selection was based on the Akaike information criterion (AIC, Akaike 1974).
-
2.
The significance of the tree random effect was determined by a likelihood ratio test, as the reduction of the statistic −2 × log likelihood (−2LL), after introducing the tree random effect, follows a χ2 distribution with 1 degree of freedom. A α value of 0.05 was considered to indicate an improvement in the covariance structure.
-
3.
If the tree random effect was significant, the presence of spatial correlation was determined. In particular, the following isotropic power covariance model was used:
$$\text{cov} (b_{i(j)} ,b_{i^\prime (j)} ) = \sigma^{2}_{b} \rho^{dii^\prime }$$(3)where \(di{i^\prime}\) is the distance between trees i and \({i^\prime}\) in location j; σ2b is the variance component at tree level; and ρ is a parameter to be estimated with |ρ| < 1. The spatial covariance between observations at different locations in the pruning treatment model was considered zero.
-
4.
After selection of the best variance–covariance structure, the fixed effects were estimated by a generalized least squares equation (Searle 1971) and the significance of each effect was determined with an F test. The significance of all differences was determined with the Scheffe´ test. All statistical analysis was performed with SAS/ETS (ver. 9.2).
Results
Soil treatments
The best structure of the variance–covariance matrix for the 12 × 12 blocks of the R matrix was unstructured with four bands. This indicates that the variance of observations was different in different months (heterogeneous structure) and that there was a temporal correlation for groups in four consecutive months. The tree random effect was highly significant (p < 0.0001), but spatial covariance was not (p = 0.934), indicating the presence of significant growth differences among trees but that these differences cannot be explained by tree location within the plots.
The results of the soil treatment model indicates that there were no significant differences in growth among the different soil treatment groups or in any interaction in which soil treatment is included (Table 2).
Pruning treatments
The best structure of the variance–covariance matrix for the 12 × 12 blocks of the R matrix was unstructured with two bands. We found a highly significant tree effect (p < 0.0001), but could not determine the presence of spatial covariance due the lack of convergence.
The selected model indicated that pruning treatment was not significant, although there were significant treatment × month and treatment × month × plot interactions (Table 3). In other words, this model indicates that pruning is associated with significant differences in growth during some months, and that plots behave differently under some treatments during some months. These two interactions only account for a small amount of the total variance (Table 4), and in some months is even negative, indicating that variance does not decrease after the introduction of this interaction in some months. However, in July, October, and November the treatment × month × plot effect explained 1.09–3.94 % of the residual variance, and in October the treatment × month effect explained 1 % of the variance (Table 4).
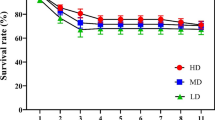
Growth differences throughout the year were more evident in the CA plot (Fig. 1). For example, in July there was greater stem contraction in trees given heavy pruning (−0.030 mm day−1) than in control trees (−0.019 mm day−1) and in trees given light pruning (−0.017 mm day−1). In September, when stems rapidly rehydrate, this trend was just the opposite being the growth greater in trees given heavy pruning (0.025 mm day−1) than in those given light pruning (0.017 mm day−1) or control trees (0.014 mm day−1). The differences in growth rates of the three groups were greatest during October, when heavy pruning was associated with the greatest growth rate. These differences were maintained in November, although they were not as pronounced.

Least squares mean estimates of daily girth increment (mm day−1 ± standard error) in the CA plot during different months for trees given different pruning treatments. Different letters indicate significant differences between treatments in a month (p < 0.05)
In the SB plot, the differences between treatments throughout the year were less evident (Fig. 2). In this plot, trees given heavy pruning generally had the slowest growth, and trees given moderate pruning had the greatest growth, although there were only significant differences in February, March, April, and September.

Least squares mean estimates of daily girth increment (mm day−1 ± standard error) in the SB plot during different months for trees given different pruning treatments. Different letters indicate significant differences between treatments in a month (p < 0.05)
The lack of significance of the treatment × year effect, and of all interactions that include the treatment × year effect suggests that the effect of pruning on growth did not vary throughout the study period.
Discussion
Effect of soil treatments
Ploughing decreases competition from herbaceous plants and woody shrubs, improves water infiltration, and reduces short term compaction of the uppermost soil layer in heavy grazed areas (Coelho et al. 2004; Cubera and Moreno 2007a). PL enhances stem and height growth of holm oak samplings (Sánchez-Andrés et al. 2006), which are highly dependent on soil features to survival (Gómez-Aparicio et al. 2008). In our study plots, soil treatments did not influence stem growth in holm oak, suggesting that water infiltration and shrub competition were not limiting factors for growth of adult trees. Rolo and Moreno (2011) reported that holm oak and rock-rose compete intensely for water and nutrients. Rock-rose can reduce the long-term survival of trees in densely shrub encroached stands, because it affects the water and nutritional status of trees, and has negative effects on stomatal conductance and photosynthesis. Furthermore, Moreno et al. (2007) found that shrub encroachment of dehesas reduced growth of holm oak shoots and reduced soil moisture. In our study, the similarity of growth in the ploughed and control plots may be explained by the scarcely developed shrub layer in our control plots, because treatments were carried out in 2006, but every plot was ploughed previously. On the other hand, because of the low density of trees in dehesas, holm oak does not use all available water in the soil (Cubera and Moreno 2007b). In shrub encroached dehesas, this water could be partly used by shrubs, because holm oak roots are very sparse (Moreno et al. 2005). Therefore the low density of shrubs did not have a significant detrimental effect on holm oak growth in our plots. In addition, the existence of a low density shrub layer promotes tree regeneration in dehesas (Pulido and Díaz 2005), so PL cannot be recommended extensively in these open woodland forests.
Our other soil treatment (PFS) also had no significant effect on stem growth. Holm oak and other sclerophyllous species are adapted to low nutrient availability (Monk 1966), are highly efficient in nutrient use, and consequently have long-living tissues and low growth rates (Aerts 1995). Mayor and Rodà (1994) reported no positive effects of N and P fertilization on the stem growth of holm oak. These authors suggested that holm oak preferentially allocates increased soil nutrient resources to leaf and shoot production instead of stem growth. However, Moreno et al. (2007) reported that NPK fertilization did not improve the shoot growth of holm oak. On the other hand, trees cannot efficiently use added nutrients without sufficient irrigation or rainfall. Cartan-Son et al. (1992) reported that fertilization only had a positive effect on the stem growth of holm oak during years with significant rain. According to Landsberg (1986), drought can affect the uptake and translocation of nutrients and disrupt the response to fertilizer. Fertilization may even be detrimental in arid sites because it decreases water availability. Cubera and Moreno (2007a) found that fertilization reduced the availability of soil water in dehesas during spring and early summer. However, the analysis of the physiological status and water potential of fertilized holm oak trees in this plot indicated no significant differences between control and PFS plots (Carevic et al. 2010). This supports our finding that fertilization had no significant effect on stem growth. Fertilization is not widely used in forestry because it is very expensive, but its use to enhance pasture and crop production and quality in dehesas would not affect tree growth.
Sowing of yellow lupin also had no effect on tree growth. Cubera and Moreno (2007a) suggested that addition of N increases water use efficiency and photosynthesis of holm oak, but Rivest et al. (2011) found that the amount of N2 fixed by leguminous plants in dehesas is very limited, and insufficient to compensate for the lack of this nutrient caused by pasture consumption. Yellow lupin has a shallow root system (Bramley et al. 2009), and holm oak is low dependent on resources of the uppermost soil layers (Moreno et al. 2007). Thus, the addition of N by growth of leguminous plants did not affect tree growth. On the other hand, recent phytopathology studies (Serrano et al. 2010) reported that yellow lupin can act as a vector of Phytophthora cinnamomi, one of the main agents responsible for “oak decline”. This serious pathogen is widespread at the regional scale, so planting of yellow lupin may have detrimental effects on holm oak in certain sites.
The lack of significance of spatial covariance indicates that tree location did not influence stem growth, suggesting that there were not better or worse locations within the plot for growth. The lack of significance of treatment x month, treatment x year, and treatment x month x year interactions, indicated that the effect of soil treatments on stem growth did not vary throughout the study period. Contrary, Cartan-Son et al. (1992) found a delayed positive effect of fertilization on the stem growth of holm oak to in a year with significant rain. In our study, we have repeated the application of the treatments every 3 years, and the study period comprised dry and wet years. Hence, our results suggested that there were not delayed or accumulative effects of fertilization on the growth of holm oak in our study site. Regarding the PL treatment, the lack of significance of this treatment throughout the whole study period could be explained by the slow shrub encroachment of control plots. Ramírez and Díaz (2008) found low covers of rock-rose in abandoned dehesas with less of 10 years of no grazing.
Effect of pruning
Pruning did not have a significant overall effect on stem growth in our plots, but it did have some significant effects during certain months (Figs. 1, 2).
Our results indicated that heavy pruning slightly decreased stem growth during the spring growth period. Previous studies on Pinus ponderosa (Gyenge et al. 2010) and Acacia nilotica (Siddiqui et al. 2010) indicated that pruning had a negative effect on diameter growth. The removal of a significant part of the crown disrupts the balance of the roots and crown and provokes the reallocation of resources to rebuild the aboveground biomass (Cañellas et al. 2007). This probably decreases the resources that would otherwise be used for stem growth. Intense pruning can also decrease total photosynthesis (Balandier et al. 2000) and carbon fixation (Gyenge et al. 2010) of trees. Our observation of greater stem contractions in heavy pruned trees during the summer drought in CA plot is possible related to a greater response of pruned trees to water stress.
In addition, heavy pruning can interrupt the normal translocation of water and nutrients (Dagit and Downer 2002), exacerbating the effects of water stress (Jackson et al. 2000; Gyenge et al. 2009). From an ecophysiological standpoint, the foliar area/sapwood ratio of a tree depends on water availability, and pruning can imbalance the hydraulic system at the level of the whole plant (Gyenge et al. 2010). According to Dagit and Downer (2002), heavy pruning may also negatively affect the roots, because it may deprive them of nutrients normally synthesized in the leaves. Water recycling of heavily pruned trees then becomes less efficient, adversely affecting the feedback between roots and crown (Ringgenberg 2001). Trees in CA could be additionally stressed by shallow and poorly developed soils, and even by an overall influence of oak decline (Brasier 1995), increasing therefore the sensitivity of pruned trees, despites the trees included in the study did not show visible symptoms during the study period.
Holm oak rehydrate quickly after the first rain events following the summer drought, provoking water stem expansions followed by true stem growth (Campelo et al. 2007; Gutiérrez et al. 2011; Martín et al. 2014). Hence, the greater stem increments of moderately pruned and heavily pruned trees in the CA plot in autumn after the marked summer stem contractions (Fig. 1) could be explained by the greater sensitivity of pruned trees to climate (Balandier et al. 2000; Gyenge et al. 2010), leading to changes in intra-annual growth patterns, with lower temporal autocorrelation among months. On the contrary, in the SB plot, where soils were more developed and oak decline was no detected, no significant stem contractions occurred during the summer and there were no significant differences between pruned and control trees during the autumn (Fig. 2).
Tree growth following moderate pruning was slightly greater during spring at the SB plot (Fig. 2), indicating that moderate pruning had no negative effect on growth in an area with better soils and good sanitary status (Pinkard and Beadle 1998). Growth of lightly pruned trees at the same plot was not significantly different from unpruned (control) trees, indicating that removal of small, suppressed, or dead branches did not affect the photosynthetic capacity or hydraulic balance of these trees (Gyenge et al. 2010).
The effects of the pruning treatments remained throughout the study period, as showed the lack of significance of the treatment × year effect, and of all interactions that include the treatment × year effect. According to Pinkard and Beadle (1998), the effects of pruning on growth are related to the growth rates of the species. The slower the growth rate, the longer will be the period to crown restoration following pruning. Holm oak is a slow-growing species and this could explain the long-lasting effects of the pruning treatments, despite they were applied only once before the beginning of the study.
Conclusions
Soil treatments had no effect on stem growth of holm oak. These treatments mainly alter the upper layer of soil, where holm oak roots are sparse. Fertilization and atmospheric nitrogen fixation by yellow lupin also had no significant effect on stem growth. Thus, its use to enhance pasture and crop production and quality would not affect tree growth.
Pruning affected the stem growth of holm oak during some months (February, March, April, July, September, October, and November), but depending on plot features. Heavy pruning slightly reduced growth during the spring, and moderate and heavy pruning increased the normal stem contractions (due to water stress during drought) and expansions (due to rehydration after drought) in sites with poorly developed soils or other stress causes. Hence, heavy pruning could affect the vigor and vegetative status of trees in areas where tree survival is already compromised. Light pruning did not affect tree growth, so this treatment may be acceptable if the extraction of fuelwood or biomass is one of the management objectives.
References
Aerts R (1995) The advantages of being evergreen. Trends Ecol Evol 10:402–407
Akaike H (1974) A new look at the statistical model identification. IEEE Trans Autom Control 19(6):716–723
Alejano R, Tapias R, Fernández M, Torres E, Alaejos J, Domingo J (2008) Influence of pruning and the climatic conditions on acorn production in Holm oak (Quercus ilex L.) dehesas in SW Spain. Ann For Sci 65(2):209
Alejano R, Vázquez-Piqué J, Carevic F, Fernández M (2011) Do ecological and silvicultural factors influence acorn mass in Holm Oak (southwestern Spain)? Agrofor Syst 83:25–39
Balandier P, Lacointe A, Le Roux X, Sinoquet H, Cruiziat P, Le Dizès S (2000) “SIMWAL”: a structure—function model simulating single walnut tree growth according to climate and pruning. Ann For Sci 57:571–585
Bramley H, Turner NC, Turner DW, Tyerman SD (2009) Roles of morphology, anatomy, and aquaporins in determining contrasting hydraulic behavior of roots. Plant Physiol 150:348–364
Brasier CM (1995) Phytophthora cinnamomi and oak decline in southern Europe. Environmental constraints including climate change. Ann For Sci 53:347–358
Bravo JA, Roig S, Serrada R (2008) Selvicultura en montes bajos y medios de Quercus ilex L., Quercus pyrenaica Willd. y Quercus faginea Lam. In: Serrada R, Montero G, Reque JA (eds) Compendio de Selvicultura aplicada en España. INIA, Madrid, pp 657–744
Campelo F, Gutiérrez E, Ribas M, Nabais C, Freitas H (2007) Relationships between climate and double rings in Quercus ilex from northeast Spain. Can J For Res 37:1915–1923
Campelo F, Nabais C, García-González I, Cherubini P, Gutiérrez E, Freitas H (2009) Dendrochronology of Quercus ilex L. and its potential use for climate reconstruction in the Mediterranean region. Can J For Res 39:2486–2493
Cañellas I, Roig S, Poblaciones MJ, Gea-Izquierdo G, Olea L (2007) An approach to acorn production in Iberian dehesas. Agrofor Syst 70:3–9
Carevic FS, Fernández M, Alejano R, Vázquez-Piqué J, Tapias R, Corral E, Domingo J (2010) Plant water relations and edapho-climatic conditions affecting acorn production in a holm oak (Quercus ilex L. ssp. ballota) open woodland. Agrofor Syst 78:299–308
Cartan-Son M, Floret C, Galan MJ, Grandjanny M, Le Floc’h E, Maistre M, Perret P, Romane F (1992) Factors affecting radial growth of Quercus ilex L. in a coppice stand in southern France. Vegetatio 99–100:61–68
Coelho COA, Ferreira AJD, Laouina A, Hamza A, Chaker M, Naafa R, Regaya K, Boulet AK, Keizer JJ, Carvalho TMM (2004) Changes in land use and land management practices affecting land degradation within forest and grazing ecosystems in the Western Mediterranean. In: Schnabel S, Ferreira A (eds) Advances in GeoEcology 37: Sustainability of Agrosilvopastoral Systems—Dehesas, Montados. Catena Verlag, Reiskirchen, pp 137–153
Cubera E, Moreno G (2007a) Effect of land-use on soil water dynamics in dehesas of Central-Western Spain. Catena 71:298–308
Cubera E, Moreno G (2007b) Effect of single Quercus ilex trees upon spatial and seasonal changes in soil water content in dehesas of Central Western Spain. Ann For Sci 64:355–364
Dagit R, Downer J (2002) To Prune or Not to Prune: Responses of Coast Live Oaks (Quercus agrifolia) to Canopy Retention during Transplanting. USDA Forest Service. http://www.californiaoaks.org/ExtAssets/RosiDagitTransplantingOaks.pdf. Accessed 20 August 2011
Eichhorn MP, Paris P, Herzog F, Incoll LD, Liagre F, Mantzanas K, Mayus M, Moreno G, Papanastasis VP, Pilbeam DJ, Pisanelli A, Dupraz C (2006) Silvoarable systems in Europe—past, present and future prospects. Agrofor Syst 67:29–50
Fritts HC (1976) Tree rings and climate. Academic Press, London
Gallardo A, Covelo F, Morillas L, Delgado M (2009) Ciclos de nutrientes y procesos edáficos en los ecosistemas terrestres: especificidades del caso mediterráneo y sus implicaciones para las relaciones suelo-planta. Ecosistemas 18:4–19
Gea-Izquierdo G, Cherubini P, Cañellas I (2011) Tree-rings reflect the impact of climate change on Quercus ilex L. along a temperature gradient in Spain over the last 100 years. For Ecol Manage 262:1807–1816
Gómez-Aparicio L, Pérez-Ramos IM, Mendoza I, Quero JL, Matías L, Castro J, Zamora R, Marañón T (2008) Oak seedling survival and growth along resource gradients in Mediterranean forests: implications for regeneration under current and future environmental scenarios. Oikos 117:1683–1699
Gutiérrez E, Campelo F, Camarero JJ, Ribas M, Muntán E, Nabais C, Freitas H (2011) Climate controls act at different scales on the seasonal pattern of Quercus ilex L. stem radial increments in NE Spain. Trees 25:637–646
Gyenge JE, Fernández ME, Schlichter TM (2009) Effect of pruning on branch production and water relations in widely spaced ponderosa pines. Agrofor Syst 77:223–235
Gyenge JE, Fernández ME, Schlichter TM (2010) Effect of stand density and pruning on growth of ponderosa pines in NW Patagonia, Argentina. Agrofor Syst 78:233–241
IPCC (2007) Climate change 2007: synthesis report. In: Core Writing Team, Pachauri RK, Reisinger A (eds) Contribution of working groups I, II and III to the fourth assessment report of the intergovernmental panel on climate Change. IPCC, Geneva, pp. 104
IUSS Working Group WRB (2007) World reference base for soil resources 2006, first update 2007, world soil resources reports no. 103. FAO, Rome
Jackson NA, Wallace JS, Ong CK (2000) Tree pruning as a means of controlling water use in an agroforestry system in Kenia. For Ecol Manag 126:133–148
Keeland BD, Young PJ (2014) Installation of traditional dendrometer bands. U.S. Geological Survey. National Wetlands Research Center. http://www.nwrc.usgs.gov/topics/Dendrometer/. Accessed 23 January 2014
Landsberg JJ (1986) Physiological ecology of forest production. Academic Press, New York, p 198
Littell RC, Milliken GA, Stroup WW, Wolfinger RD, Schabenberger O (2006) SAS system for mixed models. SAS Institute, Cary
Markewitz D, Figueiredo RD, de Carvalho CJR, Davidson EA (2012) Soil and tree response to P fertilization in a secondary tropical forest supported by an Oxisol. Biol Fertil Soils 48:665–678
Martín D, Vázquez-Piqué J, Fernández M, Alejano R (2014) Effect of ecological factors on intra-annual stem girth increment of holm oak. Trees 28:1367–1381
Mayor X, Rodà F (1994) Effects of irrigation and fertilization on stem diameter growth in a Mediterranean holm oak forest. For Ecol Manag 68:119–126
McMaster GS, Jow WM, Kummerow J (1982) Response of Adenostoma fasciculatum and Ceanothus greggii chaparral to nutrient additions. J Ecol 70:745–756
Monk CD (1966) An ecological significance of evergreenness. Ecology 47:504–505
Moreno G, Obrador JJ, Cubera E, Dupraz C (2005) Fine root distribution in dehesas of Central-Western Spain. Plant Soil 277:153–162
Moreno G, Obrador JJ, García E, Cubera E, Montero MJ, Pulido FJ, Dupraz C (2007) Driving competitive and facilitative interactions in oak dehesas with management practices. Agrofor Syst 70:25–40
Natalini F, Alejano R, Vázquez-Piqué FJ, Cañellas, I, Gea-Izquierdo G (2013) Growth trends and sensitivity to climate of declining Mediterranean open woodlands exhibiting widespread mortality in Southern Spain. Second American Dendrochronology Conference Ameridendron. Tucson, Arizona
Navarro R (2011) Situación actual de las dehesas. In: Alejano Monge R, Domingo Santos JM, Fernández, M (eds) Manual para la Gestión Sostenible de las dehesas andaluzas, Foro Encinal - Universidad de Huelva. Huelva, España, pp 62-68
Patterson HD, Thompson R (1971) Recovery of inter-block information when block sizes are unequal. Biometrika 58:545–554
Pinkard EA, Beadle CL (1998) Effects of green pruning on growth and stem shape of Eucalyptus nitens Deane and Maiden (Maiden). New Forest 15:107–126
Pulido FJ, Díaz M (2005) Regeneration of a Mediterranean oak: a whole cycle approach. EcoScience 12:92–102
Quézel P, Médail F (2003) Ecologie et Biogéographie des Forêts du Bassin Méditerranéen. Elsevier, Paris
Ramírez JA, Díaz M (2008) The role of temporal shrub encroachment for the maintenance of Spanish holm oak Quercus ilex dehesas. For Ecol Manag 255:1976–1983
Ringgenberg S (2001) Flow in the phloem. Arborist News 10(1):21–26
Rivest D, Rolo V, López-Díaz ML, Moreno G (2011) Belowground competition for nutrients in shrub-encroached Mediterranean dehesas. Nutr Cycl Agroecosyst 90:347–354
Rolo V, Moreno G (2011) Shrub species affect distinctively the functioning of scattered Quercus ilex trees in Mediterranean open woodlands. For Ecol Manag 261:1750–1759
San Miguel A (1994) La dehesa española: origen, tipología, características y gestión. Fundación Conde del Valle de Salazar, Madrid
Sánchez-Andrés R, Sánchez-Carrillo S, Benítez M, Sánchez-López A (2006) Tillage induced differential morphometric responses and growth patterns in afforestation with Quercus ilex. Soil Tillage Res 90:50–62
Scowcroft PG, Friday JB, Idol T, Dudley N, Haraguchi J, Meason D (2007) Growth response of Acacia koa trees to thinning, grass control, and phosphorus fertilization in a secondary forest in Hawai‘i. For Ecol Manage 239:69–80
Searle SR (1971) Linear Models. John Wiley & Sons, New York
Serrano MS, Fernández P, De Vita P, Carbonero MD, Trapero A, Sánchez ME (2010) Lupinus luteus, a new host of Phytophthora cinnamomi in Spanish oak-rangelands ecosystems. Eur J Plant Pathol 128:149–152
Siddiqui T, Farrakh-Nawaz M, Ahmed I (2010) Effect of different pruning intensities on the growth of Acacia nilotica (Kikar). Agrociencia 44(1):93–97
Acknowledgments
This study was sponsored by the Department of Innovation, Science and Business of the Regional Government of Andalusia, Spain (ref: C03-192) and by MEC-INIA, Spain (ref: SUM2006-00026-00-00).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Martín, D., Vázquez-Piqué, J. & Alejano, R. Effect of pruning and soil treatments on stem growth of holm oak in open woodland forests. Agroforest Syst 89, 599–609 (2015). https://doi.org/10.1007/s10457-015-9794-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-015-9794-x