Calcareous algae and microproblematica have been studied in the Middle Permian part of the Dalan Formation in Dena Mountain, Zagros, southwest Iran. Murgabian (=Wordian) microfossils are the most diversified. The assemblages encompass, among the dasycladales, Anthracoporella spectabilis, Epimastopora piae, Epimastoporella japonica, E. likana, Paraepimastopora? cf. densipora n. comb., Gyroporella cf. nipponica, G.? aff. fluegeli n. comb., G.? aff. symmetrica, G.? aff. digitula n. comb., Mizzia yabei; among the gymnocodiaceans, Gymnocodium bellerophontis, G. nodosum, Permocalculus cf. digitatus, P. forcepinus, P. cf. fragilis, P. plumosus, P. cf. solidus, Tauridium? sp.; among the “phylloid algae” Eugonophyllum? sp.; among the other incertae sedis algae Stacheoides sp., Ungdarella uralica, Fourstonella (Efluegelia) johnsoni, and among some microproblematica alternatively assigned to foraminifers or algae Aeolisaccus dunningtoni, Tubiphytes obscurus, Pseudovermiporella nipponica, P. sodalica, and P.longipora. Each taxon is briefly characterized, whereas a more detailed analysis of the epimastoporacean algae is given. Among them, the genera Epimastopora and Epimastoporella are emended. All the described microfossils have a relatively broad stratigraphic distribution in the Permian period but they are paleoecologically important. They are indeed confident micropaleontological Middle Permian proxies, particularly for shallow-marine, warm, well-oxygenated and relatively high-energy environments. They were affected by the end-Guadalupian crisis because they strongly decrease in the upper carbonate unit of the Dalan Formation and are not found in the Triassic sediments of the area.
Middle Permian algae are generally very productive, relatively well known, and cosmopolitan. The most abundant algae are the Gymnocodiaceae Permocalculus Elliott, 1955 and Gymnocodium Pia, 1920, and the dasycladale Mizzia Schubert, 1909. Some Early Permian forms persist such as Epimastopora (see below for the exact designation of the authors of this taxon) and Gyroporella Gümbel, 1872. Some other microorganisms, occasionally assigned to algae, are also rather common: Aeolisaccus Elliott, 1958, Tubiphytes Maslov, 1956b, Pseudovermiporella Elliott, 1958, as well as rare algosponges (i.e., incertae sedis algae), especially Ungdarella Maslov, 1956a.
In the literature, other Permian calcareous algae, such as the codiaceans, are poorly known. The codiaceans appeared in the Ordovician and were numerous and diverse in back-reef environments during the Late Silurian and Early to Middle Devonian (Rothpletz 1908; Poncet 1982, 1990; Mamet and Préat 1994; Vachard 1994) but unquestionable representatives are very rare during Carboniferous and Permian times (e.g., Vachard et al. 1989b, 2001). Moreover, some forms assigned to the Permian codiaceans belong in fact to the Gymnocodiaceae; a confusion due to the resemblance, after diagenesis, of the undivided sites of the laterals and siphons of the former with the files of cells of the second group. That is especially obvious in Succodium Konishi, 1955, Aphroditicodium Elliott, 1970, Tauridium Güvenç, 1966, and Thaiporella Endo, 1969 which, although described as codiaceans, rather belong to gymnocodiaceans. Although attributed to the red algae by the genial paleophycologist Julius von Pia in 1937 (principal arguments being, the presence of cellular files and conceptacles), the gymnocodiaceans were recently transferred to the green algae by some authors such as Mu Xinan and Riding (1983), Chuvashov et al. (1987), Bucur (1994), Radoičić (2004), and Schlagintweit and Sanders (2007). We continue to favor an assignment to the galaxauracean red algae (shape and subcortical location of the conceptacles, long files of cells and not unicellular siphons, etc.). Inversely, it seems possible that some so-called Paleozoic “codiaceans” belong, in fact, to red ancestral gymnocodiaceans. Nevertheless, the unique Devonian species formally assigned to the group, “Gymnocodium” devonicum Mamet et al., 1993, is most probably a fossil ulotrichale (according to the criteria proposed by Kozlowski and Kazmierczak 1968). However, these discussions about the botanical assignment of gymnocodiacean at least document the difficulty to distinguish various fossil green and red algae (for other examples see Vachard and Cózar 2010).
Compared with Mizzia and Gyroporella, the other Permian genera of dasycladales are generally rare and endemic. There are Atractyliopsis Pia, 1937, Anthracoporella Pia, 1920, Clavaporella Kochansky-Devidé and Herak, 1960 emend. Vachard, 1980, Eoclypeina Emberger in Vachard, 1985, Goniolinopsis Milanović, 1966b, Imperiellla Elliott and Süssli, 1975, Kochanskyella Milanović, 1974, Likanella Milanović, 1966a (non-1965); Nanjinoporella Mu and Elliott, 1982, Pseudotabasoporella Rashidi and Senowbari-Daryan, 2010, Salopekiella Milanović, 1965a (non-1965b), Sinoporella Yabe, 1949, Uragiellopsis Vachard in Vachard and Montenat 1981, Tabasoporella Rashidi and Senowbari-Daryan, 2010, and Velebitella Kochansky-Devidé, 1964. The Lopingian (i.e., Late Permian) algal assemblages seem to be restricted to Mizzia, Permocalculus, and Gymnocodium, except in Armenia (formerly Transcaucasia) with the very poorly known genera Johnsonia Kordé, 1965 and Endoina, Kordé 1965; Greece with Clavaporella and Macroporella Pia, 1912 (Vachard et al. 1993a, b, 1995, 2003), or in northern Italy with Atractyliopsis lastensis Accordi, 1956, “Physoporella”, and “Macroporella” (Accordi 1956; Praturlon 1963; Noé 1987).
The gymnocodiacean alga Gymnocodium has its first acme during the Midian (with G. exile Mu, 1981, G. bellerophontis (Rothpletz, 1894) Pia, 1920, and G. nodosum Ogilvie-Gordon, 1927), whereas Permocalculus is abundant since the Kubergandian (Vachard, 1980) or perhaps as soon the Artinskian or even the Sakmarian (Vachard and Moix, unpublished data; Krainer, Lucas and Vachard, unpublished data).
“Phylloid algae” are relatively rare in our material. They constitute, in fact, a composite group, the representatives of which have been assigned to the codiaceans, ancestral corallinales, and algospongia (Vachard et al. 1989a, b, 2001; Vachard and Cózar 2010; Granier 2012) or considered as a result of complex biological interactions (Vachard et al. 1989a, b, 2001). For example, the Eugonophyllum illustrated by Forsythe et al. (2002) or specimens of Ivanovia illustrated by Torres (2002) are comparable with recrystallized, peripherically microbored blades of an ancestral corallinale, the red alga Archaeolithophyllum Johnson, 1956. This transition between Archaeolithophyllum and “phylloid algae” is peculiarly conspicuous in the Midian deposits of Tunisia (Vachard et al. 1989a).
Within the reefal ecosystems, the Cisuralian (early Early Permian) reefs with the giant calcareous red alga Palaeoaplysina Krotov 1988, well known in Arctic regions of Canada, Russia, and Norway (Morin et al. 1994; Hüneke et al. 2001; Weidlich 2002a, b; Reid et al. 2007; Anderson and Beauchamp 2010; Blomeier et al. 2011) disappear and are replaced in the Middle Permian by the CAC: calcisponge-algal-cement reefs of Flügel et al. (1984) (see also Weidlich 2002a, b). The transitional period, Artinskian and Kungurian, is generally poorly known (see Flügel and Flügel-Kahler 1980 in the Trogkofelkalk of the Carnic Alps) but it seems accepted that the end-Early Permian reef crisis is due to a general cooling (due to a glaciation, increasing paleobathymetry, change from tropical to temperate climate, upwelling, or “Auernig paradox”) or a general rise of the sea level related to the end of the LPIA (great Gondwanan glaciation) (see James 1997; Samankassou 1999, 2002; Soreghan et al. 2008) with a generalized change of photozoan assemblages to heterozoan ones.
A special, or at least especially developed during the Permian, micro-ecosystem is constituted by “algal biscuits”; large oncoids or complex biopisoliths with cyanobacteria, Claracrusta Vachard and Montenat, 1981, Ellesmerella Mamet and Roux in Mamet et al., 1987 (e.g., Henbest 1963; Vachard 1980; Flügel and Flügel-Kahler 1980; Roux 1985; Toomey et al. 1988; Sanders and Krainer 2005; Gaetani et al. 2009). Some incorporated “oncoid-dwelling” microfossils in the sense of Schlagintweit and Gawlick (2009a) are represented by the algosponge Fourstonella (Efluegelia) and the rare dwarf foraminifer Tetrataxis.
Other peculiar Permian constructions are those of Tubiphytes (non Shamovella Rauzer-Chernousova, 1950), a microfossil generally considered as an alga (Razgallah and Vachard 1991; Riding and Barkham 1999; Fagerstrom and Weidlich 2005) but also of a hyperspecialized foraminifer as well as of Pseudovermiporella (Vachard and Krainer 2001a, b).
Permian calcareous algae and problematic microfossils were described from numerous Tethyan localities (Table 1). For additional references concerning the genera and species of Upper Paleozoic dasycladales, see the compilations of Pia (1920), Kochansky-Devidé and Gusic (1971), Emberger (1976), Bassoullet et al. (1979), Vachard (1980, 1985), Roux (1985), Deloffre (1988), Riding and Guo (1992), Deloffre and Granier (1991), Granier and Deloffre (1995), Granier and Grgasović (2000), Pille (2008), Pille et al. (2010), Cózar et al. (2009), and Mamet and Préat (2010). Calcareous algae and problematic microfossils, including dasycladaleans and other algal groups, have not been well studied in Iran, particularly in the Permian of Zagros. Golesstaneh (1979) reported the occurrence of 15 algal genera and numerous species of Permian dasycladales from the Zagros Mountains, southeast Iran, without, however, giving a detailed description and illustration.
Table 1 List of calcareous algae and problematic microfossils from Tethyan localities and from some selected localities of the Americas
In central, southwestern, and northern Iran, several authors have reported, in the last 50 years, previously known algal genera (e.g., Flügel 1963; Kalantari 1986; Vaziri et al. 2005; Gaillot and Vachard 2007; Kolodka et al. 2012). New algal taxa were introduced by Elliott and Süssli (1975) and Rashidi and Senowbari-Daryan (2010), and Senowbari-Daryan and Rashidi (2010, 2011). Elliott and Süssli (1975) described the genus Imperiella, with the type species I. iranica, from the Permian Ruteh limestone of the Alborz Mountains in northern Iran. Senowbari-Daryan and Rashidi (2010, 2011) reported dasycladales and the codiacean genera Anchicodium Johnson, 1946 and Iranicodium Senowbari-Daryan and Rashidi, 2010, as well as Lercaritubus problematicus Flügel et al. 1990 and Vangia telleri (Flügel in Flügel et al. 1984), two problematic organisms from the Permian Jamal Formation of Shotori Mountains in Central Iran (Senowbari-Daryan and Rashidi 2011). Nevertheless, Iranocodium is very difficult to characterize, and Vangia is probably a junior synonym of Bacinella Radoičić, 1959.
The most important environments and microfossils from a paleobathymetrical point of view are described more accurately in the following paragraphs. Overall, more than 30 genera of cyanobacteria, true algae (dasycladales and rhodophycophyta), incertae sedis algae or protista (algospongia), and microproblematical species were found in the Permian deposits of Dena Mountain.
Geological setting
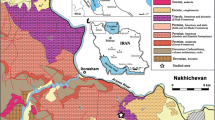
The Iranian plateau extends over a number of continental fragments welded together along suture zones of oceanic character. The fragments are delineated by major boundary faults, which appear to be inherited from older geological periods. Each fragment differs in its sedimentary succession, nature, and age of magmatism and metamorphism, and its structural character and intensity of deformation (Berberian and King 1981; Heydari 2008). These fragments are exposed in the following structural provinces: (1) Zagros; (2) Sanandaj-Sirjan; (3) Urumieh-Dokhtar; (4) Central Iran; (5) Alborz; (6) Kopet Dagh; (7) Lut; and (8) Makran (Fig. 1).
Fig. 1
General map of Iran showing the eight geologic provinces. Dena Mountain is located in the Zagros province (adopted from Heydari et al. 2003)
The Late Permian to Early Triassic periods were characterized in the Zagros Mountains by marine carbonate sedimentation (Setudehnia 1978; Koop and Stoneley 1982). These deposits, known as the Dalan and Kangan Formations, correspond to the Khuff Formation in Saudi Arabia, Kuwait, UAE, and Qatar (Szabo and Kheradpir 1978; Vaslet et al. 2005; Hughes 2005).
The Permian Dalan and Khuff Formations contain extensive gas reservoirs in the Greater Persian Gulf area (Kashfi 1992). The Dalan Formation corps out in some mountains at the base of Zagros zone in southwestern Iran such as Zard-Kuh (Setudehnia 1976), Dena (Setudehnia 1976; Baghbani 1997; Insalaco et al. 2006; Gaillot and Vachard 2007), Gakhum (Insalaco et al. 2006; Kolodka et al. 2012), and Oshtoran-Kuh and Surmeh (Sharland et al. 2001; Gaillot and Vachard 2007).
In 1976, the Stratigraphic Committee of Iran adopted a new term for the Permian succession in southern Iran, the Deh Ram Group, which was named after a village on the southwestern flank of the Kuh-e Surmeh in southern Iran.
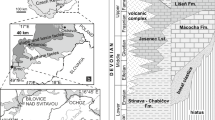
Subsequently, the Deh Ram Group was divided into three formations: the Lower Permian Faraghan Formation, the Upper Permian Dalan Formation, and the Lower Triassic Kangan Formation (Kashfi 2000) (Fig. 2). The Dalan Formation is equivalent to (a) the Jamal Formation (Stöcklin et al. 1965; Partoazar 1995; Aghanabati 2004) in the Shotori Range of the Tabas area (eastern Iran); (b) the Surmaq, Abadeh, and Hambast Formations (Taraz 1969, 1974; Iranian-Japanese Research Group 1981; Partoazar 1995, 2002; Baghbani 1996; Kobayashi and Ishii 2003) in the Abadeh region (Central Iran); (c) the Ali Bashi and Jolfa Formations (Partoazar 1995; Baghbani 1996) in the Jolfa region (northwestern Iran), and (d) the Nesen and Ruteh Formations (Assereto 1963; Glaus 1964; Vachard et al. 1993a, b, c) in the Alborz Range (northern Iran) (Fig. 3). The Dalan Formation is conformably underlain by the Faraghan Formation and disconformably overlain by the thin-bedded limestone of the Kangan Formation (Szabo and Kheradpir 1978; Gaillot 2006; Gaillot and Vachard 2007).
Fig. 2
Permian–Triassic stratigraphy of the Zagros basin, slightly modified from James and Wynd (1965)
The objective of our research has been the study of the Dalan Formation in a section located 58 km northwest of Yasouj city (the center of the Kohgilouye and Boyerahmad province) and approximately 7 km northwest of Sisakht town, at the southern flank of the Kuh-e Dena hill (coordinates 30°55′52″ N, 51°25′24″ E; Fig. 4). The Dalan Formation in this section, the Dena section, is 325.7 m thick and can be divided into three rock units: Lower Dalan Member, Nar Member, and Upper Dalan Member. The Lower Dalan Member is Murgabian to late Midian in age (Szabo and Kheradpir 1978; Johnson 1981; Gaillot 2006; Gaillot and Vachard 2007); the Nar Member is dated as latest Midian (Baghbani 1988; Vachard et al. 2002; Insalaco et al. 2006; Gaillot 2006); and the Upper Dalan Member is Djulfian to Dorashamian in age (Insalaco et al. 2006; Gaillot 2006; Gaillot and Vachard 2007). The Lower Dalan Member yields rare trilobites, sponges, corals, gastropods, brachiopods, bryozoans (Ernst et al. 2011), and various foraminifers (Baghbani 1997; Insalaco et al. 2006; Gaillot 2006; Gaillot and Vachard 2007), calcareous algae, and microproblematica. In Dena Mountain, the evaporites of the Nar unit are replaced by thick beds of sandstone (Aghanabati 2004), and the age of the Dalan Formation is Murgabian (=Wordian) to early Djulfian (=Wuchiapingian). As the Upper Dalan Member has been accurately described by Gaillot (2006), the aim of this paper is to describe the microproblematica and algae of the Lower Dalan Member (Fig. 5).
Stratigraphic column with distribution and abundance of Dasycladales, red algae, and problematic microfossils of the studied Dalan Formation in Dena Mountain
Abbreviations: L = length; D = outer diameter; d = inner diameter; s = thickness of wall; p = diameter of pores (= diameter of laterals); ip = interval between two pores (i.e., two laterals).
1920Anthracoporella spectabilis n. gen. n. sp.—Pia: 15, text-fig. 3, pl. 1, figs. 7–11
1968aAnthracoporella spectabilis Pia—Elliott: 21, pl. 2, figs. 1–2
2000Anthracoporella spectabilis Pia—Granier and Grgasović: 11 (cum syn.)
2001Anthracoporella spectabilis Pia—Vachard et al.: 385, 387, figs. 12.1–12.10
Description
All the specimens observed in the lower carbonate unit of the Dena section have the form of straight and long fragments. They can be differentiated from epimastoporacean fragments by the acrophore or slightly phloiophore, aspondyl, more or less parallel, simple or rarely bifurcated laterals.
Dimensions
L = 200–2.26 mm; s = 0.40–0.60 mm; p = 0.03–0.04 mm; ip = 0.04–0.06 mm.
Epimastopora alpina Kochansky-Devidé and Herak, 1960 (=Epimastopora sp. sensu Pia 1937, pl. 97, fig. 4).
Discussion
The genus Epimastopora of Pia (1922) and Pia (1937) does not follow the rules for creating a taxon as no type species has been defined. Moreover, both names of Pia are not really identical because the first one (1922) refers informally to a species “?Gyroporella n. f.” sensu Gortani (1906) that will become an Epimastoporella (see below), whereas the second one (1937) corresponds more to the first valid description of the genus with the description of E. alpina. Before the typification of this latter species by Kochansky-Devidé and Herak (1960), Johnson (1946), Kordé (1951), and Endo (1959) had created successively some species of epimastoporaceans. Nevertheless, all are very remote from the type area (the Austrian–Italian border) and therefore do not correspond to the intentions of Pia (1922 or 1937). Furthermore, the taxon of Kordé was renamed Globuliferoporella Chuvashov, 1974 and that of Johnson Paraepimastopora Roux, 1979. Although the material of Gortani (1906) has been revised and re-described as Epimastopora alpha and E. beta, respectively by Elliott (1956), it is difficult to attribute the authorship of Epimastopora to this author because he did not understand the nature of this taxon very well. Indeed, (a) Epimastopora “alpha” Elliott 1956 might be a Globuliferoporella (also present among the unpublished topotypes of Vachard and Argyriadis and those of Krainer); (b) E. “beta” Elliott 1956 is an Epimastoporella according to Roux (1979) and Granier and Grgasović (2000), (c) E. minima Elliott, 1956, which is evidently a gymnocodiacean and not a dasycladale (Elliott 1968a; Granier and Grgasović 2000; Vachard et al. 2012), and (d) E. malaysiana Elliott, 1968b resembles many things but no an epimastoporean. Furthermore, according to Granier (pers. comm. September 2012), the 1988 edition of the International Code of Botanical Nomenclature states in Article 23.6 that “The following are not to be regarded as specific epithets: (b) ordinal adjectives used for enumeration, for instance, alpha, beta, gamma, etc.”
Consequently, the nomenclatural proposals as “Epimastopora Pia ex Kordé 1951, emend. Roux 1979” or “Epimastopora (Pia 1922) Elliott, 1956” cannot be accepted. Hence, we suggest here the name Epimastopora (Pia, 1937) Kochansky-Devidé and Herak, 1960 emend. Vachard in Perret and Vachard, 1977 (see also Gaillot 2006; Vachard et al. 2012). Indeed, the emended diagnosis and the designation of Epimastopora alpina (=Epimastopora sp. sensu Pia 1937, pl. 97, fig. 4) as type by Kochansky-Devidé and Herak (1960) are most consistent with the intentions of Pia (1937). We will explain below why the name “Epimastopora” sensu Pia (1922) can be abandoned and replaced by Epimastoporella Roux, 1979.
Occurrence
As already indicated by Johnson (1963), the distribution of this cosmopolitan alga is Middle Pennsylvanian–Late Permian; the other species of the genus, recorded from the late Bashkirian to Jurassic, have been erroneously assigned to the genus as re-described here.
1960Epimastopora piae n. sp.—Bilgütay: 55–57, text-fig. 2a–c, pl. 2, figs. 1–4
2000Epimastopora piae Bilgütay—Granier and Grgasović: 57 (with synonymy list)
Description
A few specimens of this alga have been observed in the lower carbonate unit of Dena section. The thalli are broken in elongate fragments. The laterals are simple, cylindrical, acrophorous, parallel, and relatively wide.
Dimensions
L = 1.73–2.29 mm; s = 0.34–0.55 mm; p = 0.05–0.10 mm; ip = 0.01–0.03 mm.
Comparison
Our specimens have smaller interpores than the type of specimens from Turkey studied by Bilgütay (1960).
Occurrence
Early Permian of Turkey, Carnic Alps (Austria), Italy, Slovenia, Urals, Sumatra, Xizang/Tibet. Discovered in the Murgabian (Middle Permian) of the Dena section.
Epimastoporella differs from Epimastopora emend. herein by inflated laterals, either ellipsoidal or acrophorous at the two extremities with a vesiculifer median part.
Discussion
The invalid taxon “?Gyroporella n. f.” Gortani (1906) was initially included in “Epimastopora” Pia, 1922. Elliott (1956) described two species in the material of Gortani, Epimastopora alpha, and E. beta. Consequently, one of these species could appear as the type species of Epimastopora. Nevertheless, these species were subsequently neglected; probably because, as indicated by Granier and Grgasović (2000), these names are invalid with reference to article 23.6 of the ICBN (see above). Epimastopora japonica Endo, 1951 is generally regarded as a synonym of “?Gyroporella n. f.”. As E. japonica is also the type species of Epimastoporella, we suggest that (1) Epimastoporellajaponica is the first valid name of “?Gyroporella n. f.” Gortani (1906); (2) the name Epimastoporella Roux, 1979 emend. herein must replace Epimastopora (Pia 1922), a nomen nudum; (3) from now on, the name Epimastopora corresponds only to Epimastopora (Pia 1937) Kochansky-Devidé and Herak, 1960, emend. Vachard in Perret and Vachard, 1977, re-emend herein.
Occurrence
Late Pennsylvanian-Middle Permian. Epimastoporella emend. appears after and disappears before Epimastopora emend.; both genera are cosmopolitan.
1979Epimastoporella japonica (Endo) n. comb.—Roux: 809
1987Epimastoporella japonica (Endo) n. comb.—Mamet et al.: 34, pl. 15, figs. 6–12, pl. 6, figs. 1–3 (partim; the proposed synonyms of E. likana excluded)
1960Epimastopora likana n. sp.—Kochansky-Devidé and Herak: 78, pl. 4, figs. 5–10
2000Epimastoporella japonica (Endo) Roux—Granier and Grgasović: 62 (with synonymy list, 21 references erroneously assigned to E. japonica)
Description
The elongate fusiform shape of the laterals is characteristic and differs from those of E. japonica in contrast to the opinion of Mamet et al. (1987).
Dimensions
L = 2.12 mm; s = 0.27 mm; p = 0.075 mm; ip = 0.10 mm.
Occurrence
Late Pennsylvanian–Late Permian. Croatia, Slovenia, Montenegro, Carnic Alps Austria-Italy), Greece, Turkey, Urals (Russia), Iran (Azerbaijan, eastern Alborz), Oman, Afghanistan, New Mexico, Guatemala. Discovered in the Murgabian (Middle Permian) of the Dena section.
Genus Paraepimastopora Roux, 1979 emend. Krainer and Vachard, 2002.
Paraepimastopora is probably the most abundant dasycladale in the North American Carboniferous limestone (Johnson 1946; Roux 1989; Krainer et al. 2005; Gómez-Espinosa et al. 2008). This genus is well defined and easy to identify (Roux 1989; Krainer and Vachard 2002) due to (1) its wide axial cell; (2) its abundant, thin, parallel, acrophorous and aspondyl laterals; and (3) its oblique well-developed intusannulations. Nevertheless, previous numerous reciprocal misinterpretations between Paraepimastopora and Orthriosiphon Johnson and Konishi 1956 exist in the literature. For example, Orthriosiphon sp. of Maslov (1973, pl. 11, fig. 6) is a typical Paraepimastopora with a characteristic imperforate intusannulation (bottom, left).
Occurrence
The genus appeared first in the Tethys in the late Viséan (Sánchez-Chico et al. 1995; Cózar and Somerville 2004; Cózar et al. 2005; Pille 2008). It is then known in the early Serpukhovian of Austria (Krainer and Vachard 2002), ?Bashkirian of Bolivia (Mamet 1994), and Moscovian of Tunisia (Glintzboeckel and Rabaté 1964, pl. 12, fig. 2, pl. 16, figs. 1–2) and the Urals (Ivanova 2008, pl. 28, figs. 8–9; under the name Epimastopora grandis Chuvashov and Anfimov, 1988, correctly assigned to Paraepimastopora by Mamet and Préat 2010, p. 34–35). The genus appeared in North America during the Moscovian (Mamet et al. 1987) or perhaps as soon as the late Bashkirian (Groves 1984, text-fig. 7, p. 289). Its cosmopolitan acme occurred during the middle-late Virgilian/Gzhelian (Roux 1989; Krainer et al. 2005). It last appeared in the Early Permian (Sakmarian) (Mamet et al. 1987; Roux 1989; Mamet 1991; Vachard and Krainer 2001b; Vachard et al. 2012). The present study shows that it survived in the Murgabian of southern Iran.
Paraepimastopora? cf. densipora (Endo 1969) n. comb.
1969Epimastopora densipora n. sp.—Endo: 80, Pl. 42, figs. 1–2
cf. 2000Epimastopora densipora Endo—Granier and Grgasović: 55 (with synonymy)
Description
This alga is rare in the lower carbonate unit of Dena section. Its fragments are curved and their thickness remains the same across their length. The interval between pores is very small. The Dena profile specimen has larger pore diameters than the specimens studied by Endo (1969).
Dimensions
L = 2.50 mm; s = 0.38 mm; p = 0.05 mm; ip = 0.02 mm.
Occurrence
Middle Permian of Thailand, Japan, Slovenia, Iran (Tabas area). Discovered in the Murgabian (Middle Permian) of the Dena section.
Tribe Gyroporelleae Pal, 1976 emend. Bassoullet et al., 1979
Genus Gyroporella Gümbel, 1872 emend. Kochansk-Devidé, 1970b
Dasycladales and Gymnocodiaceae of the Dena section. Scale bars = 1 mm. a-c, d? e, g? Mizzia yabei; a oblique section; sample D41; b oblique section; Sample D79; c oblique section. Sample D36e; d cross section; sample D41; e longitudinal section; sample D36; g cross section; sample D47. fGyroporella aff. symmetrica; cross to oblique section; sample D74. h–jPermocalculusplumosus; h longitudinal section; sample D47; i longitudinal section; sample D43; j longitudinal section; sample D44
1952Gyroporella sp.—Konishi: 157, pl. 14, figs. 15–16
1955Gyroporellanipponica n. sp.—Endo and Hashimoto: 705, fig. 1
1959Gyroporella igoi n. sp.—Endo: 190, pl. 34, figs. 1–5, pl. 35, figs. 1–5, pl. 40, fig. 3, pl. 41, figs. 7–8
2000Gyroporella nipponica Endo and Hashimoto—Granier and Grgasović: 75 (with synonymy)
Description
Abundant longitudinal and oblique sections of the species have been found in the lower carbonate unit of the Dena section. This alga exhibits a particularly typical cylindrical thallus and vesiculiferous laterals. The ratio of outer to inner diameter is 0.75. The specimens from Dena Mountain are nearly similar to the specimen described and illustrated by Endo and Hashimoto (1955) from the Permian of Japan, but the Dena samples are larger.
Dimensions
D = 1.20–2.74 mm; d = 0.72–2.06 mm; s = 0.16–0.27 (0.83) mm; p = 0.11–0.16 (0.36) mm.
Occurrence
Late Pennsylvanian-Permian. Japan, Spain, Croatia, Montenegro, Carnic Alps, Urals, Turkey, Iran (Central Iran), South China, Thailand, SW Canada, NW USA. Murgabian (Middle Permian) of the Dena section.
aff. 1978Epimastopora fluegeli n. sp.—Kulik: 198, pl. 5, figs. 4–5
aff. 2000Epimastopora fluegeli Kulik—Granier and Grgasović: 56 (with synonymy)
Description
Rare oblique sections of the Faxon have been found in the lower carbonate unit of the Dena section. The alga is cylindrical with vesiculiferous laterals, i.e., it belongs more probably to Gyroporella or a similar genus (e.g., Gyroporella cf. prisca sensu Krainer et al. 2005, fig. 6.13). The distal extremities of these laterals are very inflated, and as the proximal acrophore extremity is not calcified the laterals appear almost spherical.
Dimension
D = 1.20–2.74 mm; s = 0.16–0.27 mm; p = 0.11–0.16 mm; ip = 0.025–0.05 mm.
Occurrence
Early Permian of Russia and Italy. Murgabian (Middle Permian) of the Dena section.
aff. 2000Epimastotopora symmetrica (Johnson) Roux—Granier and Grgasović: 63, (with synonymy)
aff. 2004Epimastopora symmetrica (Johnson) Roux—Mamet and Villa: 157
Description
A poor material of this alga has been observed in the lower carbonate unit of the Dena section. The alga is cylindrical or slightly undulating. The branches are closely arranged and the distance between pores is relatively narrow for a true Gyroporella. The specimen investigated in this study has larger pores and smaller interpores than the specimens studied by Johnson (1951). In fact, this taxon is very ambiguous. pl. 10, figs. 1–2, and 5 of Johnson probably corresponds to a Gyroporella, whereas pl. 10, fig. 3 is a Gymnocodium and pl. 8, fig. 7 and pl. 10, fig. 4 possibly are diploporaceans. In the past, the taxon has been assigned to Globuliferoporella and “Epimastopora” sensu Roux (1979). It is possibly also related to Gyroporella microporosa afghanica Vachard in Vachard et Montenat (1981), an unquestionable Gyroporella.
Dimensions
L = 2.38 mm; D = 0.60 mm; p = 0.19 mm; ip = 0.03–0.11 mm.
Occurrence
Permian of Texas, Turkey, Afghanistan, and Tunisia. Midian (Middle Permian) of the Dena section.
Gyroporella? aff. digitula (Chuvashov and Anfimov 1988) n. comb.
aff. 1988Epimastopora digitula n. sp.—Chuvashov and Anfimov: 62, pl. 25, figs. 4–5
Description
This taxon is very rare in our material and apparently unpublished. Only the Moscovian species Epimastopora digitula seems to have some resemblance by its wide, regularly parallel vesiculiferous laterals and the beginning of segmentation of the test, with a more or less triangular pseudo-annulation. Nevertheless, it is evident that this species is not an Epimastopora and corresponds more likely to a Gyroporella. More material might permit to describe it as a new taxon.
Dimensions
L = 2.83 mm; s = 0.20 mm; p = 0.07 mm; ip = 0.01/0.05 mm.
Occurrence
Type material described from the late Moscovian (Middle Pennsylvanian) of the Urals (Russia). Murgabian (Middle Permian) of the Dena section.
2005Mizzia velebitana Schubert—Hughes: pl. 1, figs. 21–23
Description
Numerous specimens of this species were found. M. yabei is apparently the only species of the genus present in the section. For example, neither M. velebitana Schubert 1909 nor M. cornuta Kochansky-Devidé and Herak, 1960 were truly identified, although some specimens resemble these species (Fig. 7d, g, respectively). M. yabei is characterized by club-shaped articles and the drum-stick-shaped laterals. M. velebitana differs by wider spherical articles, and M. cornuta by “horny” protuberances at the extremities of the laterals. These differences are clearly specific during all transitional stages, and cannot justify the creation of the genus Yabeites Chuvashov, 2001 for M. yabei and Cornutella Chuvashov, 2001 for M. cornuta. Furthermore, in our material, atypical specimens are occasionally similar to Mizzia bramkampi Rezak, 1959 and/or the genera Eogoniolina Endo, 1953 or Permoplexella Elliott, 1968a.
In general, in the Near and Middle East, the most common species of Mizzia seems to be M. yabei, whereas M. velebitana (well characterized by Pia 1920; Vachard 1980; and Flügel et al. 1984) is rare or absent. In particular, in our opinion Mizzia velebitana as illustrated by Elliott (1968a) and Hughes (2005) belong unquestionably to M. yabei.
The transverse sections are difficult to distinguish (Fig. 7a–d, g) but the axial sections are characteristic (Fig. 7e). Some specimens in our material exhibit relatively prominent lateral extremities (Fig. 7g). This feature is generally due to a weak, differential abrasion of the outer surface and should not be confused with M. cornuta.
Dimensions
D = 1.30–2.30 mm; d = 0.80–0.130 mm; s = 0.16–0.68 mm; p = 0.16–0.38 mm.
Occurrence
Permian of Japan, South China, Guatemala, southwestern USA, Thailand, Malaysia, Pakistan, Afghanistan, Iran (Alborz, Zagros; Bozorgnia 1973; Kolodka et al. 2012 (no 2006); this study), Saudi Arabia, Tunisia, Turkey, Greece, Carnic Alps, Hungary, Slovenia, Croatia, Montenegro. Murgabian and Midian (Middle Permian) of the Dena section.
This alga has been sporadically found in the lower and upper carbonate unit of the Dena section. The specimens are perfectly similar to an illustration of Partoazar (1995, pl. 5, fig. 1), but differ from the type material of Pia by a more regularly cylindrical thallus, which is weakly curved, with the files of cells smaller and less marked through the wall. In one specimen, the rounded conceptacle has a diameter of 0.096 mm. The specimens studied by Rezak (1959) have broader pore diameters than those of the Dena specimens. Conversely, the latter have larger pores and thicker walls than the specimens studied by Elliott (1955).
Dimensions
L = 1.22–1.76 mm; D = 1.14–1.18 mm; d = 0.56–0.86 mm; s = 0.33–0.40 mm; p = 0.05–0.09 mm.
Occurrence
Middle-Late Permian of Europe (Slovenia, Serbia, Bosnia, Montenegro, Italy, Austria, Greece, Hungary), Tunisia, Urals, Near and Middle East [Turkey, Caucasus, Saudi Arabia, Oman, Iraq, Armenia, and Iran (Lys et al. 1978; Partoazar 1995; Vaziri et al. 2005; Kolodka et al. 2012)], central and southwestern Asia (Pakistan (Salt Range), Afghanistan, Uzbekistan (Pamir), Tibet, South China, Thailand, Malaysia, Japan; New Zealand; ?Guatemala. Murgabian (Middle Permian) of the Dena section.
This species is larger than G. bellerophontis with marked thallus undulations. G.nodosum was mentioned but not illustrated by Jenny (1977, p. 159, 160) from the Nesen Formation of eastern Alborz. As corrected by Mu (1982), G. nodosum sensu Wang (1974) is a G. bellerophontis.
Dimensions
L = 2.96 mm; D = 1.48 mm; d = 0.96 mm; s = 0.60 mm; p = 0.05–0.09 mm.
Occurrence
Late Permian of Italy, Austria, Turkey, ?Tunisia, Saudi Arabia, Armenia, Iran, and Ladakh (Himalaya). Midian (Middle Permian) of the Dena section.
cf. 1992Gymnocodium bellerophontis Pia—Okla: pl. 46, figs. 3–5
Description
The alga is found in the lower and middle carbonate units of the Dena section. The thallus is cylindrical or finger-shaped. The medullar zone is always completely uncalcified, and the cellular files in the cortical zone are few abundant and resemble acrophorous laterals. These cortical files of cells are arranged slightly oblique to the wall and are occasionally divided into two or three branches with diameters of 0.052–0.068 mm. Their density in the cortex is moderate. Rare conceptacles were observed (Fig. 8k). Contrary to the opinion of Elliott (1955), this species cannot be the basal portion of a Permocalculus fragilis because both species are morphologically very different in our material.
Dimensions
L = 1.37–2.29 mm; D = (0.37)–1.35–1.40 mm; d = (0.18)–0.76–0.94 mm; s = 0.12–0.24 mm; p = 0.03 mm.
Occurrence
Murgabian (Middle Permian) of the Dena section. This alga has been found sporadically in thin-sections of the lower and middle units of the Dena section, associated with Permocalculus spp. and Gyroporella spp.
1951Gymnocodiumforcepinum n. sp.—Johnson: 28, pl. 9, figs. 3, 9
1991Permocalculusforcepinus (Johnson)—Roux: 152, pl. 2, fig. 3 (with synonymy)
Description
Species characterized by the deep segmentation of its thallus; i.e., a “waxing-and-waning” outline [consequently, P. fragilis forma moniliformis (Pia) sensu Kochansky-Devidé et Slisković (1969) and P. anatoliensis Güvenç, 1966 are probably junior synonyms]. The thallus is generally extensively calcified with disappearance of skeletal details.
Dimensions
L = 4.22–6.76 mm; D = 2.14–2.58 mm; s = 0.33–0.60 mm; p = 0.05–0.09 mm.
Occurrence
Middle Permian of Texas, Iraq, Armenia, ?Turkey. Murgabian (Middle Permian) of the Dena section.
cf. 1937Gymnocodium fragile n. sp.—Pia: 70, pl. 12, figs. 1–2
cf. 1955Permocalculusfragilis (Pia)—Elliott: 86, pl. 1, figs. 1–2
cf. 1960Permocalculus fragilis (Pia)—Herak and Kochansky-Devidé: 188, pl. 2, fig. 1–4, pl. 3, figs. 1–8
cf. 1963Permocalculus fragilis (Pia)—Herak and Kochansky-Devidé: 65, pl. 3, figs. 2–9, pl. 4, fig. 1
cf. 1977 Permocalculus fragilis (Pia)—Vachard in H. Termier et al.: 10, pl. 1, figs. 1–3, 5–6
non 1983Permocalculus cf. fragilis (Pia)—Schäfer and Senowbari-Daryan: 116, pl. 8, fig. 7
cf. 1991Permocalculusfragilis (Pia)—Roux: 152, pl. 2, figs. 4–5 (with synonymy)
Description
This alga is common in the lower and middle carbonate units of the Dena section. The thallus, when well preserved, is ovoid to spherical. The uncalcified medullar zone is large and the cortical zone thin. The density of the cortical file of cells is variable but occasionally high. They exhibit 3 or 4 ramifications in the cortical zone. Rare conceptacles located at the boundary between the medullar and cortical zones have been observed (Fig. 8c).
Dimensions
L = 1.69–4.70 mm; D = (0.52)–1.14–2.50 mm; d = 1.00–2.16 mm; s = (0.06)–0.14–0.20 mm; p = 0.03–0.085 mm.
Occurrence
Murgabian (Middle Permian) of the Dena section. This alga co-occurs with Mizzia sp., Permocalculus sp., and Epimastoporella spp.
1955Permocalculus plumosus n. sp.—Elliott,: 87, pl. 3, figs. 2–5
1982Permocalculus aff. plumosus Elliott—Mu: 214, pl. 1, fig. 3
1991Permocalculusplumosus Elliott—Roux: 158, pl. 2, figs. 11–12 (with synonymy)
Description
A number of longitudinal and oblique sections have been found in thin-sections the lower and middle carbonate units of Dena succession. The thallus is slightly segmented with ovoid-elongate elements. Some specimens exhibit a preserved medulla and relatively numerous conceptacles as dark oval cavities with diameters of 0.46–0.66 mm (Fig. 7h–j). The cortex is well calcified and the files of cells, almost perpendicular to the wall, often show three to four ramifications. The specimens from the Dena Mountain have generally larger diameters than those described by Elliott (1955).
Dimensions
L = 2.63–5.53 mm; D = 1.22–3.06 mm; d = 0.53–2.26 mm; s = 0.13–1.05 mm; p = 0.02–0.035 mm.
Occurrence
Murgabian (Middle Permian) of the Dena section. This alga co-occurs with Mizziayabei and Permocalculus spp.
cf. 1937Gymnocodiumsolidum n. sp.—Pia: 28, pl. 9, fig. 1?, pl. 13, figs. 1–2
cf. 1955Permocalculussolidus (Pia)—Elliott: 86, pl. 2, figs. 5––6
cf. 1969Permocalculus fragilis forma solidus (Pia)—Kochansky-Devidé and Slisković: 106, pl. 2, fig. 2
cf. 1991Permocalculussolidus (Pia) Elliott—Roux: 158, pl. 2, fig. 3
cf. 1992Succodium difficile Kordé—Okla: pl. 46, figs. 6–8
Description
The cylindrical to finger-shaped thalli are very well preserved in the cortical zone but also in the medullar one (Fig. 8d). Consequently, these specimens are almost at the stage of Pyrulites Mu, 1981 where the medullar zone is entirely preserved. Other evidence that Pyrulites is only a stage and no a genus or subgenus is its presence in the Cretaceous (Bucur 1994; Schlagintweit and Sanders 2007).
Dimensions
L = 2.22–2.76 mm; D = 2.74–2.88 mm; s = 0.33–0.60 mm; p = 0.02–0.05 mm.
Occurrence
Late Permian of the Carnic Alps, Saudi Arabia, Iraq, Tunisia, Pakistan. Murgabian (Middle Permian) of the Dena section.
According to Vachard et al. (2005), Tauridium is another preservational stage of Permocalculus where the files of cells are remarkably preserved. We have observed one specimen with this characteristic.
Dimensions
L = 2.76 mm; D = 2.18 mm; d = 1.76 mm; s = 0.17–0.20 mm; p = 0.075–0.09 mm.
Occurrence
The genus has been recorded from the late Middle and Late Permian of Turkey, Iraq, and Saudi Arabia. The form from Turkey, illustrated by Flügel (1990, pl. 4, fig. 7), is in fact an algosponge. This is the first, albeit doubtful, record of the genus from Iran.
Algae incertae sedis
Phylloid algae (green and/or red algae)
Description
Phylloid algae are present in our material as sparitized fragments with micro-perforations. The latter are interpreted either as siphons of codiaceans (e.g., Chuvashov et al. 1987; Mamet 1991) or as secondary small tunnels bored by possible bacteria (Vachard et al. 1989a). Different forms of siphons or microborings permit the distinction of several morphogenera, in particular Eugonophyllum Konishi and Wray, 1961 and Ivanovia Khvorova, 1946.
Remarks
We agree with Schlagintweit (2010) and Granier (2012) to consider the phylloid algae as an informal botanical group. Calcifolium and the calcifoliids differ totally (Vachard and Cózar 2010). Nevertheless, other forms such as Archaeolithophyllum, Anchicodium, Ivanovia, Eugonophyllum, and Neoanchicodium remain botanically and diagenetically related according to the scenarios previously proposed by Vachard et al. (1989a, 2001, 2012).
Two particular longitudinal sections have been observed in the lower carbonate unit of the Dena profile. They resemble some Eugonophyllum of the literature but might also correspond to gastropod and/or bivalve fragments perforated by microendolithic cyanobacteria or algae. The fragments are broad and irregularly arcuate. Their lengths range from 0.17 to 0.43 mm. On the surface, there are small and elongate tunnels, siphons or microborings with diameters of 0.03–0.07 mm and a length of 0.11–0.46 mm. The inner parts do not show a crystallized and structural medulla and cortex. Eugonophyllum? sp. is the only “phylloid alga” observed in the Dena section.
Occurrence
Murgabian (Middle Permian) part of the Dena section. This taxon has been found in high-energy carbonates associated with Permocalculus spp, and the foraminifer Pachyphloia spp.
Class Algospongia G. Termier et al. 1977a, b emend. Vachard and Cózar, 2010
Order Aoujgaliida G. Termier et al., 1975 emend. Vachard and Cózar, 2010
Family Aoujgaliidae G. Termier et al., 1975 nom. translat. Vachard and Cózar, 2010
Incertae sedis algae and microproblematica of the Dena section. Scale bars 1 mm. a-bPseudovermiporella sodalica; typical specimens; a sample D44; b sample D45. c, j-k, mUngdarella uralica; typical specimens; c sample D36; j sample D24; k sample D31; m sample D36. d, gFourstonella (Efluegelia) johnsoni; typical specimens; d sample D36; g sample D54. e–fStacheoides sp., typical specimens; e sample D49; f sample D 36. hPseudovermiporellanipponica; typical specimen; sample D19. iPseudovermiporella longipora; typical specimen; sample D19. l, pTubiphytes obscurus; different specimens; l sample D24; p sample D25. n–oAeolisaccus dunningtoni; typical specimens; n sample D25; o sample D52
In the lower carbonate unit of the Dena section, some Stacheoides sp. were found. They are attached, and consist of concentric rows of hemispherical cells or chambers. No aperture or connexion visible. Wall calcitic, yellowish, and hyaline. This species seems to be similar to that mentioned from Afghanistan by Vachard (1980).
Dimensions
L = 1.30–1.80 mm; cell dimensions = 0.03–0.05 mm × 0.05–0.10 mm; cell wall thickness = 0.01–0.03 mm.
Occurrence
Late Middle Mississippian-Late Permian (see references in Vachard and Cózar 2010). Cosmopolitan up to the Early Permian; after that limited to the shelves of the Paleo- and Neotethys. Murgabian (Middle Permian) part of the Dena section.
Family Ungdarellidae Maslov, 1956a emend. Vachard and Cózar, 2010
Description
Aoujgaliida showing an attached-to arborescent shape, cylindrical and branched, and embracing rows of the chambers. Occasionally, the initial stage is attached, similar to Stacheoides, followed by an erect stage. The endoskeleton is composed of conical to paraboloid concentric laminae growing upward or downward in continuous rows of chambers, and perforated pillars, transverse, perpendicular to each lamina, acting as communications between the chambers (Vachard 1980, fig. 74). Bifurcations of the laminae and thallus have the same orientation as the growth direction contrary to e.g., Komia Kordé, 1951. Wall calcitic, yellowish, and hyaline.
Occurrence
Middle Mississippian (late Visean zone MFZ14) to Late Permian (Vachard and Cózar 2010). Cosmopolitan up to the Early Permian; after that restricted to the Palaeo- and Neotethys.
1950 Ungdarella uralica n. gen. n. sp.—Maslov: 75, fig. 1
1956aUngdarella uralica Maslov—Maslov: 73, pl. 21, figs. 2–3, pl. 23, figs. 1–4, 18–19
1972 Ungdarella uralica Maslov—Homann: 155, pl. 1, fig. 4
1972Ungdarella uralica Maslov—Mamet and Rudloff: 91. pl. 9, figs. 1–5
1973Ungdarella uralica Maslov—Ivanova: pl. 21, fig. 8, pl. 28, figs. 4–5, pl. 34, fig. 7
1977Ungdarella cf. uralica Maslov—Mamet and Roux: 229, pl. 6, fig. 3
Description
This microfossil has been observed sporadically in lower and middle parts of the Dena profile. The thallus is rod-shaped, branched, with an apparently cellular construction. Due to the generally strong recrystallization, only yellow or cream-colored indistinct bodies are seen.
Dimensions
L = 1.39–4.16 mm; D = 0.46–0.81 mm; height of cell laminae = 0.01–0.017 mm and width of cells = 0.017–0.022 mm.
Comparison
Many species of Ungdarella exist in the literature. Several authors, however, consider that only the very polymorphic U. uralica should be recognized.
Occurrence
As for the genus, Permian representatives are relatively common in the Carnic Alps, Russia, Turkey, Armenia, Iraq, Afghanistan, Himalaya, and Iran (e.g., Kolodka et al. 2012). Murgabian (Middle Permian) part of the Dena section.
Family Stacheiidae Loeblich and Tappan, 1961 nomen translat. Vachard and Cózar, 2010
Description
Attached Aoujgaliida, showing many rows of quadratic cells or chamberlets with a uniserial, partly overlapping growth. Chamberlets square or higher than wide with distal and proximal, curved sides, and rectilinear lateral sides.
Occurrence
Late Viséan-Late Permian, probably cosmopolitan (especially Efluegelia).
Efluegelia can be synonymized with Fourstonella (see discussion in Vachard and Cózar 2010) or considered as a subgenus of Fourstonella because it replaces definitively the true Fourstonella at the end of the Moscovian and is the unique representative of the stacheiids during Late Pennsylvanian and Permian times (Vachard et al. 2012; this work). The subgenus is currently monospecific, although some undescribed new species exist in the Middle Pennsylvanian of Iran and Russia (Vachard, unpubl, data). Fourstonella (Efluegelia) differs from F. (Fourstonella) by the absence of axial symmetry of the thallus, some differences in the cell walls (Vachard et al. 1989b), and a difference in age. During the Permian, it has been commonly reported from Austria, Russia, Tunisia, Afghanistan, Oman, and Iran (Kolodka et al. 2012) and is well known in the USA (Toomey et al. 1988; Groves 1983; Mamet et al. 1987; Krainer et al. 2003b, 2009; Vachard et al. 2012).
Occurrence
Late Visean-Late Permian with an acme in the Late Pennsylvanian and Early Permian; cosmopolitan.
1966Cuneiphycus johnsoni n. sp.—Flügel: 17, pl. 2, figs. 1–5
1971Cuneiphycus johnsoni Flügel—Lys and Lapparent: 91
1979Eflugelia johnsoni (Flügel) n. gen. n. comb.—Massa and Vachard: 34
Discussion
Scattered specimens have been found in thin-sections from the lower unit of the Dena section. The yellowish thallus can display numerous morphological variations. It develops on different substrates, which are often not preserved, and is mostly found as fragments. Most of its structure is not quite recognizable due to recrystallization.
Dimensions
L = 0.68–0.98 mm; w = 0.86 mm; cell width = 0.01–0.02 mm; cell height = 0.015–0.025 mm; cell wall thickness = 0.005–0.01 mm.
Occurrence
As for the genus, Murgabian (Middle Permian) part of the Dena section.
1958Aeolisaccus dunningtoni n. gen. n. sp.—Elliott: 422, 424, pl. 3, figs 5–6, 8–9
2006 Earlandia dunningtoni (Elliott)—Krainer and Vachard: 69, pl. 2, figs. 8, 11–12, 17–18, pl. 6, fig. 8 (with synonymy)
2012Aeolisaccus dunningtoni Elliott—Hughes: pl. 4, figs. 1–15
Description
A few representatives of this microfossil have been found in thin-sections from the lower parts of the Dena section. It consists of carbonate tubes apparently open at both ends, although Gaillot and Vachard (2007) and Krainer and Vachard (2011) have encountered very rare specimens with a closed and slightly inflated extremity, which has been compared with to the proloculus of foraminifers by these authors. For other authors (e.g., Hughes 2012), the wall differs from that of the foraminifer Earlandia.
Dimensions
L = 0.84–0.90 mm; D = 0.06–0.14 mm; d = 0.04–0.12 mm; s = 0.018–0.02 mm.
Occurrence
The microfossil was first reported from Late Permian rocks of Iraq and subsequently from other parts of the world such as Afghanistan, Europe, and Iran (Gaillot and Vachard 2007). Murgabian (Middle Permian) part of the Dena section.
Consortial associations between foraminifers Miliolata (Vachard and Krainer 2001b; Vachard et al. 2010) with cyanobacterial algae. Poorly transformed chambered tubes were called Cordiformis Güvenç, 1965 (nom. nud.; see also Vachard and Montenat 1981, pl. 13, fig. 10; Vachard and Ferrière 1991, pl. 1, fig. 4), Foraminifer gen. et sp. indet. by Senowbari-Daryan and Flügel (1993, pl. 3, figs. 7–10, pl. 4, figs. 5–6), etc. Typical forms appear as masses of a rounded, thick-walled taxon with a small, rounded cavity (e.g., Razgallah and Vachard 1991; Senowbari-Daryan and Flügel 1993; Fig. 9l, p). The family crosses the Permio-Triassic boundary.
Occurrence
Late Middle Pennsylvanian (Late Moscovian)-Early Cretaceous. For Pennsylvanian forms, see Vachard and Moix (2011) and Vachard et al. (2012), and for the Mesozoic forms, see Senowbari-Daryan et al. (2008) and Schlagintweit and Gawlick (2009b). True Tubiphytes are limited to Permian strata (Vachard et al. 2012).
2003Tubiphytes ex gr. obscurus Maslov—Vachard et al.: pl. 4, figs. 1, 4–5
Description
Common in thin-sections from the lower and middle carbonate units of the Dena section. In general, these remains are common in material from Iran, where they are very diversified, and vary from morphotypes looking like foraminifers (compare Vachard et al. 1993b, pl. 3, fig. 4) to small domes with one or several cavities (compare Vachard et al. 1993b, pl. 3, fig. 1). Typical specimens were found at Kuh-e Gakhum (Zagros, Iran) by Gaillot (2006) in the Lower Dalan Formation. In contrast, Tubiphytes of the Dena section are commonly irregularly cylindrical in shape, with smooth external surface and characteristic dark walls. They are segmented and each segment has a spherical, oval, cylindrical, or annulated cavity that is filled with sparite.
Dimensions
Outer diameter = 0.45–0.55 mm, diameter of the central cavity = 0.10–0.125 mm, wall thickness = 0.075–0.15 mm. Typical forms attain 5–6 mm in length (Elliott 1962).
Occurrence
Throughout the Permian, from the earliest Wolfcampian to the latest Changhsinghian (Lys et al. 1978; Altiner and Özkan-Altiner 1998; Vachard et al. 2003, 2012). This microfossil has been reported from the Urals, Japan, Indonesia, Afghanistan, Himalaya, Guatemala, Mexico, USA, Algeria, Turkey, Iraq, Greece, Oman, China (Mu et al. 2005), and Iran (Vaziri et al. 2005; Kolodka et al. 2012). Its range is Midian-Wuchiapingian in the Zagros Mts (Gaillot 2006). Murgabian (Middle Permian) part of the Dena section.
1954Vermiporella nipponica n. sp.—Endo in Endo and Kanuma: 191, pl. 13, figs. 2–5
2007Pseudovermiporella nipponica (Endo)—Gaillot and Vachard: 82, pl. 58, fig. 16, pl. 59, figs. 4, 19 (with synonymy)
Description
The species is characterized by a moderately thick wall; pits straight, of small diameter.
Dimensions
L = up to 2.00 mm; D = 0.40–0.65 mm; d = 0.10–0.45 mm; s = 0.07–0.12 mm; pits diameter = 0.005–0.01 mm.
Occurrence
Relatively common and cosmopolitan from the Sakmarian (Vachard and Krainer 2001a; Krainer et al. 2003b, 2009) to the Changsinghian. Already mentioned in Zagros by Gaillot (2006) and Gaillot and Vachard (2007). Murgabian part of the Dena section.
The microfossil includes some tubular skeletons with circular, oval, or irregular transverse sections. Pits are perpendicular to the central chamber and are polygonal or rounded in tangential sections.
Dimensions
L = 0.96–2.74 mm; D = 0.56–1.08 (2.20) mm; d = 0.52–1.16 mm; s = 0.08–0.31 (0.69) mm; pits diameter = 0.02–0.06 mm (with a wall of 0.01–0.04 mm).
Comparison
The species differs from P. nipponica and P. longipora by the shape and dimensions of the pits. They are shorter and wider in P. nipponica, and larger and narrower in P. longipora. Pseudovermiporella sodalica is often confused with true algae [e.g., Mizzia; Gaetani et al. (2009) or Thaumatoporella; Schlagintweit (2011)].
Occurrence
Middle-Late Permian of Iraq, Oman, Afghanistan, Turkey, Hungary, Greece, Slovenia, Croatia, Armenia, North America, Tunisia, China, and SE Asia (Mu et al. 2005), and Iran (Kolodka et al. 2012). Murgabian (Middle Permian) part of the Dena section.
1954 Vermiporella nipponica var. longipora n. var.—Praturlon: 126, pl. 2, figs. 1–7
2003Pseudovermiporella ex gr. nipponica (Endo)—Vachard et al.: pl. 4, fig. 3
2007Pseudovermiporella nipponica (Endo)—Gaillot and Vachard: pl. 58, fig. 16, pl. 59, figs. 4, 19 (with synonymy)
Description
Some of these microfossils have been found in thin-sections from the lower and middle carbonate units of the Dena section. This microfossil is irregularly cylindrical. The pits are long, curved, relatively narrow, and thin-walled. Some pits divide into two branches outward.
Dimensions
L = 0.87–2.37 mm; D = 0.31–0.43 mm; s = 0.05–0.11 mm.
Comparison
Pseudovermiporella longipora differs from P. nipponica and P. solida by having more slender, larger, and curved pits (= pores of the authors).
Occurrence
Late Permian of Italy, Greece, Turkey, and Oman. Murgabian (Middle Permian) part of the Dena section.
Discussion
The described assemblage is traditional, being dominated by Mizzia and Permocalculus. We assigned all sections of Mizzia to a unique species, M. yabei, but we identified several species for our Permocalculus because (a) as illustrated our taxa have very different morphologies and (b) attempts to synonymize previous species (but often with the creation of new species) are rare or have never been confirmed (Elliott 1955; Kochansky-Devidé and Sliskovic 1969; Termier et al. 1977a, b; Vachard 1980; Roux 1991). Gymnocodium is quite rare in our material, but epimastoporellaceans and gyroporellaceans are relatively abundant and diversified. All these microfossils have a relatively broad distribution in the Permian period but are paleoecologically very important (Elliott 1968a; Vachard 1980; Gaillot 2006; Hughes 2012). They characterize very shallow but somewhat confined environments (with relatively high salinity and moderate water energy) above the fair-weather wave-base.
As light-sensitive organisms and thus indicators for light penetration, they are very important for paleoecological reconstructions. Paleobathymetric considerations for this group are based on the presence of modern dasycladales in the uppermost meters of the water column and in high water temperatures. Dasycladales are a parangon of the shallowest marine depths with a peak abundance at approximately 5–6 m, becoming rare between 10 and 30 m, and exceptional beyond these depths. The deepest ones were found at 90 m, but at a water temperature surpassing 17 °C (Elliott 1968a, 1977; Purser 1983; Roux 1985; Kirkland and Chapman 1990; Bucur and Sasaran 2005). Optimal temperatures of growth are quoted as 25–27 °C (Berger and Kaever 1992; Aguirre and Riding 2005). The dasycladales are all photophile but members of the group have different depth preference.
The modern caulerpales extend to a water depth of 120–150 m but co-dominate the fore-reef from 25–45 to 50–60 m (e.g., Purser 1983; Roux 1985; Blair and Norris 1988; Aponte and Ballantine 2001; Wilson and Vecsei 2005).
Permian true red algae are represented by Gymnocodium, Permocalculus, and rare solenoporaceans. Modern rhodophyta exhibit a wide depth range (Adey and Macintyre 1973; Purser 1983; Martindale 1992; Aponte and Ballantine 2001; Fagerstrom and Weidlich 2005; Wilson and Vecsei 2005; Perry 2005). Their accumulations or constructions exist between 0 m (algal ridge) to 180 m, with an acme between 60 and 120 m (Adey and Macintyre 1973; Roux 1985; Steneck et al. 1997; Aponte and Ballantine 2001); whereas the deepest Recent crustose coralline red alga was collected at a depth of 268 m (Aponte and Ballantine 2001). Relatively eurybathic, they nevertheless could constitute a high-resolution climate indicator for the mid and high latitude, today (Halfar et al. 2008).
Two groups of incertae sedis algae, very abundant during the Early Carboniferous, are still sporadically present in Permian times: (a) a group of tubular, septate, and ramified tests or thalli considered as green algae, microspongia, or undeterminate protists, which are called moravamminids or Paleoberesellaceae, and (b) a group of laminar, cellular or chambered tests or thalli considered as red algae, microspongia, or undeterminate protists, which are called Aoujgaliids or Ungdarellaceae. Both groups are encompassed in the class Algospongia assigned to incertae sedis algae or protists. According to their interpretations as algae or protists, their paleobathymetric implications are very controversial (e.g., Lees and Miller 1985; Hennebert and Lees 1985; Lees 1997; Vachard and Cózar 2010).
Conclusions
Eight genera and 22 species of calcareous algae and eight genera and five species of microproblematica were recognized in the Dena section, and some of them were investigated structurally in this study. Among the algae and problematic microfossils identified in this study, gymnocodiacean algae, problematic microfossils, and dasycladacean algae had the highest abundance in the lower carbonate unit of the Dena section. In the upper carbonate unit of the section, early Djulfian (= early Wuchiapingian) in age, there is a sharp decrease in the abundance of calcareous algae and problematic microfossils. The calcareous algae studied in this section are widespread in the lower carbonate unit in lagoonal and shallow-water environments. These true algae are accompanied by problematic algae or by consortia with algae or cyanobacteria that are also photophile, including Tubiphytes obscurus (with thick external wall) and Ungdarella uralica indicating clear water with depths below 25 m. Therefore, the lower carbonate unit of the Dalan Formation in this section formed in an area exposed to light and wave energy.
References
Accordi B (1956) Calcareous algae from the Upper Permian of the Dolomites (Italy), with stratigraphy of the “Bellerophon-zone”. J Paleont Soc India 1:75–84
Aghanabati A (2004) Geology of Iran. Geol Surv Iran, Teheran, 586 p (in Farsi)
Aguirre J, Riding R (2005) Dasycladalean algal biodiversity compared with global variations in temperature and sea level over the past 350 Myr. Palaios 20:581–588
Altiner D, Özkan-Altiner S (1998) Baudiella stampflii n. gen. n. sp., and its position in the evolution of Late Permian ozawainellid fusulines. Rev Paléobiol 17:163–175
Anderson KD, Beauchamp B (2010) The origin and ecology of late Paleozoic Palaeoaplysina in Arctic Canada: an aberrant ancestral coralline alga that grew at a time of high atmospheric CO2. In: GeoCanada Conference Abstract, Calgary, Alberta
Aponte NE, Ballantine DL (2001) Depth distribution of algal species on the deep insular fore reef at Lee Stocking Island, Bahamas. Deep-Sea Res I 48:2185–2194
Baghbani D (1988) Shanita zone and its biostratigraphic significance in south and southwest Iran. In: Benthos’1986, Genève, 37 p
Baghbani D (1996) Lithostratigraphy and biostratigraphy of the Permian sediments in the Shareza. Abadeh and Jolfa areas (Central and North west Iran). Unpubl. PhD Thesis. Islamic Azad Univ, Science and Research Branch, 233 p
Baghbani D (1997) Correlation charts of selected Permian strata from Iran. Permophiles 30:24–26
Blair SM, Norris JN (1988) The deep-water species of Halimeda Lamouroux (Halimedaceae, Chlorophyta) from San Salvador Island, Bahamas: species composition, distribution and depth records. Coral Reefs 6:227–236
Blomeier D, Dustira A, Forke H, Scheibner C (2011) Environmental change in the Early Permian of NE Svalbard: from a warm-water carbonate platform (Gipshuken Formation) to a temperate, mixed siliciclastic-carbonate ramp (Kapp Starostin Formation). Facies 57:493–523
Bucur II (1994) Lower Cretaceous Halimedaceae and Gymnocodiaceae from southern Carpathians and Apuseni Mountains (Romania) and the systematic position of the Gymnocodiaceae. Beitr Paläont 19:13–37
Canuti P, Marcucci M, Pirini Radrizzani C (1970) Microfacies e microfaune nelle formazioni paleozoiche dell’anticlinale di Hazro (Anatolia sud-orientale, Turchia). Boll Soc Geol Ital 89:21–40
Chuvashov BI (1974) Permskie izvestkovye vodorosli Urala (Permian calcareous algae from Urals). In: Papulov GN, Chuvashov BI (eds) Vodorosli, brakhiopodi i miospory iz permskikh otlozhenii zapadnogo Urala (Algae, brachiopods and miospores from the Permian deposits of western Urals). Akademiya Nauk SSSR, Uralskii Nauchnyi Tsentr, Trudy Inst Geol Geokhim 109:1–76 (in Russian)
Chuvashov BI (2001) Taxonomy, biostratigraphy and biogeography of the Permian Mizzia-like calcareous algae (Dasycladales, Cyclocrinaceae). Paleont J 35:97–104
Chuvashov BI, Anfimov AL (1988) Novye izvestkovye vodorosli srednego karbona-nizhnei permi Urala i Priuralya (New calcareous algae from Middle Carboniferous to Early Permian of Urals and Preural). In: Zhuravleva IT, Puchkov VM (eds) Izvestkovye vodorosli i stromatolity (calcareous algae and stromatolites). Akad Nauk SSSR, Sib Otdel, Inst Geol Geofiz, “Nauka”pp 54–70 (in Russian)
Chuvashov BI, Luchinina VA, Shuysky VP, Shaikin IM, Berchenko OI, Ishchenko AA, Saltovskaya VD, Shirshova DI (1987) Iskopaemye izvestkovye vodorosli morfologiya, sistematika, metody izucheniya (Fossil calcareous algae, morphology, systematics, methods of study). Akade Nauk SSSR, Sib Otdel, Trudy Inst Geol Geofiz 674:5–224 (in Russian)
Cózar P, Somerville ID (2004) New algal-foraminiferal evidence for the recognition of the Asbian-Brigantian boundary in northern England. Proc Yorkshire Geol Soc 55:43–65
Cózar P, Vachard D (2004) A new Mississippian dasyclad alga (Chlorophyta) from Spain: implications for the reproductive evolution of the dasyclads during the Late Paleozoic. Eclog geol Helv 97:175–181
Cózar P, Somerville ID, Medina-Varea P (2005) Note on the earliest occurrences of the calcareous algae Paraepimastopora and Archaeolithophyllum in Mississippian rocks. Coloq Paleont 55:7–20
Cózar P, Vachard D, Somerville ID, Pille L, Medina-Varea P (2009) Revision and new species of the Late Palaeozoic dasyclad algae Windsoporella and Eovelebitella. Palaeontographica B 282:1–24
Deloffre R, Granier B (1991) Hypothèse phylogénique des algues dasycladales. C R Acad Sci Paris 312, sér II:1673–1676
Dragastan O, Mertmann D, Ahmed S (1990) Some algal species from the Upper Permian deposits, Sakesar-section, Salt Range, Pakistan. Rev Paléobiol 9:49–63
Elliott GF (1977) Inferred isocrymal distribution of Jurassic dasycladacean algae in Europe, North Africa and southwestern Asia. J Geol Soc Lond 133:363–373
Elliott GF, Süssli P (1975) Imperiella gen. nov., a new alga from the Ruteh Limestone, Upper Permian (Central Alborz Mountains, north Iran). Eclog Geol Helv 68:449–455
Emberger J (1976) Les algues (Chlorophyceae, Prasinophyceae, Rhodophyceae) du Carbonifère et du Permien. Essai d’un inventaire bibliographique, géographique, stratigraphique. Bull Inst Géol Bassin d’Aquit, num spec: 1–168
Endo R (1951) Stratigraphical and paleontological studies on the Later Paleozoic calcareous algae in Japan I. Several new species from the Sakamotozawa section, Hikoroichi-mura, Kesen-gun, in the Kitakami Mountainous Land. Trans Proc Paleont Soc Jpn NS 4:121–129
Endo R (1953) Stratigraphical and paleontological studies of the Later Paleozoic calcareous algae in Japan, V. Several species from the Iwaizaki limestone, Motoyoshi-gun, in the Kitakami mountainous land. Trans Jpn J Geol Geogr 23:117–126
Endo R (1959) Stratigraphical and paleontological studies of the Later Paleozoic calcareous algae in Japan. XIV. Fossil algae from the Nyugawa Valley in the Hida Masssif. Sci Rep Saitama Univ Ser B 3:177–207
Endo R (1969) Fossil algae from the Khao Phong Phrab District in Thailand. In: Kobayashi T, Torixama R (eds) Contribution to the geology and palaeontology of Southeast Asia 74, Geol Palaeont Southeast Asia 7:33–85
Endo R, Hashimoto W (1955) Unquestionably Paleozoic (Permian) fossils found in Hokkaido, Japan. Proc Jpn Acad 31:704–708
Endo R, Kanuma M (1954) Stratigraphical and paleontological studies of the Later Paleozoic calcareous algae in Japan, VII. Geology of the Mino Mountain Land and southern part of Hida Plateau with description of the algal remains found in those districts. Sci Rep Saitama Univ, comment vol dedicated to Professor Riuji Endo, Ser B 1:177–205
Ernst A, Parvizi T, Rashidi R (2011) Some Bryozoa from the Upper Permian Dalan Formation of Dena Mountain in SW Iran. Freiberg Forschh 19:71–81
Fagerstrom JA, Weidlich O (2005) Biologic response to environmental stress in tropical reefs: lessons from modern Polynesian coralalgal atolls and Middle Permian sponge and Shamovella-microbe reefs (Capitan Limestone USA). Facies 51:501–515
Flügel E (1977) Environmental models for Upper Paleozoic benthic calcareous algal communities. In: Flügel E (ed) Fossil algae, recent results and developments. Springer, Berlin Heidelberg New York, pp 314–343
Flügel E (1980) Die Mikrofazies der Kalke in den Trogkofel-Schichten der Karnischen Alpen. In: Flügel E (ed) Die Trogkofel-Stufe im Unterperm der Karnischen Alpen. Carinthia II 36:51–99
Flügel E (1990) “Einschnitte” in der Entwicklung permischer Kalkalgen. Mitteilungen Naturwiss Ver Steiermark 120:99–124
Flügel E, Di Stefano P, Senowbari-Daryan B (1990) Lercaritubus problematicus n. gen. n. sp., a lower Permian reef organism from western Sicily. Boll Soc Paleont Ital 29:361–366
Flügel E, Senowbari-Daryan B, Di Stefano P (1991) Microfacies and depositional structure of allochthonous carbonate base-of-slope deposits: the Late Permian Pietra di Salomone Megablock, Sosio Valley (western Sicily). Facies 25:147–186
Gaetani M, Angiolini L, Ueno K, Nicora A, Stephenson MH, Sciunnach D, Rettori R, Price GP, Sabouri J (2009) Pennsylvanian-Early Triassic stratigraphy in the Alborz Mountains (Iran). In: Brunet MF, Wilmsen M, Granath JW (eds) South Caspian to Central Iran Basins. Geol Soc Lond Spec Publ 312:79–128
Gaillot J (2006) The Late Permian-Early Triassic Khuff Formation in the Middle-East, sequence biostratigraphy and palaeoenvironments by means of calcareous algae and foraminifers. Unpubl. Thèse PhD Univ Lille 1, 687 p
Gaillot J, Vachard D (2007) The Khuff Formation (Middle East) and time equivalents in Turkey and South China: biostratigraphy from Capitanian to Changhsingian times (Permian), new foraminiferal taxa, and palaeogeographical implications. Coloq Paleont 57:37–223
Gómez-Espinosa C, Vachard D, Buitrón-Sánchez B, Almazán-Vázquez E, Mendoza-Madera C (2008) Pennsylvanian fusulinids and calcareous algae from Sonora (Northwestern Mexico), and their biostratigraphic and paleobiogeographic implications. C R Palevol 7:259–268
Gortani M (1906) Contribuzioni allo Studio del Paleozoico Carnico. I. La Fauna Permo-Carbonica del col Mezzodi presso Forni Alvoltri. Palaeontogr Ital 12:1–84
Granier B (2012) The contribution of calcareous green algae to the production of limestones: a review. In: Basso D, Granier B (eds) Calcareous algae and the global change: from identification to quantification. Geodiversitas 34:35–60 doi: 10.5252/g2012n1a3
Granier B, Deloffre R (1995) Inventaire critique des algues dasycladales fossiles, IIIe partie: les algues dasycladales du Permien et du Trias. Rev Paléobiol 14:49–84
Granier B, Grgasović T (2000) Les Algues Dasycladales du Permien et du Trias. Nouvelle tentative d’inventaire bibliographique, géographique et stratigraphique. Geol Croat 53:1–197
Groves JR (1983) Calcareous foraminifers and algae from the type Morrowan (Lower Pennsylvanian) region of northeastern Oklahoma and northwestern Arkansas. Oklahoma Geological Survey, Bulletin 133:1–39
Groves JR (1984) Foraminifers and biostratigraphy of the Arco Hills, Bluebird Mountain, and lower Snaky Canyon Formations (Mid-Carboniferous) of East-Central Idaho. J Foram Res 14:282–302
Gümbel CW (1872) Die sogenannten Nulliporen (Lithothamnium und Dactylopora) und ihre Betheiligung an der Zusammensetzung der Kalkgesteine. Zweiter Theil: die Nulliporen des Thierreichs (Dactyloporideae) nebst Nachtrag zum ersten Theile. Abh königl bayer Akad Wiss Math-Phys Classe 11:231–290
Güvenç T (1965) Etude stratigraphique et micropaléontologique du Carbonifère et du Permien des Taurus occidentaux dans l’arrière-pays d’Alanya (Turquie). Unpubl. Thèse de Doctorat d’Université, Paris, 273 p
Güvenç T (1966) Description de quelques espèces d`algues calcaires (Gymnocodiacées et Dasycladacées) du Carbonifère et du Permien des Taurus occidentaux (Turquie). Rev Micropaléont 9:94–103
Henbest LG (1963) Biology, mineralogy, and diagenesis of some typical late Paleozoic sedimentary foraminifera and algal-foraminiferal colonies. Cushman Found Foram Res Spec Publ 6:1–44
Herak M, Kochansky-Devidé V (1963) Bükkhégysegi Újpaleozóos Mészalgak (Jungpaläozoische Kalkalgen aus dem Bükk-Gebirge (Nordungarn)). Geol Hung Ser Palaeont 28:45–78
Heydari E, Hassanzadeh J, Wade W, Ghazi A (2003) Permian Triassic boundary interval in the Abadeh section of Iran with implications for mass extinction. Part 1. Sedimentology. Palaeogeogr Palaeoclimatol Palaeoecol 193:405–423
Homann W (1972) Unter- und tief-mittelpermische Kalkalgen aus den Rattendorfer Schichten, dem Trogkofel-Kalk und dem Treßdorfer Kalk der Kanischen Alpen (Österreich). Senckenbergiana Leth 53:135–313
Hughes GW (2005) Saudi Arabian Permo-Triassic biostratigraphy, micropaleontology and palaeoenvironment. In: Powell AJ, Riding JB (eds) Recent developments in applied biostratigraphy. Micropalaeont Soc Spec Publ 91–108
Hughes GW (2012) Late Permian to Late Jurassic microproblematica of Saudi Arabia: possible palaeobiological assignments and roles in the palaeoenvironmental reconstructions. GeoArabia 18:57–92
Hüneke H, Joachimski M, Buggisch W (2001) Marine carbonate facies in response to climate and nutrient level: the Upper Carboniferous and Permian of Central Spitsbergen (Svalbard). Facies 45:93–136
Insalaco E, Virgone A, Courme B, Gaillot J, Kamali M, Moallemi A, Lotfpour M, Monibi S (2006) Upper Dalan member and Kangan Formation between the Zagros Mountains and offshore Fars, Iran: depositional system, biostratigraphy and stratigraphic architecture. GeoArabia 11:75–176
Iranian-Japanese Research Group (1981) The Permian and the lower Triassic systems in Abadeh Region, Central Iran. Mem Fac Sci Kyoto Univ, Ser Geol Min 47(2):61–133
Johnson B (1981) Microfaunal biostratigraphy of the Dalan Formation (Permian) Zagros Basin, southwest Iran. In: Neale JW, Brasier MD (eds) Microfossils from Recent and fossil shelf seas. British Micropalaeont Soc Series, Ellis Horwood, pp 52–61
Johnson JH, Konishi K (1956) Mississippian Algae from the Western Canada Basin and Montana. In: Johnson JH, Konishi K (eds) Studies of Mississippian algae. Quart Colorado School Mines 51:85–107
Kalantari A (1986) Microfacies of carbonate rocks of Iran. Ministry of Oil, NIOC, 287 p
Karpinsky A (1909) Einige problematische Fossilien aus Japan. Verh Russ Miner Ges St. Petersburg 2:257–272
Khvorova IV (1946) O novom rode vodoroslei iz srednekamennougolnykh otlozhenii Podmoskovnogo basseina (On a new genus of algae from the Middle Carboniferous deposits of the Submoscovite Basin). Dokl Akad Nauk SSSR 53:741–744 (in Russian, English translation 737–739)
Kirkland BL (1995) Distribution of dasycladacean algae in the Permian Capitan Formation and Carlsbad group, Guadalupe mountains, Texas and New Mexico, U.S.A. In: Piller WE (eds) Proceedings of international symposium field meeting “Alpine Algae 93”. Beitr Paläont Wien 19:153–159
Kirkland BL, Chapman RL (1990) The fossil green alga Mizzia (dasycladaceae): a tool for interpretation of paleoenvironment in the Upper Permian Capitan Reef Complex, southeastern New Mexico. J Phycol 26:569–576
Kobayashi F, Ishii KI (2003) Paleobiogeographic analysis of Yahtashian to Midian fusulinacean faunas of the Surmaq Formation in the Abadeh region, central Iran. J Foram Res 33:155–165
Kochansky-Devidé V, Gusic I (1971) Evolutions-Tendenzen der Dasycladaceen mit besonderer Berücksichtigung neuer Funde in Jugoslawien. Paläont Z 45:82–91
Kochansky-Devidé V, Slisković T (1969) Gornjopermski Mikrofosili Han Orahoviće i Suljaca u Bosni s osobitim obzirom na algu Permocalculus fragilis. Geol Vjesnik Inst Geol Istraz Zagrebu 22:105–110
Kolodka C, Vennin E, Vachard D, Trocme V, Goodarzi M (2012) Timing and progression of the end-Guadalupian crisis in the Fars province (Dalan Formation, Kuh-e Gakhum, Iran) constrained by foraminifers and other carbonate microfossils. Facies 58:131–153
Konishi K (1955) Succodium, a new codiacean genus, and its algal associates in the late Permian Kuma Formation of southern Kyushu, Japan. Tokyo Univ Fac Sci 2(9):225–240
Kordé KB (1951) Novye rody i vidy izvestkovykh vodoroslei iz kamennougolnykh otlozhenii Severnogo Urala (New genera and species of calcareous algae from Carboniferous deposits of northern Urals). Byull Moskov Obshch Ispyt Prir, Otdel Geol 1:175–182 (in Russian)
Kordé KB (1965) Algae. In: Ruzhentsev VE, Sarycheva TG (eds) Razvitie i smena morskikh organizmov na rubezhe Paleozoya i Mesozoya (Development and succession of marine organisms at the boundary Paleozoic and Mesozoic). Akad Nauk SSSR, Trudy Paleont Inst 108:268–284 (in Russian)
Kordé KB (1973) Vodorosli Kembriya (Cambrian algae). Akademiya Nauk SSSR, Trudy Paleont Inst 139:1–349 (in Russian)
Krainer K (2007b) Gzhelian Anthracoporella mounds, Carnic Alps, Austria. In: Vennin E, Aretz M, Boulvain F, Munnecke A (eds) Facies from Palaeozoic reefs and bioaccumulations. Mém Mus Nat Hist natur 195:283–286
Krainer K, Vachard D (2002) Late Serpukhovian (Namurian A) microfacies and carbonate microfossils from the Carboniferous of Nötsch (Austria). Facies 46:1–26
Krainer K, Vachard D (2007) Sakmarian-Artinskian Shamovella (= Tubiphytes)-mounds, Carnic Alps. In: Vennin E, Aretz M, Boulvain F, Munnecke A (eds) Facies from Palaeozoic reefs and bioaccumulations. Mém Mus Nat Hist natur 195:291–294
Krainer K, Vachard D (2011) The Lower Triassic Werfen Formation of the Karawanken Mountains (southern Austria) and its disaster survivor microfossils, with emphasis on Postcladella n. gen. (Foraminifera, Miliolata, Cornuspirida). Rev Micropaléont 54:59–85
Krainer K, Flügel E, Vachard D, Joachimski M (2003a) A close look at late carboniferous algal mounds: schulterkofel, Carnic Alps, Austria. Facies 49:325–350
Krainer K, Vachard D, Lucas SG (2003b) Microfacies and microfossil assemblages (smaller foraminifers, algae, pseudo-algae) of the Hueco Group and Laborcita Formation (Upper Pennsylvanian-Lower Permian), South-Central New Mexico, USA. Riv Ital Paleont Stratigr 109:3–36
Krainer K, Vachard D, Lucas SG (2005) Lithostratigraphy and biostratigraphy of the Pennsylvanian-Permian transition in the Jemez Mountains, north-central New Mexico. In: Lucas SG, Zeigler KE, Spielmann JA (eds) The Permian of Central New Mexico. New Mexico Mus Nat Hist Sci 31:74–89
Krainer K, Lucas SG, Vachard D (2007) Wolfcampian Laborcita mound complex, New Mexico, USA. In: Vennin E, Aretz M, Boulvain F, Munnecke A (eds) Facies from Palaeozoic reefs and bioaccumulations. Mém Mus Nat Hist natur 195:287–290
Krainer K, Vachard D, Lucas SG (2009) Facies, microfossils (smaller foraminifers, calcareous algae) and biostratigraphy of the Hueco Group, Doña Ana Mountains, southern New Mexico, USA. Riv Ital Paleont Stratigr 115:3–26
Krotov P (1988) Geologicheskie issledovaniya na zapadnom sklone Solikamskogo i Cherdynskogo Urala (Geological investigations on the western slope of Urals in the areas of Solikam and Cherdyn). Trudy Geol Kom St Pétersbourg 6:551–553
Kulik EL (1978) Izvestkovye zelenye (sifonovye) vodorosli asselskogo i sakmarskogo jarusov biogermnogo massiva Shakhtau (Bashkiriya) (Calcareous green (siphonal) algae of the Asselian and Samkmarian stages of the Shakhtau bioherm massif (Bashkiria)). Voprosy Mikropaleont 21:182–215
Kützing FT (1843) Phycologia Generalis oder Anatomie, Physiologie und Systemkunde der Tange. F. A. Brockhaus, Leipzig, xxxii + 142 p
Lees A (1997) Biostratigraphy, sedimentology and palaeobathymetry of Waulsortian buildups and peri-Waulsortian rocks during the late Tournaisian regression, Dinant area, Belgium. Geol J 32:1–36
Lucas SG, Krainer K, Vachard D (2011) Stratigraphy, depositional environments, age, and regional tectonic significance of the Powwow Member of the Hueco Canyon Formation, Lower Permian of the Hueco Mountains, West Texas. West Texas Geol Soc Bull 50:20–40
Lys M, Stampfli G, Jenny J (1978) Biostratigraphie du Carbonifère et du Permien de l’Elbourz oriental (Iran du NE). Notes Lab Paléont Univ Genève 13:63–99
Mamet B, Villa E (2004) Calcareous marine algae from the Carboniferous (Moscovian-Gzhelian) of the Cantabrian zone (NW Spain). Rev Espan Paleont 19:151–190
Maslov VP (1950) Znaschenie bagryanykh vodoroslei dlya stratigrafii SSSR (Importance of red algae for the stratigraphy of USSR). Dokl Akad Nauk SSSR 70:75–78 (in Russian)
Maslov VP (1956a) Onovom iskopaemon semeistve bagryanikh i dvukh novykh rodakh sinezelenykh vodoroslei (New Carboniferous family of fossil red algae and two new Cyanophycean genera). Dokl Akad Nauk SSS 107:151–154 (in Russian, French translation CEDP 1381)
Maslov VP (1956b) Iskopaemye izvestkovye vodorosli SSSR (Fossil calcareous algae from USSR). Trudy Inst Geol Nauk, Akad Nauk SSSR 160:1–301 (in Russian; French translation BRGM no 3517)
Maslov VP (1973) Atlas porodoobrazufschchikh organizmov (izvestkovykh i kremnevykh) (Atlas of rock-building organisms (calcareous and siliceous)). Izdat “Nauka” pp 1–111 (in Russian)
Massa D, Vachard D (1979) Le Carbonifère de Lybie Occidentale: biostratigraphie et micropaléontologie; position dans le domaine téthysien d’Afrique du Nord. Rev Inst Français Pétrole 34:3–65
Milanović M (1965a) Zwei neue Gattungen der Familie Dasycladaceae aus dem Perm des Velebitgebirges. Bull Sci Cons Acad, RSF Yougoslavie, Sect A 10(6):179–180
Mohtat-Aghai P, Vachard D (2005) Late Permian foraminiferal assemblages from the Hambast region (central Iran) and their extinctions. Rev Esp Micropaleont 37:205–227
Mu X (1982) Some calcareous algae from Xizang. In: The series of the scientific expeditions to the Qinghai-Xizang Plateau. Paleontology of Xizang, Book V Science Press, Beijing, pp 205–238, (in Chinese)
Mu X, Elliott GF (1982) Nanjinoporella, a new Permian dasyclad (calcareous alga) from Nanjing, China. Bull Brit Mus (Nat Hist) Geol 36:109–116
Mu X, Huichun Y, Yue L (2005) The latest Permian calcareous algae of the Changxingian from the lower Yangtze region, East China, and the establishment of a new algal province: the Pyrulites Province. 5th regional symposium of the IFAA 1:34–35
Noé SU (1987) Facies and paleogeography of the marine Upper Permian and of the Permian-Triassic boundary in the Southern Alps (Bellerophon Formation, Tesero Horizon). Facies 16:89–142
Pal AK (1976) The algal family Dasycladaceae, its taxonomy and evolution. In: Proceedings of 6th Indian colloqu micropaleontology and stratigraphy pp 154–181
Partoazar H (1995) Permian deposit in Iran. Geol Surv Iran 22:1–340
Pille L (2008) Foraminifères et algues calcaires du Mississippien supérieur (Viséen supérieur-Serpukhovien): rôles biostratigraphique, paléoécologique et paléogéographique aux échelles locale, régionale et mondiale. Unpubl. Thèse Univ Lille, Lille, 226 + 255 + 72 p
Pille L, Vachard D, Argyriadis I, Aretz M (2010) Revision of the late Visean-Serpukhovian (Mississippian) calcareous algae, foraminifers and microproblematica from Balia Maden (NW Turkey). Geobios 43:531–546
Radoičić R (2004) Permocalculus nikolapantici n. sp. (Udoteaceae?) from the Upper Turonian of NW Serbia. Bull Acad Serbe Sci Arts, Cl Sci Math Natur, Sci Natur 42:237–257
Rashidi K, Senowbari-Daryan B (2010) Dasycladales from the Permian Jamal Formation of the Shotori Mountains, northeast Iran. Part I: Imperiella Elliott and Süssli, Nanjinoporella Mu and Elliott, Tabasoporella nov. gen., and Pseudotabasoporella nov. gen. Facies 56:111–137
Rauzer-Chernousova DM (1949) (sometimes indicated as 1951). Stratigrafiya verkhnekammennougolnyhk i artinskikh otlozhenii bashkirskogo Priuralya (Stratigraphy of the Late Carboniferous and Artinskian deposits from the Bashkir Pre-Urals). Akad Nauk SSR, Trudy Inst Geol Nauk 105, Geol Ser 35:1–21 (in Russian, French translation by Pietresson de St-Aubin, Institut Français du Pétrole, des Carburants et Lubrifiants, no. 1522–747:1–18)
Rauzer-Chernousova DM (1950) Fatsii verkhnekamennougolnykh i artinskikh otlozhenii Sterlitamaksko-Ishimbayskogo Priuralya (na osnove izucheniya fuzulinid). (Facies of Late Carboniferous and Artinskian deposits in the Sterlitamak-Ishimbaevo region of the Pre-Urals, based on a study of fusulinids). Akad Nauk SSSR, Trudy Inst Geol Nauk 119, Geol Ser 43:1–109 (in Russian)
Razgallah S, Vachard D (1991) Systématique et biosédimentologie des algues constructrices permiennes Tubiphytes et Archaeolithoporella suivant l’exemple du Jebel Tebaga (Murghabien de Tunisie). Palaeontographica B 221:171–205
Reid CM, James NP, Beauchamp B, Kyser TK (2007) Faunal turnover and changing oceanography: Late Palaeozoic warm-to-cool water carbonates, Sverdrup basin, Canadian Arctic Archipelago. Palaeogeogr Palaeoclimatol Palaeoecol 249:128–159
Roux A (1985) Introduction à l’étude des algues fossiles paléozoïques (de la bactérie à la tectonique des plaques). Bull Centr Rech Explor-Product Elf-Aquitaine 9:465–699
Roux A (1989) Study of a Paraepimastopora kansasensis (dasyclad alga) topotypical material from the Upper Pennsylvanian of Kansas, USA. Rev Paléobiol 8:323–333
Roux A (1991) Révision des Gymnocodiaceae (algues rouges Permien–Crétacé). Taxonomie, biostratigraphie, paléobiogéographie, 2e partie: inventaire taxonomique critique des espèces de Gymnocodiacées du Permien et du Trias. Rev Micropaléont 34:136–173
Samankassou E (1999) Drowning of algal mounds: records from the upper carboniferous lower Pseudoschwagerina Limestone, Carnic Alps, Austria. Sediment Geol 127:209–220
Samankassou E (2002) Cool-water carbonates in a paleoequatorial shallow-water environment: the paradox of the Auernig cyclic sediments (Upper Pennsylvanian, Carnic Alps, Austria-Italy) and its implications. Geology 30:655–658
Sánchez-Chico F, Mamet B, Moreno-Eiris E, Rodríguez S (1995) Algas calcáreas del Viseense de Los Santos de Maimona (Badajoz). Rev Esp Micropaleont 27:67–96
Schlagintweit F (2010) Gosavisiphon gen. nov. based on Halimeda paucimedullaris Schlagintweit and Ebli, 1998, a remarkable macroalga (Udoteaceae?) from the Late Cretaceous of the Northern Calcareous Alps (Austria and Germany) with affinities to Late Paleozoic and Late Triassic phylloids. Geol Croat 63:27–53
Schlagintweit F (2011) Remarks on the Pseudovermiporella Elliott, 1958 and the origin of thaumatoporellacean algae. In: Bucur II, Sasaran E (eds) Abstracts, 10th international symposium on Fossil Algae, Cluj-Napoca, Romania, 12–18 September 2011, Cluj, pp 76–77
Schlagintweit F, Gawlick H-J (2009a) Oncoid-dwelling foraminifera from the Late Jurassic shallow-water carbonates of the Northern Calcareous Alps (Austria and Germany). Facies 55:259–266
Schlagintweit F, Gawlick H-J (2009b) Enigmatic tubes associated with microbial crusts from the Late Jurassic of the Northern Calcareous Alps (Austria): a mutualistic sponge-epibiont consortium? Lethaia 42:452–461
Schlagintweit F, Sanders D (2007) Permocalculus (Pyrulites) theresiensteinensis n. sp., a new calcareous alga from the Upper Cretaceous Lower Gosau Subgroup of the Northern Calcareous Alps (Austria). Jb Geol B-A 147:607–614
Schönlaub HP, Forke HC (2007) Die post-variszische Schichtfolge der Karnischen Alpen–Erläuterungen zur geologischen Karte des Jungpaläozoikums der Karnischen Alpen 1: 12500. Abh Geol B-A 61:3–157
Senowbari-Daryan B, Flügel E (1993) Tubiphytes Maslov, an enigmatic fossil: classification, fossil record and significance through time, part I: Discussion of Late Paleozoic material. In: Barattolo F, De Castro P, Parente M (eds) Studies on fossil benthic algae. Boll Soc Paleont Ital spec vol 1:353–382
Senowbari-Daryan B, Rashidi K (2010) The codiacean genera Anchicodium Johnson, 1946 Iranicodium nov. gen. from the Permian Jamal Formation of Shotori Mountains, Northeast Iran. Riv Ital Paleont Stratigr 116:3–21
Senowbari-Daryan B, Rashidi K (2011) Lercaritubus problematicus Flügel, Senowbari- Daryan and Di Stefano and Vangia telleri (Flügel): two problematic organism from the Permian Jamal Formation of Shotori Mountains: northeast Iran. Riv Ital Paleont Stratigr 117:105–114
Senowbari-Daryan B, Bucur II, Schlagintweit F, Sasaran E, Matyszkiewicz J (2008) Crescentiella, a new name for “Tubiphytes” morronensis Crescenti, 1969: an enigmatic Jurassic-Cretaceous microfossil. Geol Croat 61:185–214
Soreghan GS, Soreghan MJ, Hamilton MA (2008) Origin and significance of loess in late Paleozoic western Pangaea: a record of tropical cold? Palaeogeogr Palaeoclimatol Palaeoecol 268:234–259
Sremac J (2007) The Palaeozoic of Velebit Mt. In: Grgasović T, Vlahović I (eds) 9th International symposium on fossil algae, field trip guidebook, pp 13–19
Stampfli GM (1978) Etude géologique générale de l’Elbourz oriental au sud de Gonbad-e-Qabus, Iran NE. Unpubl. Thèse Univ Genève, Genève, 329 p
Steneck RS, Macintyre IG, Reid RP (1997) A unique algal ridge system in the Exuma Cays, Bahamas. Coral Reefs 16:29–37
Termier H, Termier G, Vachard D (1977b) Monographie paléontologique des affeurements permiens du Djebel Tebaga (Sud Tunisien). Palaeontographica A 156:1–109
Vachard D (1980) Téthys et Gondwana au Paléozoïque supérieur; les données afghanes: biostratigraphie, micropaléontologie, paléogéographie. Doc Trav IGAL, Inst Géol Albert de Lapparent 2:1–463
Vachard D (1985) Remarques sur les Dasycladales (Algues vertes) du Permien supérieur du Djebel Tebaga. 1er Congrés national Sciences Terre, Tunis 1981 vol 1, pp 271–281
Vachard D (1994) Foraminifères et moravamminides du Givétien et du Frasnien du domaine Ligérien (Massif Armoricain, France). Palaeontographica A 231:1–92
Vachard D, Ferrière J (1991) Une association à Yabeina (foraminifère fusulinoïde) dans le Midien (Permien supérieur) de la région de Whangaroa (Baie d’Orua, Nouvelle-Zélande). Rev Micropaléont 34:201–230
Vachard D, Moix P (2011) Late Pennsylvanian to Middle Permian revised algal and foraminiferan biostratigraphy and palaeobiogeography of the Lycian Nappes (SW Turkey): palaeogeographic implications. Rev Micropaléont 54:141–174
Vachard D, Montenat C (1981) Biostratigraphie, micropaléontologie et paléogéographie du Permien de la région de Tezak (Montagnes centrales d’Afghanistan). Palaeontographica B 178:1–88
Vachard D, Gargouri-Razgallah S, Chaouachi MC (1989a) Sur les biohermes à algues solénoporacées et phylloïdes du Permien supérieur de Tunisie (Murghabien du Djebel Tebaga) et sur les incidences de la diagenèse carbonatée sur la systématique algaire. Rev Paléobiol 8:121–141
Vachard D, Martini R, Zaninetti L, Zambetakis-Lekkas A (1993a) Révision micropaléontologique (foraminifères, algues) du Permien inférieur (Sakmarien) et supérieur (Dorashamien) du Mont Beletsi (Attique, Grèce). Boll Soc Paleont Ital 32:89–112
Vachard D, Clift P, Decrouez D (1993b) Une association à Pseudodunbarula (fusulinoïde) du Permien supérieur (Djoulfien) remaniée dans le Jurassique d’Argolide (Péloponnèse, Grèce). Rev Paléobiol 12:217–242
Vachard D, Oviedo A, Flores de Dios A, Malpica R, Brunner P, Guerrero M, Buitron BE (1993c) Barrance d’Olinala (Guerrero): une coupe de référence pour le Permien du Mexique Central; étude préliminaire. Ann Soc Géol Nord 2:153–160
Vachard D, Martini R, Zaninetti L (1995) Le Murgabien à fusulinoïdes des îles d’Hydra, Crète et Mytilène (Permien supérieur de Grèce). Geobios 28:395–406
Vachard D, Fourcade E, Romero JE, Mendez J, Cosillo A, Alonzo M, Requeña J, Azema J, Cros P (1997) Foraminifères et algues du Permien du Guatemala. Geobios 30:745–784
Vachard D, Hauser M, Martini R, Zaninetti L, Matter A, Peters T (2001) New algae and problematica of algal affinity from the Permian of the Aseelah Unit of the Batain Plain (East Oman). Geobios 34:375–404
Vachard D, Hauser M, Martini R, Zaninetti L, Matter A, Peters T (2002) Middle Permian (Midian) foraminiferal assemblages from the Batain Plain (Eastern Oman): their significance to Neotethyan paleogeography. J Foramin Res 32:155–172
Vachard D, Zambetakis-Lekkas A, Skourtsos E, Martini R, Zaninetti L (2003) Foraminifera, algae and carbonate microproblematica from the late Wuchiapingian/Dzhulfian (late Permian) of Peloponnesus (Greece). Riv Ital Paleont Stratigr 109:339–358
Vachard D, Gaillot J, Vaslet D, Le Nindre Y-M (2005) Foraminifers and algae from the Khuff Formation (late Middle Permian-Early Triassic) of central Saudi Arabia. GeoArabia 10(4):137–186
Vachard D, Pille L, Gaillot J (2010) The Palaeozoic foraminifera: their systematics, palaeoecology and responses to the global changes. Rev Micropaléont 53:209–254
Vachard D, Krainer K, Lucas S. (2012) Pennsylvanian (Late Carboniferous) calcareous microfossils from Cedro Peak (New Mexico, USA); Part 1: Algae and microproblematica. Ann Paléont (http://dx.doi.org/10.1016/j.annpal.2012.06.003)
Vaslet D, Le Nindre YM, Vachard D, Broutin J, Crasquin S, Berthelin M, Gaillot J, Halawani M, Al-Husseini M (2005) The Permian-Triassic Khuff Formation of Central Saudi Arabia. GeoArabia 10(4):77–134
Vaziri SH, Yao A, Kuwahara K (2005) Lithofacies and microbiofacies (foraminifers and radiolarians) of the Permian Sequence in the Shalamzar area, Central Alborz, North Iran. J Geosci 48:39–69
Vennin E (2007a) Archaeolithoporella-Shamovella (= Tubiphytes)-bryozoan reef-mound complexes, Russia. In: Vennin E, Aretz M, Boulvain F, Munnecke A (eds) Facies from Palaeozoic reefs and bioaccumulations. Mém Mus Nat Hist Nat 195:295–297
Vennin E (2007b) Artinskian bryozoan-phylloid algal-Shamovella bioherms, Russia. In: Vennin E, Aretz M, Boulvain F, Munnecke A (eds) Facies from Palaeozoic reefs and bioaccumulations. Mém Mus Nat Hist Nat 195:299–301
von Pia J (1912) Neue Studien über die triadischen Siphoneae verticillatae. Beitr Paläont Geol Österr-Ung Orients 25:25–81
von Pia J (1920) Die Siphoneae verticillatae vom Karbon bis zur Kreide. Abh zool-bot Ges Wien 11(2):1–263 (French translation: Editions Technip pp 1–236)
von Pia J (1922) Einige Ergebnisse neuerer Untersuchungen über die Geschichte der Siphoneae verticillatae. Z Indukt Abstamm Vererbungsl 30:63–98
von Pia J (1937) Die wichtigsten Kalkalgen des Jungpaläozoikums und ihre geologische Bedeutung. Compte Rendu du 2e Congrès Avancement Etudes de Stratigraphie du Carbonifère, Heerlen, 1935, vol 2:765–856
Wang KL (1974) Carboniferous Foraminifera. In: Nanjing Inst Geol Paleont (ed) Stratigraphical and paleontological atlas of southwest district. Science Publication House, Beijing, pp 248–256 (in Chinese)
Weidlich O (2002a) Middle and Late Permian reefs-distributional patterns and reservoir potential. In: Kiessling W, Flügel E, Golonka J (eds) Phanerozoic reef patterns. SEPM Spec Publ 72:339–390
Weidlich O (2002b) Permian reefs re-examined: extrinsic control mechanisms of gradual and abrupt changes during 40 my of reef evolution. Geobios, Mém spéc no 24:287–294
Wettstein RR (1901) Handbuch der systematischen Botanik. Vol. 1 Deuticke, Leipzig: 201 p
Wilson MEJ, Vecsei A (2005) The apparent paradox of abundant foramol facies in low latitudes: their environmental significance and effect on platform development. Earth-Sci Rev 69:133–168
The authors appreciate the Payame Noor University of Ardekan, Yazd for providing financial support. Our thanks go to Prof. I. Bucur for his help. Dr. Pedro Cózar (Madrid) reviewed a first draft of the paper; Prof. Dr. Bruno Granier (Brest) revised our manuscript; both are warmly thanked for their constructive criticism.
Author information
Authors and Affiliations
Geology Department, Payame Noor University, 19395-3697, Tehran, Iran
Tahere Parvizi & Koorosh Rashidi
Université Lille 1, UMR CNRS 8217 (Géosystèmes), Bâtiment SN5, 59655, Villeneuve d’Ascq Cédex, France
Parvizi, T., Rashidi, K. & Vachard, D. Middle Permian calcareous algae and microproblematica (Dalan Formation, Dena Mountain, High Zagros, SW Iran).
Facies59, 149–177 (2013). https://doi.org/10.1007/s10347-012-0357-6