
Abstract
The forest fragments surrounding the Boabeng-Fiema Monkey Sanctuary (BFMS) in central Ghana shelter small populations of Colobus vellerosus. Little is known about these populations or the ability of the fragments to support them, despite the fact that these fragments represent potentially important habitat for the colobus in this region. We compared the diet of three groups of C. vellerosus in the fragments to two groups in BFMS. We also examined the differences in plant species composition and food abundance among fragments. The study took place from June to November 2003. Dietary data were collected using scan sampling. Plant species composition and food abundance were evaluated using tree plots and large tree surveys. As in BFMS groups, leaves constituted the highest proportion of the diet of fragment groups, yet the colobus in fragments fed on more lianas than did those in BFMS. Over 50% of all species observed eaten by colobus in the fragments were not consumed in BFMS groups during the same season. Food abundance was similar between fragments and BFMS, although species composition differed. There was no relationship between the density of colobus and the density of food trees or percentage of food species, suggesting that other factors may be influencing the number of colobus present. This study highlights the broad dietary range of C. vellerosus, which may be a factor allowing its survival in these fragments.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Many primate species live in areas where rates of deforestation are among the highest in the world (e.g., Africa, South America; FAO 2000). Deforestation is associated with forest fragmentation. Fragmentation does not only reduce the size of available habitat, but can also create different conditions in the habitat. For instance, the level of disturbance in fragments is often greater than in continuous forests as a result of “edge effects”, which may lead to higher tree mortality or microclimatic changes in the interior of the forest (Lovejoy et al. 1986; Kapos et al. 1997), or increased human activity (Struhsaker et al. 2004). Forest fragments can have a higher proportion of pioneer species and a lower proportion or smaller number of shade tolerant species compared to larger forest blocks (Zhu et al 2004). Overall plant diversity (Estrada and Coates-Estrada 1996) and size of trees (Schwarzkopf and Rylands 1989) can also vary between forest fragments, or disturbed forests, as opposed to continuous forests. These factors may influence food availability for specific species using this habitat (Schwarzkopf and Rylands 1989; Menon and Poirier 1996).
Habitat quality, diet and dietary breadth are important factors influencing the ability of primates to survive in fragments or in disturbed forests (Lovejoy et al. 1986; Menon and Poirier 1996). It seems that folivorous species fare better in forest fragments than frugivorous species (Lovejoy et al. 1986; Johns and Skorupa 1987; Estrada and Coates-Estrada 1996; Gilbert and Setz 2001). However, it is not possible to determine whether a fragment is able to support primates and in what numbers until information is available on the dietary range of a species, and characteristics of a particular fragment such as its size, species composition, and forest structure.
The Boabeng-Fiema Monkey Sanctuary (BFMS) in central Ghana is a 192-ha forest fragment surrounded by even smaller forest fragments, some of which are inhabited by Colobus vellerosus (the Ursine colobus). The colobus apparently travel between some of the fragments. Some of our groups have been observed travelling partly on the ground. We have shown elsewhere (Wong and Sicotte 2006) that fragment size or isolation distance from BFMS were not sufficient to explain colobus density in these fragments. Here, we investigate whether food availability and habitat quality could explain colobus density in this fragmented habitat. We established which tree species were eaten by colobus in BFMS and in forest fragments, documented potential food sources in each fragment, and compared these potential foods to what is actually eaten. We also compared the plant composition and food abundance in fragments with and without C. vellerosus to document differences in habitat quality.
Methods
Study site
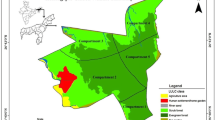
This study took place in five small forest fragments within 2 km of the Boabeng-Fiema Monkey Sanctuary (BFMS) in Central Ghana (7°43′N, 1°42′W): Akrudwa Kuma, Bonte, Busunya, Akrudwa Panyin and Bomini (Fig. 1), from June to November 2003. At the time of the study, colobus monkeys inhabited all fragments except Bomini and Akrudwa Panyin (Table 1; Wong and Sicotte 2006). Some fragments also had small numbers of Cercopithecus campbelli lowei (Akrudwa Kuma=3, Bomini=2). BFMS is a dry semi-deciduous forest fragment (192 ha), separated from any other large forest block by 50 km (Beier et al. 2002). BFMS and surrounding fragments are located in an area characterized by a marked dry season from November to March. Rainfall over 5 years (1985–1990) collected in Nkoranza (20 km south of BFMS) was 1,250 mm (SD:21.1) (Fargey 1992). The long rains last from April–July, the short rains fall in September.

BFMS and surrounding forest fragments. The presence of roads is indicated by the thin lines. There is a village adjacent to each of the small forest fragments (Akrudwa Kuma, Akrudwa Panyin, Busunya, Bomini and Bonte). The villages of Boabeng and Fiema are adjacent to BFMS
The monkeys at BFMS (Colobus vellerosus and Cercopithecus campbelli lowei) are protected at the local level by a traditional taboo which translates into a hunting ban (Fargey 1992; Saj et al. 2005). In 1974, the Ghana Wildlife Division established BFMS as a “Monkey Sanctuary”, a designation that means this area is managed to ensure “perpetuation” of the two monkey species at the site (World Bank 1993). The same level of protection is not extended to the forest (Fargey 1992; Saj et al. 2005). It is possible that the colobus groups found in the smaller fragments originated from BFMS (Wong and Sicotte 2006). Predation pressure is low and likely to be equivalent among the fragments and BFMS.
Data collection
Dietary data from the groups in the forest fragments were collected from August to November 2003 by S.W. Mean rainfall for these 4 months between 2000 and 2002 were respectively: 125 mm (range: 58.7–185.7 mm), 196.7 mm (range: 143.9–244.9 mm), 116.8 mm (range: 75.3–194 mm), 37.3 mm (range: 8.6–66 mm). Three focal groups were selected, one each from Akrudwa Kuma (AU), Bonte (BO) and Busunya (BU). Full day follows were conducted from 0600 to 1800 hours for 3 consecutive days once or twice per month (following Teichroeb et al. 2003 and Saj 2005), for a total of 15 full-day follows for each group. Dietary data were collected for 8–10 h each day. Scan samples were conducted every 15 min (Fashing 2001), and data on the behaviour of the first five individuals spotted in the group were collected (Davies et al. 1999). If the individual was feeding, the plant species and part [leaf (includes leaf bud), fruit, seedpod, flower (includes flower bud), pith, bark, sap, petiole] being consumed were recorded. Five minutes were allowed for finding the individuals and the group was circled between scans to get a random selection of individuals. Dietary data from BFMS were collected on two groups (WW and B) from September to November 2000 and January to August 2001 using focal sampling (Saj 2005).
Characterizations of the size and the species of the trees in each fragment were conducted from July to November 2003 by S.W. and two trained research assistants to estimate species diversity and food abundance (tree plots and large tree survey described below). Tree plots in BFMS were done in September 2001 by T.S. and two trained research assistants. Large tree surveys of the home range of focal groups in BFMS were also conducted, but comparisons with these surveys are not included in this study.
Tree plots
Tree plots were placed at random in all fragments. All plots were placed inside the home range of the focal groups. Species and diameter at breast height (DBH) were recorded for all trees DBH≥10 cm within the plots. The size and number of plots per fragment were as follows: two 50×50 m plots in Bomini and Bonte, three 50×50 m plots in Busunya and Akrudwa Kuma and one 50×50 m plot and one 25×25 m plot in Akrudwa Panyin. This represented a mean of 3.3% per fragment (2.2% in Akrudwa Kuma, 1.5% in Bonte, 1.4% in Busunya, 9.9% in Akrudwa Panyin and 1.6% in Bomini) and a mean of 9.9% of each group’s home range (3.8% in Akrudwa Kuma, 7.7% in Bonte, 18.1% in Busunya). All plots in BFMS were 50×50 m and the number in each group’s home range were as follows: WW=5 plots (10.4% of home range) and B=4 plots (9.1% of home range).
Large tree survey
Trees with DBH≥40 cm were identified to species and DBH was measured. Each tree was mapped with a Trimble GeoExplorer 3.0 GPS receiver. The large tree survey represented a mean of 89.6% of each fragment area (63.9% in Akrudwa Kuma, 95.5% in Bonte, 100% in Busunya, 100% in Akrudwa Panyin and 88.6% in Bomini), and we concentrated measurements in areas of the fragments used by the colobus. As a result of these methods, the habitat quality in Akrudwa Kuma may be over-estimated, since it does not include all the unused areas of the forest (i.e., areas that may lack edible trees or large trees).
Data analysis
The relative proportion of each species consumed was calculated using the percentage of scans spent feeding on each different species. The percentage of scans spent feeding on different plant parts was also calculated and compared to WW and B for August to November. The fragment diet was compared to WW and B groups by calculating: (1) the number of species consumed in fragments but not BFMS, and of those species, which ones were present in BFMS, and (2) the number of species consumed in BFMS but not the fragments, and, of those species, which were present in the fragments. These comparisons were done using WW and B dietary data from the same 4 months of the year (August–November) and also from the complete list of the food species used by both groups. Sorensen’s coefficient of community (Smith 1992) was calculated for all pairs of groups to compare relative similarity in diet. The number of new food species recorded each month for BFMS groups was plotted to evaluate the proportion of dietary elements that 4 months of data collection in the fragments could potentially yield. The ten most commonly eaten tree species for all five groups were also compared.
Spearman rank correlation tests were used to examine if a relationship existed between the frequency of consumption and stem density and the frequency of consumption and basal area for fragment groups, using data from both the tree plots and the large tree survey. Sample size for these correlations was limited by the number of tree species consumed and whether stem density and basal area could be calculated. All tests were two-tailed and were significant at P<0.05. Tests were performed using SPSS 11.5.
To assess the availability of food resources for colobus (i.e., the “habitat quality”), a variety of indices were calculated from the tree plots and the large tree survey. From the tree plots, we calculated: stem density of all trees (food and non-food), Shannon-Wiener index of diversity, density of potential food trees, average DBH of food trees, the percentage of tree species that were potential food trees, and the percentage of potential food tree species consumed by that colobus group. Potential food trees were all trees on which colobus have been recorded feeding in any of the fragments or in the BFMS. The same metrics were calculated for the large tree survey as described above. This was done for both the total area mapped and the home range of the focal groups.
To examine the relationship between the density of colobus and habitat quality, linear regressions were used to examine the relationship between: (1) colobus density and density of all trees, (2) colobus density and density of food trees, and (3) colobus density and percentage of species which were potential food trees. This was done using results from the large tree survey (using only data from the fragments) and the tree plots (using data from each fragments and pooling data from B and WW to get one value for BFMS). The colobus density numbers for BFMS derive from our 2003 census of the whole fragment (Wong and Sicotte 2006).
The top ten species were determined for each fragment from the tree plots. Similarities between BFMS and fragments were examined from the tree plots only, as the BFMS large tree survey was not available for BFMS at the time. The percentages of tree species found in fragments and not BFMS and vice versa were also calculated. For this comparison, species found in fragments used data from both the tree plots and the large tree survey. Data from the 2001 BFMS tree plots (Saj 2005) and the BFMS inventory (Abu-Juam et al. 1996) were used for BFMS.
To examine the level of disturbance in fragments compared to BFMS, comparisons between the proportion of pioneer species and non-pioneer species were made [distinction based on the classifications by Abu-Juam et al. (1996) based on Hawthorne (1990)]. These proportions were calculated for each fragment (using the data from the tree plots and the large tree survey) and BFMS (using data from the 2001 tree plots for B and WW and for this analysis only, data from the BFMS large tree survey).
Results
Diet
A total of 1,754 scans were recorded over 45 days (605 for AK, 555 for BO, 594 for BU). These scans yielded a total of 1,528 individual feeding records for the three focal groups (410 for AU, 583 for BO, and 535 for BU). The diet of fragment groups and BFMS groups (from August to November) consisted primarily of leaves (mature and young) from both tree species and lianas (AU=90.6%, BO=87.5%, BU=70.0%, WW=85.4% and B=95.8%). BU consumed more fruit and seedpods than any other group (Fig. 2). In addition, all fragment groups consumed some insects (<1% in “other” category), which had not been recorded for either WW or B groups. Insects were usually licked but sometimes picked off the bark of trees. Consumption of insects was recognizable because the colobus were obviously picking or licking a moving target. The type of insect is not known. A greater proportion of the diet consisted of lianas in the fragments (AU=26.3%, BO=9.3%, BU=18.0%) compared to BFMS (1.6%; Saj 2005). Fragment groups also consumed two non-native species, Cassia siamea and Azadirachta indica. Cassia siamea was the fourth most consumed species in BU.

Percentage of plant parts consumed by Colobus vellerosus other than leaves in fragments and BFMS. Comparisons made for August to November. Leaves included leaf buds. Flower included flower buds. *Other includes: petioles, bark, sap, pith, soil and insects
A total of 48 plant species were consumed by the combined groups in the fragments (AU=31, BO=22, BU=29) and 35 plant species were consumed in the two BFMS groups (WW=29, B=19) from August to November and 52 plant species (WW=42, B=34) were recorded over 11 months (Fig. 3). A greater proportion of the plants consumed by the colobus in the fragments was not consumed in BFMS when only data from August to November was used (Fig. 3). This was not true when the full year data for BFMS was included (Fig. 3). Of the species not consumed in BFMS, 40.9% (n=9) were found in the home range of WW or B or both. Of the 11 BFMS species (August–November) not consumed in fragments, 6 were found in Akrudwa Kuma, and 2 were found in Busunya. Bonte did not contain any of the unconsumed species.

Comparisons between species consumed by Colobus vellerosus in fragments and BFMS
The rate at which new species were seen to be added to the diet of BFMS groups in relation to time showed that after 7 months of data collection, the number of new species began to plateau. After 4 months of data collection at BFMS, 69.0% (WW) and 55.9% (B) of the species consumed during the full year had been recorded.
Sorensen’s coefficient of community did not show any clear trends in similarities of diet; most values were close to 0.5 (Table 2). Only one species (Adansonia digitata) was a top ten species consumed in all five groups. All groups had at least two species in their top ten species which were consumed exclusively in that group (Table 3).
No relationship existed between frequency of consumption and stem density using data from tree plots (P>0.233). There was a positive association between frequency of consumption and basal area for Akrudwa Kuma (n=16, r s=0.562, P=0.024) but not Bonte (n=10, r s=0.576, P=0.082) or Busunya (n=13, r s=0.503, P=0.079). Using data from the large tree surveys, the frequency of consumption and stem density were positively associated in Akrudwa Kuma (n=21, r s=0.651, P=0.001) and Busunya (n=20, r s=0.491, P=0.028) but not Bonte (n=14, r s=0.207, P=0.477). There was also a positive association between frequency of consumption and basal area for Busunya (n=20, r s=0.801, P < 0.001) but not Akrudwa Kuma (n=21, r s=0.382, P=0.087) or Bonte (n=14, r s=0.341, P=0.233).
Habitat quality
The total number of species found in the tree plots for all fragments combined was 69 (38 for Akrudwa Kuma, 25 for Bonte, 24 for Busunya, 17 for Akrudwa Panyin and 28 for Bomini). The tree plots in BFMS found 69 species (WW=52 and B=38). Of the 69 species found in the fragments combined, 51% were not found in the home ranges of either WW or B in BFMS. The percentage of overlap was 46.4%. All fragments had lower Shannon-Wiener indices than WW and B (Tables 4, 5). Akrudwa Panyin, with no colobus, had the highest mean DBH of food trees and Busunya, with colobus, had the lowest. All fragments had lower density of food trees than WW and B, with fragments not containing colobus having the lowest (Table 4). The percentage of the potential food species was lowest for WW and B compared to the fragments (Table 4). The percentage of food species consumed was highest in Busunya and lowest in WW (Table 4).
A total of 94 large tree species was found during the large tree survey in all the fragments combined (67 in Akrudwa Kuma, 39 in Bonte, 64 in Busunya, 26 in Akrudwa Panyin and 52 in Bomini). In general, the values of the habitat quality indices were higher in the home ranges of the groups than in the entire forests, with the exception of the Shannon-Wiener indices. Stem density in Akrudwa Panyin, with no colobus, was within the values of stem density in the home ranges of the groups. Bomini, also with no colobus, had the lowest stem density. Bomini also had the lowest average DBH of food trees, but DBH of food trees in Akrudwa Panyin was within the values of the home ranges of Akrudwa Kuma and Bonte. The density of food trees (with the exception of Bonte) and the percentage of potential food trees (with the exception of Akrudwa Kuma) in the home ranges of the groups were higher than in the fragments containing no colobus. Of the 16 potential food species available in Bonte’s home range, the group consumed 15 of them (93.8%).
Using data from the tree plots (data from B and WW were pooled to give one value for BFMS), there was no relationship between the density of colobus (Table 1) in the forest and the density of all trees (n=6, r 2=0.297, P=0.264), density of food trees only (n=6, r 2=0.046, P=0.683) or the percentage of species which were potential food trees (n=6, r 2=.003, P=0.914). For the large tree survey (which does not include data from BFMS), there was no relationship between number of colobus in the fragment and density of large trees (n=5, r 2=0.200, P=0.450), density of large food trees (n=5, r 2=0.131, P=0.550), or percentage of species which were potential food species (n=5, r 2=0. 534, P=0. 161)
Considering the ten most abundant tree species found in the trees plots in the fragments and in BFMS, the plots in WW group’s range in BFMS had the least overlap with other plots, with eight distinct top ten species (Table 6). By contrast, Bonte’s plots had only one distinct top ten species (Table 6). Many species found in the fragments were not found in BFMS. For example, of the 108 species found in all fragments combined (using data from the tree plots and large tree survey), 54 species (50%) were not found in WW or B tree plots. When data from the BFMS inventory (Abu-Juam et al. 1996) were included, there were still 27 species (25%) from the fragments which were not found in either WW or B’s home range. Of the 61 species which were identified (8 species are unknowns) in BFMS (WW and B combined), 20 (32.8%) were not found in the fragments.
The percentage of pioneer species in the fragments was higher than BFMS for both the tree plots and the large tree survey (Table 7). The number of non-native species per fragment found during the tree plots was: Akrudwa Kuma=1 (2.6% of total tree species present), Bonte=1 (4.0%), Busunya=1 (4.2%), Akrudwa Panyin=0 and Bomini=1 (3.6%). The number of non-native species found during the large tree survey was Akrudwa Kuma=2 (3.0% of total tree species present), Bonte=0, Busunya=2 (3.1%), Akrudwa Panyin=0 and Bomini=2 (3.8%). The non-native species found most commonly was Cassia siamea.
Discussion
This study documents the diet of C. vellerosus in central Ghana (in BFMS and five surrounding smaller fragments) to determine if differences in habitat quality and presence of potential food could explain differences in colobus densities between these small forests. Since the study in the fragments surrounding BFMS lasted only 4 months, the results should be considered preliminary.
Diet of C. vellerosus in the fragments compared to BFMS
Leaves constituted the highest proportion of plant parts consumed for all fragments and BFMS. The proportions of other plant parts consumed were slightly different. For example, BU consumed less leaves than any other group and more fruit and seedpods. The higher proportion of seedpods in their diet was due to the high abundance of Cassia siamea, which constituted 83% of the seedpods consumed. Insects were eaten in the fragments, which has also been reported for Colobus satanas living in fragments (Tutin 1999). The groups in fragments also consumed a higher proportion of lianas than the BFMS groups, a tendency which has also been found in Callithrix geoffroyi living in fragments (Passami and Rylands 2000). Forest fragments suffer from more “edge effects” and thus contain a higher amount of secondary vegetation (Lovejoy et al. 1986), such as lianas (Zhu et al. 2004). It has been suggested that this secondary vegetation is beneficial to folivores and contributes to their ability to flourish in fragmented forests (Lovejoy et al. 1986). Oates (1977) found that secondary-growth species and forest edge species were common in the habitat of C. guereza in different localities, though species composition changed among sites. Since the abundance of lianas in the fragments and BFMS was not evaluated, it cannot be confirmed whether lianas are more common in the fragments than in BFMS.
During our 4-month study, the number of potential food species was greater than the actual species fed upon for all groups, with the exception of Bonte. The same phenomenon was reported in Colobus guereza in fragments (Onderonk and Chapman 2000). The number of plant species consumed from August to November was within the same range between fragment and BFMS groups. C. vellerosus also consumed non-native species, which has been reported in other species in fragments (Macaca silenus; Bicca-Marques 2003; Singh et al. 2001) and human-altered environments (C. guereza; Grimes and Paterson 2000). This finding suggests that C. vellerosus is able to adjust species in its diet to accommodate the differences in plant composition in the forests. This form of adaptation is supported by Oates (1977) who suggested that while C. guereza is adapted to “marginal” forest habitat, the species is able to exploit vegetation in mature forests.
Alouatta palliata living in forest fragments consumed species more commonly available in those fragments (Estrada et al. 1999). The relationship between frequency of consumption and either stem density or basal area (from the tree plots and the large tree survey) shows mixed results. Only Bonte consistently showed no relationship between these variables, suggesting this group is not consuming species more common in the fragment. The other fragment groups showed at least one positive relationships between consumption and some measure of food abundance.
Habitat quality in the fragments
We would intuitively expect that the food abundance in the fragments without colobus would be lower, if habitat quality was the limiting factor for the presence of colobus. Although the tree plots showed that the density of food trees was lower in the two fragments without colobus, these two fragments were not consistently lower in other habitat quality indices, such as mean DBH of food trees, Shannon-Wiener index, or percentage of food tree species. Furthermore, the lack of relationship between density of food trees and the density of colobus suggests that the absence of C. vellerosus in Akdrudwa Panyin and Bomini cannot be explained by food abundance alone. The absence of colobus in these fragments may be due to a combination of food availability, isolation distance, fragment size, or increased human intolerance in Akrudwa Panyin and Bomini. Alternatively, it may simply be an artifact of the short duration of our study, as groups may be using some fragments seasonally.
The home ranges of the fragment focal groups contained higher food abundance than the rest of their fragment since the DBH, density and percentage of food trees was higher in their home range than in the entire fragment. This demonstrates habitat selectivity, i.e., the behavioural processes of organisms that result in non-random habitat use to maximize survival and fitness (Hutto 1985; Block and Brennan 1993; Jones 2001). Spatial analysis of used and unused/available areas would allow us to identify other important aspects of habitat quality, such as the distribution of large trees, distance between large trees, or distance between food patches, which would lead to a better understanding of foraging and habitat use decisions of C. vellerosus in this area.
Food abundance in the forest fragments containing colobus was comparable to BFMS, despite the differences in plant composition. However, the higher proportion of pioneer species in the fragments suggests a higher level of disturbance than in BFMS. Furthermore, there were more non-native species and species that are grown as commercial food crops in the fragments than in BFMS. Many non-native species are more tolerant of disturbance than native species, and can overwhelm native forests and outcompete indigenous species (Sauer 1998). The level of disturbance in Busunya seemed particularly high. Cassia siamea, which was the most common species in Busunya, is an introduced, pioneer species. Its high abundance in Busunya combined with the low percentage of non-pioneer species suggests that the Busunya fragment may be more disturbed than the other fragments and BFMS. Nevertheless, since pioneer species constituted some of the top ten species in colobus diet, it is unclear whether the higher abundance of pioneer species will inhibit or enhance colobus survival and reproduction in the long-term.
References
Abu-Juam M, Gyakari JN, Ekpe PK (1996) Boabeng-Fiema monkey sanctuary floral inventory. Report prepared for the BFMS Management Committee
Beier P, Van Drielen M, Kankam BO (2002) Avifaunal collapse in West African forest fragments. Cons Biol 16:1097–1111
Bicca-Marques JC (2003) How do howler monkeys cope with habitat fragmentation? In: Marsh LK (ed) Primates in fragments: ecology in conservation. Kluwer Academic Press/Plenum Publishers, New York, pp 283–303
Block WM, Brennan LA (1993) The habitat concept in ornithology: theory and applications. Curr Ornithol 11:35–91
Davies AG, Oates JF, Dasilva GL (1999) Patterns of frugivory in three West African Colobine monkeys. Int J Primatol 20:327–357
Estrada A, Coates-Estrada R (1996) Tropical rain forest fragmentation and wild populations of primates at Los Tuxlas, Mexico. Int J Primatol 8:299–329
Estrada A, Juan-Solano S, Martinez TO, Coates-Estrada R (1999) Feeding and general activity patterns of a howler monkey (Alouatta palliata) troop living in a forest fragment at Los Tuxtlas, Mexico. Am J Primatol 48:167–183
FAO (2000) Global forest resources assessment. Food and Agriculture Organization of the United Nations, Forestry Paper 140, Rome
Fargey PJ (1992) Boabeng-Fiema Monkey Sanctuary—an example of traditional conservation in Ghana. Oryx 26:151–156
Fashing PJ (2001) Feeding ecology of guerezas in the Kakamega Forest, Kenya: the importance of Moraceae fruit in their diet. Int J Primatol 22:579–609
Gilbert KA, Setz EZF (2001) Primates in a fragmented landscape: six species in Central Amazonia. In: Bierregaard RO Jr, Gascon C, Lovejoy TE, Mesquita RCG (eds) Lessons from Amazonia: the ecology and conservation of a fragmented forest. Yale University Press, New Haven, pp 262–270
Grimes K, Paterson JD (2000) Colobus guereza and exotic plant species in the Entebbe Botanical Gardens. Am J Primatol 51(suppl):59–60
Hawthorne W (1990) Field guide to the forest trees of Ghana. Natural Resources Institute, Overseas Development Agency
Hutto RL (1985) Habitat selection by nonbreeding, migratory land birds. In: Cody ML (ed) Habitat selection in birds. Academic, New York pp 455–476
Johns AD, Skopura JP (1987) Responses of rain-forest primates to habitat disturbance: a review. Int J Primatol 8:157–191
Jones J (2001) Habitat selection studies in avian ecology: a critical review. Auk 118:557–562
Kapos V, Wandelli E, Camargo JL, Ganade G (1997) Edge-related changes in environment and plant responses due to forest fragmentation in Central Amazonia. In: Laurance WF, Bierregaard RO Jr (eds) Tropical forest remnants: ecology, management, and conservation of fragmented communities. University of Chicago Press, Chicago, pp 33–44
Lovejoy TE, Bierregaard RO Jr, Rylands AB, Malcolm JR, Quintela CE, Harper LH, Brown KS, Powell AH, Powell GVN, Schubart HOR, Hays MB (1986) Edge and other effects of isolation on Amazon forest fragments. In: Soulé ME (ed) Conservation biology: the science of scarcity and diversity. Sinaur, Sunderland, pp 257–285
Menon S, Poirier FE (1996) Lion-tailed macaques (Macaca silenus) in a disturbed forest fragment : activity patterns and time budget. Int J Primatol 17:969–985
Oates JF (1977) The guereza and its food. In: Clutton-Brock T (ed) Primate ecology: studies of feeding and ranging behaviour in lemurs, monkeys and apes. Academic, New York pp 275–321
Onderdonk DA, Chapman CA (2000) Coping with forest fragmentation: the primates of Kibale National Park, Uganda. Int J Primatol 21:587–611
Passami M, Rylands AB (2000) Feeding behavior of Geoffroy’s marmoset (Callithrix geoffroyi) in an Atlantic forest fragment of South-eastern Brazil. Primates 41:27–38
Saj, TL 2005. Ecological influences on the social organization of Colobus vellerosus at Boabeng-Fiema, Ghana. PhD thesis, University of Calgary, Canada
Saj TL Teichroeb JA, Sicotte P (2005) The population status of Colobus vellerosus at Boabeng-Fiema sacred grove, Ghana. In: Paterson JD, Wallis J (eds) Commensalism and conflict: the Human-Primate Interface. American Society of Primatology, pp 350–375
Sauer LJ (1998) The once and future forest. Island Press, Washington, D.C
Schwarzkopf L, Rylands AB (1989) Primate species richness in relation to habitat structure in Amazonian rainforest fragments. Biol Cons 48:1–12
Singh M, Kumara HN, Kumar MA, Sharma AK (2001) Behavioural responses of lion-tailed macaques (Macaca silenus) to a changing habitat in a tropical rain forest fragment in the Western Ghats, India. Folia Primatol 72:278–291
Smith RL (1992) Elements of ecology. Harper Collins, New York, pp 305
Struhsaker TT, Marshall AR, Detwiler K, Siex K, Ehardt C, Lisbjerg DD, Butynski TM (2004) Demographic variation among Udzungwa Red Colobus in relation to gross ecological and sociological parameters. Int J Primatol 25:615–658
Teichroeb JA, Saj TL, Paterson JD, Sicotte P (2003) Effect of group size on activity budgets of Colobus vellerosus in Ghana. Int J Primatol 24:743–758
Tutin CEG (1999) Fragmented living: behavioural ecology of primates in a forest fragment in Lopé Reserve, Gabon. Primates 40:249–265
Wong S, Sicotte P (2006) The population size and density of Colobus vellerosus at the Boabeng-Fiema Monkey Sanctuary and surrounding forest fragments, Ghana. Am J Primatol 68:465–476
World Bank (1993) Ecologically sensitive sites in Africa, Volume IV: West Africa. The World Bank, Washington, DC
Zhu H, Xu ZF, Wang H, Li BG (2004) Tropical rain forest fragmentation and its ecological and species diveristy changes in southern Yunnan. Biodivers Conserv 13:1355–1372
Acknowledgements
We thank the Ghana Wildlife Division, the Management Committee of the Boabeng-Fiema Monkey Sanctuary and the chiefs and residents of Akrudwa Kuma, Akrudwa Panyin, Bonte, Busunya and Bomini for giving us permission to conduct research in these forests. Many thanks to Moses Ampofo (traditional herbalist) for his assistance in tree identification in the field. We are grateful to Adomako Seth and Effah Boa-Amponsem for their assistance with ecological surveys and to Charles Kodom, Constance Serwaa, and Afia Faustine Boahen for their assistance with behavioural data collection. Robert Ronconi and Mark Wong also provided help in the field. Thank you to Dr. Colleen Cassidy St. Clair (University of Alberta) for use of equipment. Patrick Ekpe (University of Ghana) provided assistance with plant species identification. The majority of the funding for this research was provided by the Wildlife Conservation Society. The following organizations/institutions also provided funding: Primate Conservation Inc., Natural Science & Engineering Research Council, and the University of Calgary. This research has been approved by the Animal Care Committee of the University of Calgary.
Author information
Authors and Affiliations
Corresponding author
About this article
Cite this article
Wong, S.N.P., Saj, T.L. & Sicotte, P. Comparison of habitat quality and diet of Colobus vellerosus in forest fragments in Ghana. Primates 47, 365–373 (2006). https://doi.org/10.1007/s10329-006-0186-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10329-006-0186-8