
Abstract
This review considers the factors involved in the regulation of feeding and metabolism in response to food deprivation using Caenorhabditis elegans as a model organism. Some of the sensory neurons and interneurons involved in food intake are described, together with an overview of pharyngeal pumping. A number of chemical transmitters control feeding in C. elegans including 5-hydroxytryptamine (5-HT, serotonin), acetylcholine, glutamate, dopamine, octopamine, and tyramine. The roles of these transmitters are modified by neuropeptides, including FMRFamide-like peptides (FLPs), neuropeptide-like protein (NLPs), and insulin-like peptides. The precise effects of many of these neuropeptides have yet to be elucidated but increasingly they are being shown to play a role in feeding and metabolism in C. elegans. The regulation of fat stores is complex and appears to involve the expression of a large number of genes, many with mammalian homologues, suggesting that fat regulatory signalling is conserved across phyla. Finally, a brief comparison is made between C. elegans and mammals where for both, despite their evolutionary distance, classical transmitters and neuropeptides have anorectic or orexigenic properties. Thus, there is a rationale to support the argument that an understanding of the molecular and genetic basis of feeding and fat regulation in C. elegans may contribute to efforts aimed at the identification of targets for the treatment of conditions associated with abnormal metabolism and obesity.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
There has been interest in feeding and food deprivation in C. elegans for over 30 years (Avery and Horvitz 1990). However, with the increasing occurrence of obesity in Homo sapiens as a consequence of Western diets (O’Rahilly and Farooqi 2008), there is mounting interest in using this model genetic animal to delineate molecular mechanisms of food intake, metabolism, and metabolite storage, particularly in relation to fat. Increasingly evidence suggest that the genes involved in storage and metabolism are similar in humans and C. elegans, with the latter having orthologues of a number of the relevant human genes. In this review, the regulation of feeding, food metabolism, and its storage in response to food deprivation will be considered in C. elegans, together with a brief comparison with the situation in mammals. In particular, there is a focus on the neurohormonal and neuronal pathways and the behavourial outputs that drive food intake in the absence of calorific requirement.
Feeding in C. elegans is closely linked with the regulation of pharyngeal pumping. In addition, the control of body wall muscle and the neuromuscular junctions that drive locomotion are important structures as they enable the organism to express the changes in patterns of activity that allow it to re-locate and move towards food sources. Therefore, the regulation of both systems will be described. The literature regarding the factors that induce dauer formation or regulate ageing in C. elegans is extensive but although both factors are closely associated with the availability and intake of food, neither will be considered in this review. The reader is also referred to two recent reviews which complement the current review and deal with aspects of fat storage and its regulation (Mullaney and Ashrafi 2009; Watts 2009). Tables 1 and 2 have been included to assist the reader in the identification of the main neurons and genes described in the text.
Neural networks
The neural networks involved in the regulation of food intake in C. elegans are probably under the control of a range of stimuli including chemical signals emanating from food sources in the environment (Chalasani et al. 2007), and mechanical signals produced by the physical presence of food, either externally or internally. Furthermore, but less well studied, are the chemosensory pathways and receptors that must be involved in detecting the metabolic and nutritional state of C. elegans. These sensory inputs that are activated by either external or internal signals are clearly integrated, and changes in such stimuli result in whether the animal adopts foraging behaviour. This behaviour involves a distinctive repertoire of forward movement, reversals, and turning.
A number of neurons have been implicated in behaviour associated with feeding, including the amphid wing sensory neurons, AWC, and the amphid interneurons, AIY (Gray et al. 2005). AWCs are activated following the removal of an odour which in turn stimulates the turning behaviour in the animal which underpins local area search behaviour. This is tightly temporally regulated and is observed in the first 15 min following removal from food while the animals continue to search for food in a restricted immediate area. AWCs release the neuropeptide NLP-1 that activates AIA interneurons, probably through the predicted G protein-coupled receptor for neuropeptides, NPR-11, and AIAs induce feedback inhibition onto AWCs via INS-1 (an insulin-related neuropeptide) (Chalansani et al. 2010). These authors found that INS-1 is required for the biological activity of NLP-1. AWCs also synthesise and release glutamate as a transmitter which is involved in local area search behaviour (Chalasani et al. 2007). Glutamate released from AWCs acts on a number of follower interneurons, including AIYs, Fig. 1. AIYs synthesise and release both acetylcholine and FLP-18s (FMRFamide-like neuropeptides) (Altun-Gultekin et al. 2001; Wenick and Hobert 2004). These cells express about 25% of all C. elegans G protein-coupled neuropeptide receptors, at least 6 ion channels, including a glutamate-gated chloride channel (GLC-3), SER-2 (tyramine) receptors and metabotropic receptors for acetylcholine and glutamate (Wenick and Hobert 2004). These findings support the observations of White et al. (1986) that AIYs receive many synaptic inputs and in addition are probably activated non-synaptically by neurohormones. AWCs also synapse onto the amphid interneurons, AIB, which express glr-1, a glutamate-gated cation channel. When activated by odour removal, AWCs activate AIBs via GLR-1 (an AMPA-like receptor) but inhibit AIYs via GLC-3 (glutamate-gated chloride channel/receptor). Both interneurons play a role in local search behaviour (Chalasani et al. 2007). This behaviour is manifest as repetitive bursts of reversing and turning, but these events are inhibited after about 15 min when the animals begin to extend their food search to explore other areas.

The pathway connecting sensory neurons to ventral motoneurons. The amphid sensory neuron, AWC, is activated when an odour is removed while its basal activity is reduced following application of an odour. AWC synapses onto primary interneurons (neurons with direct input from sensory neurons) including AIY which in turn synapse onto secondary interneurons (neurons only connected to other interneurons) including RIB. RIB synapses with command interneurons including AVB and AVA, and these command interneurons regulate ventral motoneurons to induce forward and backward movement (after Tsalik and Hobert 2003)
The response of an animal to being placed in an environment without food is related to its fed state. Thus, when well-fed animals are placed in an environment without food, they show a relatively high frequency of reversals as they exert behaviour which optimises the possibility of finding food in the immediate area. Conversely, when food-deprived animals are placed in an environment without food, their reversal rate is much less frequent. These animals may undertake straight forward runs of movement to rapidly escape the unfavourable environment (Tsalik and Hobert 2003). In addition, there are mechanosensory neurons which respond to food texture, viz, ADEs, PDEs and CEPs (Sawin et al. 2000). When well-fed animals are transferred to a bacterial lawn, they move more slowly compared to animals transferred to a plate lacking bacteria. This is termed “the basal slowing response” and is due to the ADE, PDE, and CEP dopaminergic neurons sensing the mechanosensory stimulus of the bacteria. ADEs synapse onto a number of neurons including the command interneurons, AVA. PDEs synapse onto DVA which is activated by body stretch to fine-tune motor activity (Li et al. 2006).
Feeding
Feeding has been extensively studied in C. elegans (Seymour et al. 1983; Avery 1993a) and depends on pharyngeal pumping activity which has been reviewed in depth (Franks et al. 2006). In a detailed investigation, Avery identified 35 genes involved in feeding including 7 phm (pharyngeal muscle) genes which when mutated led to abnormal muscle contraction. Furthermore, this work identified 26 eat genes in which mutants had a disrupted pharyngeal muscle movement but were still able to sustain relatively vigorous pumping. Avery considered that the eat genes affected either the pharyngeal nervous system function or/and muscle functions that regulate contraction. Pharyngeal muscle function is dependent on EXP-2, a voltage-activated potassium channel, which is responsible for rapid repolarization and which supports muscle relaxation and the fast closing of the lumen necessary to force food into the intestine (David et al. 1999). Pumping is regulated by an intrinsic pharyngeal nervous system consisting of 20 neurons which is linked to the rest of the nervous system through a single pair of neurons, RIP (pharyngeal ring interneurons) neurons (Albertson and Thompson 1976; White et al. 1986). In the absence of pharyngeal motoneurons, pharyngeal pumping can continue albeit slow and uncoordinated (Avery 2010; Avery and Horvitz 1989). In addition, pharyngeal pumping can be influenced by neurohormones released from neurons extrinsic to the pharyngeal nervous system. However, it should be noted that the pharynx is separated from the pseudocoelom by its own basal lamina for which the molecular structure has been described in detail (www.wormatlas.org), but it has been proposed that C. elegans lacks a functional blood–brain barrier (Nass et al., 2008), and so neurohormones could pass from the pseudocoelomic fluid to the pharynx.
The phenomenon of starvation in C. elegans was first investigated by Croll and Smith (1978) and subsequently by Avery and Horvitz (1990). Croll and Smith failed to find an effect of food deprivation (4 h fasting) on feeding behaviour measured over a 25-min period at minute intervals though they found that bacteria stimulated pumping. In their study, Avery and Horvitz defined starvation as being deprived of food for 2 h and found that starved animals responded to much lower concentrations of bacteria than well-fed C. elegans. These authors found a clear increase in pumping rate after 4 h of starvation. Starved C elegans all responded to bacteria with rapid pumping, while only 50% of the well-fed animals pumped rapidly. The quality of food also influences pumping with poor-quality food increasing the rate, while high-quality food decreases pumping rate (You et al. 2006). Starvation would therefore seem to induce a signal which can change the feeding behaviour of the animal as assayed by pharyngeal pumping. One of these factors would appear to be MAPK (mitogen-activated protein kinase) which is often engaged by stress responses including those associated with starvation. This pivotal regulator of cell signalling is susceptible to receptor activation and in C. elegans is stimulated following the activation of muscarinic receptors in the pharynx following starvation (You et al. 2006). The muscarinic agonist, arecoline, stimulated pharyngeal muscle MAPK in a way that was inhibited by the antagonist atropine in gpb-2 mutants that are sensitive to starvation due to a defect in their pharyngeal muscle. In contrast, neither nicotine nor 5-HT activated MAPK. In another study to investigate molecules that modify the starvation signal Kang and Avery (2009) found that leucine, glutamine, valine, and alanine could suppress the starvation-induced death of gpb-2 mutants. The metabotropic glutamate receptors MGL-1 and MGL-2 were involved in this effect with MGL-1 located on AIY neurons and MGL-2 receptors located on AIB neurons. While leucine suppressed starvation-induced stress resistance and the extension of life-span in wild-type animals, mutations of mgl-1 and mgl-2 prevented these effects of leucine. Kang and Avery also found evidence that AIYs and AIBs might be able to directly detect amino acids rather than through a sensory neuron, such as AWC, and that amino acids might act as anti-hunger signals in wild-type C. elegans. They found evidence to suggest that food or amino acids activate AIYs which then inhibit the starvation response, while food or amino acids inhibit AIBs which activate the response. There are no direct synaptic links between AIYs and AIBs with pharyngeal muscle, and so it is likely that neurohormonal effectors act from these neurons onto the muscle with various classes of monoamines and neuropeptides having a possible role. In this regard, AIYs express the flp-18 neuropeptide gene, while it is not known which peptide genes are expressed in AIBs. It has also been shown that AIY neurons are involved in the extension of roaming periods and are essential for efficient food seeking (Shtonda and Avery 2006). IP3 signalling through activation of IP3Rs in the terminal bulb and isthmus of pharynx and in the nervous system has also been shown to play a role in the food-induced stimulation of pharyngeal pumping in C. elegans (Walker et al. 2002).
Transmitters regulating feeding
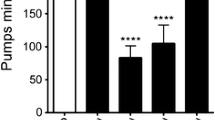
A number of amines have been implicated in the regulation of feeding with the best evidence for 5-hydroxytryptamine (5-HT, serotonin) (Horvitz et al. 1982; Avery and Horvitz 1990). Croll (1975) was first to propose a role for 5-HT in the coordination of nematode behavioural activities, including eggs laid, vaginal/vulval activity, and pharyngeal bulb contractions. Since his pioneering work, a number of papers have been published demonstrating that 5-HT can increase the resting pumping level from around 40 pumps per minute to a maximum of around 250 pumps per minute (Avery and Horvitz 1990; Niacaris and Avery 2003). It has also been shown that a reduction in endogenous levels of 5-HT through the removal or reduction in the activity of tryptophan hydroxylase, the rate limiting enzyme for the synthesis of 5-HT, results in a low pumping rate in the presence of bacteria and a raised level of stored fat (Sze et al. 2000). However, in the absence of food, tryptophan hydroxylase–deficient mutants, tph-1, pump at rates comparable to wild-type animals (Sze et al. 2000). Thus, while 5-HT is required for the up-regulation of pumping in the presence of food, it is not necessary for basal pumping rate. On closer analysis, Hobson et al. (2006) showed that tph-1 mutants can pump at rates similar to wild-type, but their pumping behaviour is erratic and they fail to sustain fast pumping. 5-HT is present in pharyngeal neurons, with the main 5-HT-containing cells being the paired NSM neurons (Horvitz et al. 1982) that synapse onto pm5 muscles of the isthmus (Albertson and Thompson 1976). It is likely that I5 whose axons extend forward to the terminal bulb and terminate in the nerve ring also contains 5-HT (Sawin et al. 2000). The observation that 5-HT cannot stimulate pumping in ser-7 mutants but that bacteria can provides evidence that 5-HT is not the only transmitter relaying physiological stimuli associated with the feeding state (Hobson et al. 2006). Interestingly, these authors found that pharyngeal pumping in ser-7 mutants was inhibited by 5-HT while pumping was stimulated by bacteria, suggesting a feedback system activated through a different 5-HT receptor. SER-1 5-HT receptors are widely distributed on pharyngeal neurons (M4, NSMs, MCs) and muscle, interneurons, motoneurons, and touch receptor neurons (Carre-Pierrat et al. 2006; Hamdan et al. 1999; Hobson et al. 2003; Tsalik et al. 2003; Xiao et al. 2006). Since the cholinergic MC neurons have SER-7 receptors, it is possible that 5-HT activation of these receptors also plays an indirect physiological role in regulating pumping. 5-HT can also activate neuropeptide signalling pathways e.g. it stimulates DAF-2 insulin-like signalling in C. elegans (Sze et al. 2000).
In addition to 5-HT, both acetylcholine and glutamate are present in identified neurons in the pharyngeal nervous system. At least 8 neurons, many of them motoneurons, contain acetylcholine and 5 neurons contain glutamate, including both I5 and the NSM neurons (Rand et al. 2000; Chiang et al. 2006; Lee et al. 1999). Since acetylcholine is excitatory on pharyngeal muscle, it probably plays a role in activating pharyngeal pumping (Avery and Horvitz 1989). This is particularly true for the MC and M4 neurons which innervate the metacarpal and isthmus muscles, respectively (Albertson and Thompson 1976). Both nicotinic and muscarinic receptors are present on pharyngeal muscle. EAT-2, EAT-18, and ACR-7 are examples of nicotinic receptors, while GAR-3 is an example of a muscarinic receptor on pharyngeal muscle (McKay et al. 2004; Steger and Avery 2004). Inhibitory junctional potentials (ijps) can be recorded from pharyngeal muscle during the period of muscle contraction and shorten the duration of each syncytial muscle contraction (Avery 1993b; Li et al. 1997). These ijps are due to activation of the glutamate-containing M3 neurons which therefore have a role in pump activity (Dent et al. 1997; Li et al. 1997). Following ablation of M3, these ijps are absent (Li et al. 1997). At least some of the sensory neurons in C. elegans are glutamatergic, including the mechanosensory (ASH) and chemosensory neurons (ASK) (Lee et al. 1999) and feeding status and 5-HT can influence the responses of these cells which activate follower command interneurons through glutamatergic synapses (Chao et al. 2004). Cation selective non-NMDA receptors are expressed in many command interneurons, including AVAs, AVEs, AVDs, AVBs, and PVCs (Hart et al. 1995; Maricq et al. 1995; Brockie et al. 2001). Thus, both 5-HT and glutamate contribute to changes in the activity of circuits which can modify pharyngeal pumping. In addition, a number of pharyngeal neurons express ionotropic glutamate receptors, viz, I1-3, I6, MI, M3-4, MCs, and NSMs (Brockie et al. 2001), indicating a key role for glutamate in the regulation of pharyngeal pumping. It is probable that changes in levels of 5-HT, acetylcholine, glutamate, or neuropeptides, due to changes in metabolic state, can influence feeding activity in C. elegans.
Octopamine and tyramine can also be added to the list of transmitters influencing feeding since they both inhibit pharyngeal activity (Rogers et al. 2001). Epinastine is a selective antagonist of the octopamine-mediated inhibition (Packham et al. 2010). Only one pair of neurons contains tyrosine decarboxylase but not tyramine-β-hydroxylase, viz, RIM motoneurones, and it is likely that these neurons release tyramine as a transmitter or neurohormone (Alkema et al. 2005). RIM has a reciprocal gap junction with AIYs. The only pair of neurons containing both tyrosine decarboxylase and tyramine-β-hydroxylase is RIC, and these neurons are likely to release tyramine and octopamine (Alkema et al. 2005). However, neither amine has been identified in neurons in the pharyngeal nervous system but both could act as neurohormones as they impact on pumping and their receptors are expressed in the pharynx. These transmitters are often discussed as the parallel of the adrenergic system. Dopamine, octopamine, and glutamate have all been implicated in the responses associated with the presence or absence of food (Hills et al. 2004; Suo et al. 2006, 2009a, b). In the presence of food, C. elegans exhibits area-restricted search (ARS) behaviour associated with the release of dopamine from certain sensory neurons, viz, CEPs, PDEs, and ADEs. Dopamine can then act on two follower cell circuits. Dopamine can act on command interneurons, viz, AVAs, AVBs, AVDs and PVCs, which also express GLR-1 and GLR-2, both AMPA-like glutamate receptors. In turn, these command interneurons synapse onto motoneurons which regulate forward and backward movement. In the presence of food, dopamine can inhibit the octopamine-containing RIC neurons through activation of DOP-3 receptors which results in ARS behaviour. In the absence of food, no dopamine is released and there is no ARS behaviour, but octopamine is now released from RIC neurons. This octopamine can activate cholinergic SIA neurons through SER-3 receptors. This leads to CREB activation in the SIA neurons (Suo et al. 2009a). In the presence of food, dopamine acts on DOP-2 and DOP-3 receptors located on SIA neurons to inhibit CREB activation through the inhibition of Gq signalling by activating GOA-1 (Goα). The interaction between DAF-7, DAF-3, octopamine, tyramine, and glutamate in pharyngeal pumping and the accumulation of fat (Greer et al. 2008) will be described in the section on the regulation of fat stores.
Neuropeptides regulating feeding
In a study using egl-3 and egl-21 mutants which have a severely depleted profile of neuropeptides, there was a decrease in the fat content of the intestine, supporting a role for neuropeptides in feeding and/or fat storage (Husson et al. 2006, 2007).
It is likely that most if not all the pharyngeal neurons contain one or more neuropeptides, in addition to small molecule neurotransmitters. These are either FLPs (FMRFamide-like peptides) or NLPs (neuropeptide-like peptides) (Kim and Li 2004; Li 2005; Li and Kim 2008; Nathoo et al. 2001). For example, these authors have identified a number of FLPs in M2, M3, M4, M5, I1, I2, I4, I5, I6, NSM, and MC neurons. FLPs and NLPs also act on pharyngeal muscle to either excite or inhibit pumping (Papaioannou et al. 2005, 2008; Rogers et al. 2001).
One of the key interneurons involved in the response following an olfactory stimulus is the primary interneuron, AIY. As already described, this neuron expresses a large number of G-protein-coupled receptor genes associated with neuropeptides including a tachykinin-like receptor, somatostatin-like receptor, bombesin-like receptor, CCK-like receptor, and an FMRFamide-like receptor (Wenick and Hobert 2004). In addition, AIYs synthesize and release FLP-18s onto follower neurons or as neurohormones and the roles of these FLPs have been investigated by Cohen et al. (2009). Four neuropeptide receptors mutants (npr) are associated with changes in intestinal fat, viz, npr-2, npr-4, npr-5, and npr-7, all having raised levels in intestinal fat compared to wild-type C. elegans. This would suggest that neuropeptides acting on these receptors are involved in the storage and metabolism of fat. Cohen et al. tried to identify the neuropeptides by injecting cRNA corresponding to each NPR into Xenopus oocytes and then applying neuropeptides to determine which activated the receptors. Since flp-18 is expressed in AIY, they first tested FLP-18 peptides. There are six FLP-18s, all with a C terminal -PGVLRFamide and with a total of 8–12 amino acids (Kim and Li 2004, 2008). FLP-18 peptides activated both NPR-4 and NPR-5 receptors, with NPR-5 responding more strongly than NPR-4. FLP-21 also activated both receptors, but FLP-23 and FLP-32 had no effect. These authors found that npr-4 was expressed in a number of sites including AVA and RIV neurons and parts of the intestine while npr-5 was expressed in a number of amphid neurons, viz, ADF, ASE, ASG, ASI, ASJ, ASK, AWA and AWB and in AIA and AUA interneurons. npr-5 was also expressed in head, neck, and body wall muscles. Cohen et al. (2009) found that flp-18 mutants were defective in finding near-threshold odour sources and that this property could be restored if flp-18 was specifically expressed in AIYs. This confirms that AIYs play a role in behaviours associated with odour detection. These authors also found evidence that FLP-18s released from AIYs activate a switch in behaviour from local search for food to dispersal after prolonged food withdrawal, i.e., playing a role in reversal behaviour. FLP-18s activated NPR-4 receptors on RIV to regulate reversals and on AVA to regulate turning behaviour, both events associated with foraging. They concluded that FLP-18 peptides released from AIY and RIG interneurons act on a group of sensory neurons and on intestinal muscle to regulate accumulation of intestinal fat. Both FLP-18 and FLP-21 peptides acting on NPR-1 have been implicated in social feeding in C. elegans with FLP-21 peptides acting in vivo to inhibit social feeding and on release could activate NPR-1 on AQR, PQR, and URX body cavity neurons (Rogers et al. 2003) as these authors showed that flp-21 is expressed in the intestine. NPR-1 has been cloned and sequenced and shown to be related to the mammalian NPY receptor (de Bono and Bargmann 1998) and is expressed on sensory, motor, and interneurons in C. elegans (Coates and de Bono 2002).
Two cholecystokinin (CCK)-gastrin-like peptides have been identified in C. elegans, viz, DYRPLQFamide (NLP-12a) and DGYRPLQFamide (NLP-12b), together with their receptors, CeCKR-2a and CeCKR-2b, respectively (Janssen et al. 2008). nlp-12 is expressed in a ring interneuron, DVA, and all six coelomocytes. DVA is associated with the integration of mechanosensory information and provides input to the anterior and posterior touch circuits (Wicks et al. 1996). The pharyngeal pumping rate of nlp-12 and ckr-2 mutants was the same as wild type, but both mutants showed an increase in fat storage. crk-2 mutants but not nlp-12 mutants had a reduced amylase secretion. Taken together these effects would indicate a reduced metabolism of the fat and carbohydrate stores. These results provide evidence for a role for NLP-12 in the regulation of fat storage in C. elegans and complement the findings that CCKs in mammals are involved in satiety to regulate food intake (Dufresne et al. 2006). They reinforce an earlier finding where tripeptidyl peptidase II (TPPII), which degrades cholecystokinin-8 in mammals and is expressed in fat-storing cells in C. elegans, regulates fat storage in C. elegans (McKay et al. 2007). RNAi of TPPII decreased fat stores in C. elegans.
As already described, AWCs play a key role in odour detection and subsequent locomotor activity. In addition to glutamate, AWCs synthesize NLP-1s. nlp-1 encodes three NLP-1-like neuropeptides with a common C terminal of -FRMSFa (Li and Kim 2008). NLP-1s act on AIAs through NPR-11 to inhibit AIAs (Chalansani et al. 2010). AIAs in turn provide feedback inhibition either directly or indirectly through the release of INS-1. An NPR which has been implicated in local search behaviour is NPR-9, an allatostatin/galanin-like receptor (Bendena et al. 2008). npr-9 mutants showed a lack of roaming behaviour on food, that is, where animals progress in a forward direction which is briefly interrupted by backward movement to change the direction of forward movement. When not on food, npr-9 mutants showed shortened forward movement with high angle turns equivalent to wild-type animals (Bendena et al. 2008). npr-9 mutants accumulate fat in their intestines and the gene is only expressed in the AIB interneurons. Bendena et al. (2008) propose that NPR-9 may act antagonistically to GLR-1 (also present on AIB) to regulate foraging. Neuropeptides, including the one which activates NPR-9, may also inhibit release of glutamate (Mellem et al. 2002). Neuropeptides have been shown to inhibit synaptic activity of glutamate neurons in mammals (Gao and van den Pol 2001). Bendena et al. (2008) consider that NLP-5 and NLP-6 are the most likely ligands to activate NPR-9 since their peptides resemble allatostatin-like peptides. There are three NLP-5s with a C terminal of -TLGGMGL/Famide and four NPL-6s with a C terminal of -F/MGF/Lamide (Li 2005).
Further, evidence for the possible role of neuropeptides comes from the work of Lee and Ashrafi (2008). These authors found that UNC-31, which has been implicated in neuropeptide release (Speese et al. 2007), acts in a group of head sensory neurons, viz, AWAs, ADFs, ADLs, and ASHs to modulate the degree of starvation survival in L1 C. elegans. The mechanism is through the action of TRPV (transient receptor potential vanilloid) channels located on the cilia of these sensory neurons. The TRPV channel is involved in sensing nutrient availability which may in turn influence the survival of L1 larva in starvation conditions. This activation results in the release of dense core vesicle material which acts on DAF-2 receptors. The material could be INS-1 since ins-1 is expressed in AWA, ADF, and ASH neurons though evidence suggests that this peptide may act as an antagonist at DAF-2 receptors (Li and Kim 2008; Pierce et al. 2001). In the absence of UNC-31, the survival in conditions of starvation is enhanced. It has also been shown that unc-31 mutants pump constitutively, that is, they pump in the absence of food, even when well fed (Avery et al. 1993). This pumping might be due to the reduction in or absence of the release of one or more neuropeptides.
Regulation of fat stores
Fats in C. elegans are stored mainly in the intestine as triglycerides which are formed by the esterification of fatty acids with glycerol. Fatty acids are obtained either from bacteria or synthesized de novo, and both systems play a role in the accumulation of C16 and C18 mono- and polyunsaturated fatty acids (MUFAs and PUFAs, respectively) (Watts 2009). Monoethyl branched chain fatty acids (mmBCFAs), C15 iso and C17 iso, are derived almost entirely from de novo synthesis since they do not occur in bacteria (Watts 2009). MUFAs synthesized by Δ9 desaturases (FAT-5,6,7) are very common in triglycerides and phospholipids. MUFAs are essential for life in C. elegans since the triple mutant, fat-5;fat-6;fat-7, is not viable (Brock et al. 2006). Thus, the correct fatty acid composition of C. elegans must be maintained.
The storage of fat in C. elegans can be regulated by several factors including 5-HT, insulin-like peptides and other growth factor like receptors ligands including DAF-7/TGFβ. The regulation of fat by 5-HT appears to be a distinct metabolic response activated by the nervous system and separate from the 5-HT system which regulates feeding (Srinivasan 2008). While 5-HT activates feeding rate (Horvitz et al. 1982) it lowers the level of stored fat. Which 5-HT receptors are involved in these two events? ser-1 and ser-7 mutants showed wild-type fat levels, but their feeding rate did not increase when exposed to 5-HT. In contrast, in the presence of 5-HT, the fat levels in mod-1 and ser-6 mutants were higher than wild type, while their feeding rates were raised. The double mutant, mod-1;ser-6 had even higher fat levels than the single mutants. From this, it can be concluded that the 5-HT receptors involved in fat storage are MOD-1 (a 5-HT-gated chloride channel) and SER-6 (Srinivasan 2008). ser-6 is expressed in a subset of head and tail neurons (Srinivasan 2008) while mod-1 is also expressed in several head and tail neurons and in the ventral cord (Ranganathan et al. 2000). There is no evidence for mod-1 expression in any muscle, suggesting it functions at the neuronal level. Srinivasan (2008) also found evidence that mobilization of fatty acids by 5-HT and the prevention of their breakdown at the same time by inactivation of genes regulating β-oxidation results in a homeostatic reduction in feeding rate. This neural fat regulating system which is independent of feeding can cause fat store reduction by stimulating lipid oxidation through activation of β-oxidation genes. Srinivasan (2008) also found evidence that the insulin pathway is not required for the reduction in fat levels by 5-HT since daf-16 mutants still showed a fall in fat levels following 5-HT treatment. However, the raised fat levels of daf-2 mutants were reduced by 5-HT treatment.
A number of receptors and ligands have been implicated in fat storage in C. elegans, including the insulin-like receptor DAF-2 and DAF-7 for which the receptors are DAF-1 and 4 (Ashrafi et al. 2003; Kimura et al. 1997; Mak et al. 2006; Watts 2008). DAF-28, INS-1, and INS-7 have all been proposed as ligands for DAF-2 (Li et al. 2003; www.wormatlas.org). In a key paper, Ashrafi et al. (2003) used RNAi to demonstrate that 305 genes when inactivated resulted in a fall in stored fat in C. elegans, while 112 genes when inactivated resulted in a rise in stored fat. This is a surprisingly high number and it is interesting that RNAi for these genes also often leads to a reduction in fat levels in daf-2, tph-1, and tub-1 mutants which have higher levels of fat than wild type. There was a particularly good correlation between wild-type and daf-2 mutants while a number of the RNAi clones failed to reduce fat levels in tph-1 and tub-1 mutants. However, there was a strong correlation between whether these RNAi clones could or could not reduce fat storage in tph-1 and tub-1 mutants. Ashrafi et al. (2003) concluded that the fat-increasing signals due to tph-1 and tub-1 mutations have at least one common pathway which is distinct from the pathway that raises fat storage in daf-2 mutants. This is linked to the observation that both 5-HT and TUB-1 signalling occur in neurons, while fat regulation associated with DAF-2 also occurs in non-nervous tissue (Apfeld and Kenyon 1999; Wolkow et al. 2000). Some of the genes described by Ashrafi et al. (2003) are known to alter fat metabolism, for example, genes encoding enzymatic components of membrane lipid biosynthesis, β-oxidation of fat and components of sterol metabolism. RNAi of a glutamate (C43H6.9) receptor and a somatostatin-like receptor (F56B6.5) resulted in raised fat levels. These authors found that over 50% of the fat regulating genes in C. elegans had mammalian homologues that had not been implicated in fat storage regulation. tub-1 is an orthologue for the mammalian tubby gene (Liu et al. 1999) and can regulate fat storage independent of daf-16 (Mukhopadhyay et al. 2005). These authors found that TUB-1 interacts with a RabGTPase-activating protein, RBG-3, to regulate fat storage. tub-1 is expressed in ciliated amphid head neurons, viz, ASI, ADL, ASK, AWB, ASA, ASJ, in ciliated phasmid tail neurons, viz, PHA and PHB, and in the dopamine-containing neurons, PDE. rbg-3 is also expressed in amphid head neurons and phasmid tail neurons. rbg-3 RNAi greatly decreased fat storage of tub-1 mutants and slightly reduced fat storage in wild-type C. elegans. It was concluded that RBG-3 acted downstream of TUB-1 (Mukhopadhyay et al. 2005). McKay et al. (2003) characterized C. elegans homologues of two mammalian genes, SREBP and C/EBP which control mammalian adipogenesis (Rosen et al. 2000) and found that RNAi targeting of either gene resulted in the loss of fat stores in C. elegans. These authors also found two mitochondrial respiratory chain genes which when inhibited resulted in a depletion of fat stores. Deletion of the nuclear hormone receptor gene, nhr-49, orthologous to mammalian PPARs (peroxisome proliferator-activated receptors) raises the level of fat in C. elegans (van Gilst et al. 2005). This is a key gene since it influences the expression of 13 genes involved in energy metabolism in C. elegans. In a recent paper, Brooks et al. (2009) found that fat storage levels in C. elegans depended on the strain of bacteria they were fed. In the absence of AIY interneurons, C. elegans still showed differential fat storage related to the bacteria they were grown on, indicating that sensory pathways were not essential for this effect. All the mutants tested, apart from pept-1 mutants, which lack a functional peptide transporter, showed differences in fat stores depending on the strain of bacteria they were fed. These authors concluded that nutritional cues deriving from the intestine regulated fat stores independently from any hormonal influences. Interestingly, they found that Nile Red staining of fixed animals provided a better indicator of fat stores than Nile Red staining of live animals. However, in vivo studies investigating lipid storage mechanisms in C. elegans have been undertaken using CARS (coherent anti-Stokes Rahman scattering) microscopy (Hellerer et al. 2007). Accumulation of epidermal fat can be detected using this method.
As mentioned earlier, there is a role for insulin-like receptors and insulin-like peptides in metabolism and growth in C. elegans. DAF-28/insulin-like peptide (56 amino acids with a C terminal of TACCPRLL) activates DAF-2/insulin-like receptor to activate growth and its gene is expressed in ciliated neurons, ASI and ASJ (Pierce et al. 2001; Li 2005). DAF-7 is also expressed in ASIs and is activated by an abundant supply of food (Ren et al. 1996). Activation of this gene in larval stages leads to progression to adult, while inhibition of DAF-7 promotes dauer formation (Ren et al. 1996). Down-regulation of either the insulin or TGF-β pathway results in fat accumulation (Ashrafi 2007). The regulation of fat and feeding has been thoroughly investigated by Greer et al. (2008). In their paper, they present evidence that the accumulation of fat and feeding behaviour is independently regulated. The pathways are summarized in Fig. 2. DAF-7 is a ligand which activates DAF-1 and DAF-4 receptors. While daf-7 is only expressed in ASI interneurons, its receptors are widely expressed in C. elegans. When DAF-1 and DAF-4 are activated, DAF-3 (a transcription factor related to vertebrate Smad-4 proteins) is inactivated. DAF-3 stimulates the release of octopamine and tyramine from RIC and RIM neurons, respectively, and both amines inhibit pharyngeal pumping. Greer et al. investigated the mechanism whereby inactivation of DAF-7 causes fat accumulation in spite of reduced feeding rate. Changes in neuropeptide, acetylcholine, or dopamine levels did not alter fat accumulation in daf-7 mutants, while changes in glutamate function reduced fat accumulation in these mutants. Loss of either mgl-1 or mgl-3 reduced excess fat accumulation in daf-7 mutants, demonstrating a role for metabotropic glutamate receptors in fat accumulation. These mutants have increased de novo fat synthesis which is dependent on daf-3 which may activate an unknown signal from RIM/RIC to regulate fat storage through release of glutamate which in turn activates MGL-1 and MGL-3 receptors in head neurons (Fig. 2 legend for description). In the absence of fasn-1 (which encodes fatty acid synthase), daf-7 mutants have reduced fat levels. fasn-1 is expressed in epidermal cells and these cells together with intestinal cells are the main sites for fat storage in C. elegans.

The role of DAF-7 in the regulation of feeding and fat storage. Expression of daf-7 is only detected in ASI amphid sensory neurons. In the presence of food, a DAF-7 is released and activates DAF-1 and DAF-4 receptors on RIM and RIC interneurons. Activation of these receptors inhibits DAF-3 and in turn no tyramine is released from RIM neurons nor octopamine from RIC neurons. Therefore, the inhibitory receptors, SER-2 and SER-3 receptors, located within the pharyngeal system, are not activated and pharyngeal pumping increases. However, in the absence of food b DAF-7 is not released from ASI neurons and DAF-1 and DAF-4 receptors are not activated. This allows activation of DAF-3 which results in the release of tyramine and octopamine from RIM and RIC neurons, respectively. Tyramine and octopamine activate SER-2 and SER-3 receptors, respectively, which reduce pharyngeal pumping rate and hence feeding. Not shown, but also of note, is that activation of DAF-3 also results in the activation of MGL-1 and MGL-3 metabotropic receptors located on head neurons leading to the stimulation of de novo fat synthesis in epidermal and intestinal cells. The signal from RIM and RIC neurons to activate glutamate release is not known (after Greer et al. 2008)
Carbohydrate stores
Glycogen is the main carbohydrate store in nematodes and is stored in a wide variety of tissues (Behm 1997, 2002). Glucose-1-phosphate is converted to glycogen via UDP-glucose pyrophosphorylase, glycogen synthase and branching enzyme. The genes for UDP-glucose pyrophosphorylase, KO8E3.5, and for branching enzyme, TO4A8.7, have been identified in C. elegans. The disaccharide, trehalose, is the only/major sugar found in the haemolymph of nematodes, while there is little free glucose (Behm 1997). Two genes have been identified in C. elegans, viz, ZK54.2 (trehalose-6-phosphate synthase) and F19H8.1 (trehalose-6-phosphate phosphatase which are involved in the synthesis of trehalose from UDP-glucose (Pellerone et al. 2003). The glyoxylate cycle, that is, the net synthesis of carbohydrate from acetyl CoA 2 carbon substrate occurs in C. elegans embryos where there is a fall in triglyceride levels as the level of carbohydrate rises.
Comparison with position in mammals
There are striking similarities between the physiological pathways that regulate fat storage in C. elegans and in mammals (Mullaney and Ashrafi 2009). For example, insulin, 5-HT, and TUBBY are all involved in fat storage in mammals (Noben-Trauth et al. 1996; Leibowitz and Alexander 1998; Lewis et al. 2002). Feeding and energy expenditure are regulated through the interaction of hormones and neurotransmitters, for example, in mammals the hypothalamus contains centres involved in the control of appetite and satiety (Arora and Anubhut 2006; Konturek et al. 2005). The major transmitters involved are amines, such as 5-HT (anorectic), dopamine and noradrenaline (equivalent to octopamine in C. elegans) (orexigenic), while the hormones include the orexins (orexigenic) and leptin (anorectic). These hormones may be synthesized in the periphery, for example, in the alimentary canal or adipose tissue (leptin) or centrally in the specific parts of the hypothalamus or both in the periphery and centrally, such as ghrelin (Nakazato et al. 2001; Kageyama et al. 2010). From Table 3, it is clear that the mammalian system has a number of hormones which are either anorectic or orexigenic and which interact to regulate food intake. Few homologies have been reported between the mammalian hormones and receptors and those in C. elegans apart from insulin-like peptides (Pierce et al. 2001), NPR-1 and NPY receptor (de Bono and Bargmann 1998), though a CCK-like peptide (NLP-12) which may reduce fat storage (Janssen et al. 2008) and a galanin-like receptor (NPR-9) (Bendena et al. 2008) have recently been identified. Two RFamide peptides have been identified in mammals which influence food intake, NPFF may regulate insulin release and therefore food intake (Panula et al. 1996) and 26RFamide which stimulates food consumption (Chartrel et al. 2003).The major hormones involved in the regulation of feeding in mammals are listed in Table 3 but will not be considered in detail in this review. The hypothalamus is the main area of the brain involved in the regulation of food intake, particularly the arcuate nucleus and the lateral hypothalamus. Leptin plays a major role in the inhibition of feeding and does this in part by inhibiting NPY and activating galanin-like peptide (GALP) and propiomelanocortin (POMC) which is cleaved to release α-melanocyte-stimulating hormone (α-MSH). It is likely that a similar, though simpler, system of peripheral and central sites involving the release of orexigenic and anorectic compounds exists in C. elegans, though no hormone has yet been identified in the fat storing cells in the intestine of C. elegans (Mullaney and Ashrafi 2009). In their review, Mullaney and Ashrafi also list the differences between the mechanisms for storage in C. elegans and in mammals. In the future, it will be interesting to try and establish the equivalent of feeding and satiety centres in C. elegans. In C. elegans, these centres are likely to be at the level of individual neurons or small groups of neurons and the combination of molecular genetic and behavioural approaches employed in the studies reviewed here provide a powerful route to move this field forward.
References
Albertson DG, Thompson JN (1976) The pharynx of Caenorhabditis elegans. Phil Trans Roy Soc Lond B 275:299–325
Alkema MJ, Hunrer-Ensor M, Ringstad N, Horvitz HR (2005) Tyramine functions independently of Octopamine in the Caenorhabditis elegans nervous system. Neuron 46:247–260
Altun-Gultekin Z, Andachi Y, Tsalik EL, Pilgrim D, Kohara Y, Hobert O (2001) A regulatory cascade of three homeobox genes, ceh-10, ttx-3 and ceh-23 controls cell fate specification of a defined interneuron class in C. elegans. Development 128:1951–1969
Apfeld J, Kenyon C (1999) Regulation of lifespan by sensory perception in Caenorhabditis elegans. Nature 402:804–809
Arora S, Anubhut C (2006) Role of neuropeptides in appetite regulation and obesity-A review. Neuropeptides 40:375–401
Ashrafi K (2007) Obesity and the regulation of fat metabolism. WormBook, ed. The C. elegans Research Community, WormBook, doi: 10.1895/wormbook. 1.130.1, http://wormbook.org
Ashrafi K, Chang FY, Watts JL, Fraser AG, Kamath RS, Ahringer J, Ruvkun G (2003) Genome-wide RNAi analysis of Caenorhabditis elegans fat regulating genes. Nature 421:268–272
Avery L (1993a) The genetics of feeding in Caenorhabditis elegans. Genetics 133:897–917
Avery L (1993b) Motor-neuron M3 controls pharyngeal muscle-relaxation timing in Caenorhabditis elegans. J Exp Biol 175:283–297
Avery L (2010) Caenorhabditis elegans behavioural genetics: where are the knobs? BMC Biology 8:69
Avery L, Horvitz HR (1989) Pharyngeal pumping continues after laser killing of the pharyngeal nervous system of C. elegans. Neuron 3:473–485
Avery L, Horvitz HR (1990) Effects of starvation and neuroactive drugs on feeding in C. elegans. J Exp Zool 253:263–270
Avery L, Bargmann CI, Horvitz HR (1993) The Caenorhabditis elegans unc-31 gene affects multiple nervous system-controlled functions. Genetics 134:455–464
Behm CA (1997) The role of trehalose in the physiology of nematodes. Int J Parasitol 27:215–229
Behm CA (2002) Metabolism. In: Lee DL (ed) The biology of nematodes. Taylor & Francis, London, pp 261–290
Bendena WG, Boudreau JR, Papanicolaou T, Maltby M, Tobe SS, Chin-Sang ID (2008) A Caenorhabditis elegans allatostatin/galanin-like receptor NPR-9 inhibits local search behaviour in response to feeding cues. Proc Natl Acad Sci USA 195:1339–1342
Brock TJ, Browse J, Watts JL (2006) Genetic regulation of unsaturated fatty acid composition in C. elegans. PLoS Genet 2:e108
Brockie PJ, Madsen DM, Zheng Y, Mellem J, Maricq AV (2001) Differential expression of glutamate subunits in the nervous system of Caenorhabditis elegans and their regulation by the homeodomain protein UNC-42. J Neurosci 21:1510–1522
Brooks KK, Liang B, Watts JL (2009) The influence of bacterial diet on fat storage in C. elegans. PLoS ONE 4:e7545
Carre-Pierrat M, Baillie D, Johnsen R, Hyde R, Hart A, Granger L, Segalat L (2006) Characterization of the Caenorhabditis elegans G protein-coupled serotonin receptors. Invert Neurosci 6:189–205
Chalansani SH, Kato S, Albrecht DR, Nakagawa T, Abbott LF, Bargmann CI (2010) Neuropeptide feedback modifies odor-evoked dynamics in Caenorhabditis elegans olfactory neurons. Nature Neurosci 13:615–621
Chalasani SH, Chronis N, Tsunozaki M, Gray JM, Ramot D, Goodman MB, Bargmann CI (2007) Dissecting a circuit for olfactory behaviour in Caenorhabditis elegans. Nature 450:63–70
Chao MY, Komatsu H, Fukuto HS, Dionne HM, Hart AC (2004) Feeding status and serotonin rapidly and reversibly modulate a Caenorhabditis elegans chemosensory circuit. Proc Natl Acad Sci USA 101:15512–15517
Chartrel N, Dujardin C, Anouar Y, Leprince J, Decker A, Clerens S, Do-Rego JC et al (2003) Identification of 26RFa, a hypothalamic neuropeptide of the RFamide peptide family with orexigenic activity. Proc Natl Acad Sci USA 100:15247–15252
Chiang JTA, Steciuk M, Shtonda B, Avery L (2006) Evolution of pharyngeal behaviour and neuronal function in free-living soil nematodes. J Exp Biol 209:1859–1873
Coates JC, de Bono M (2002) Antagonistic pathways in neurons exposed to body fluid regulate social feeding in Caenorhabditis elegans. Nature 419:925–929
Cohen M, Reale V, Olofsson B, Knights A, Evans P, de Bono M (2009) Coordinated regulation of foraging and metabolism in C.elegans by RFamide neuropeptide signaling. Cell Metab 9:375–385
Croll NA (1975) Indolealkylamines in the coordination of nematode behavioural activities. Can J Zool 53:894–903
Croll NA, Smith JM (1978) Integrated behaviour in the feeding phase of Caenorhabditis elegans (Nematoda). J Zool (Lond.) 184:507–517
David MW, Fleischhauser R, Dent JA, Joho RH, Avery L (1999) A mutation in the C. elegans EXP-2 Potassium channel that alters feeding behavior. Science 286:2501–2504
de Bono M, Bargmann CI (1998) Natural variation in a Neuropeptide Y Receptor homolog modifies social behaviour and food response in C. elegans. Cell 94:679–689
Dent JA, Davies MW, Avery L (1997) avr-15 encodes a chloride channel subunit that mediates inhibitory glutamatergic neurotransmission and ivermectin sensitivity in Caenorhabditis elegans. EMBO J 16:5867–5879
Dufresne M, Seva C, Fourmy D (2006) Cholecystokinin and gastrin receptors. Physiol Rev 86:805–847
Franks CJ, Holden-Dye L, Bull K, Luedtke S, Walker RJ (2006) Anatomy, physiology and pharmacology of Caenorhabditis elegans pharynx: a model to define gene function in a simple neural system. Invert Neurosci 6:105–122
Gao XB, van den Pol AN (2001) Melanin concentrating hormone depresses synaptic activity of Glutamate and GABA neurons from rat lateral hypothalamus. J Physiol 533:237–252
Gray JM, Hill JJ, Bargmann CI (2005) A circuit for navigation in Caenorhabditis elegans. Proc Natl Acad Sci USA 102:3184–3191
Greer ER, Perez C, Van Gilst MR, Lee BH, Ashrafi K (2008) Neural and molecular dissection of a C.elegans sensory circuit that regulates fat and feeding. Cell Met 8:118–131
Hamdan FF, Ungrin MD, Abramovitz M, Ribeiro P (1999) Characterization of a novel serotonin receptor from Caenorhabditis elegans: cloning and expression of two splice variants. J Neurochem 72:1372–1383
Hart AC, Sims S, Kaplan JM (1995) Synaptic code for sensory modalities revealed by C. elegans GLR-1 glutamate receptor. Nature 378:82–85
Hellerer T, Axang C, Brackmann C, Hillertz P, Pilon M, Enejder A (2007) Monitoring of lipid storage in Caenorhabditis elegans using coherent ant-Stokes Raman scattering (CARS) microscopy. Proc Natl Acad Sci USA 104:14658–14663
Hills T, Brockie PJ, Maricq AV (2004) Dopamine and glutamate control area-restricted search behaviour in Caenorhabditis elegans. J Neurosci 24:1217–1225
Hobson RJ, Geng J, Gray AD, Komuniecki RW (2003) SER-7b, a constitutively active Gα-coupled 5-HT7-like receptor expressed in the Caenorhabditis elegans M4 pharyngeal motoneuron. J Neurochem 87:22–29
Hobson RJ, Hapiak VM, Xiao H, Buehrer KL, Kommuniecki PR (2006) SER-7 a Caenorhabditis elegans 5HT7-like receptor is essential for the 5-HT stimulation of pharyngeal pumping and egg-laying. Genetics 172:159–169
Horvitz HR, Chalfie M, Trent C, Sulston JE, Evans PD (1982) Serotonin and octopamine in the nematode C. elegans. Science 216:1012–1014
Husson SJ, Clynen E, Baggerman G, Janssen T, Schoofs L (2006) Defective processing of neuropeptide precursors in Caenorhabditis elegans lacking proprotein convertase 2 (KPC-2/EGL-3): mutant analysis by mass spectrometry. J Neurochem 98:1999–2012
Husson SJ, Janssen T, Baggerman G, Bogert B, Kahn-Kirby AH, Ashrafi K, Schoofs L (2007) Impaired processing of FLP and NLP peptides in carboxypeptidase E (EGL-21)-deficient Caenorhabditis elegans as analyzed by mass spectrometry. J Neurochem 102:246–260
Janssen T, Meelkop E, Lindemans M, Verstraelen K, Husson SJ, Temmerman L, Nachman RJ, Schoofs L (2008) Discovery of a Cholecystokinin-Gastrin-like signaling system in Nematodes. Endocrinology 149:2826–2839
Kageyama H, Takenoya F, Shiba K, Shioda S (2010) Neuronal circuits involving ghrelin in the hypothalamus-mediated regulation of feeding. Neuropeptides 44:133–138
Kang C, Avery L (2009) Systemic regulation of starvation response in Caenorhabditis elegans. Genes Develop 23:12–17
Kim K, Li C (2004) Expression and regulation of an FMRFamide-related Neuropeptide gene family in Caenorhabditis elegans. J Comp Neurol 475:540–550
Kimura KD, Tissenbaum HA, Liu Y, Ruvkun G (1997) daf-2, an insulin receptor-like gene that regulates longevity and diapause in Caenorhabditis elegans. Science 277:942–946
Konturek PC, Konturek JW, Czesnikiewcz-Guzik M, Brzozowski T, Sito E, Konturek SJ (2005) Neuro-hormonal control of food intake; Basic mechanisms and clinical implications. J Physiol Pharmacol 56(Supp 6):5–25
Lee BH, Ashrafi K (2008) A TRPV channel modulates C. elegans neurosecretion, larval starvation survival, and adult lifespan. PLoS Genetics 4:e1000213
Lee RN, Lobel L, Hengartner M, Horvitz HR, Avery L (1999) Eat-4, a homolog of a mammalian sodium-dependent inorganic phosphate co-transporter, is necessary for glutamatergic neurotransmission in Caenorhabditis elegans. J Neurosci 19:159–167
Leibowitz S, Alexander J (1998) Hypothalamic serotonin in control of eating behaviour, meal size and body weight. Biol Psychiatry 44:851–864
Lewis GF, Carpentier A, Adell K, Giacca A (2002) Disordered fat storage and mobilization in the pathogenesis of insulin resistance and Type 2 diabetes. Endocrin Rev 23:201–229
Li C (2005) The ever-expanding neuropeptide gene families in the nematode Caenorhabditis elegans. Parasitology 131 (Suppl): S109–S127
Li C, Kim K (2008) Neuropeptides.WormBook, ed. The C. elegans Research Community, WormBook, doi: 10.1895/wormbook.1.142.1, http://www.wormbook.org
Li H, Avery L, Denk W, Hess GP (1997) Identification of chemical synapses in the pharynx of Caenorhabditis elegans. Proc Natl Acad Sci USA 94:5912–5916
Li W, Kennedy SG, Ruvkun G (2003) daf-28 encodes a C. elegans insulin superfamily member that is regulated by environmental cues and acts in the DAF-2 signaling pathway. Genes Develop 17:844–858
Li W, Feng Z, Sternberg PW, Shawn Xu XZ (2006) A C. elegans stretch receptor neuron revealed by a mechanosensitive TRP channel homologue. Nature 440:684–687
Liu LX, Spoerke JM, Mulligan EL, Chen J, Reardon B, Westlund B, Abel K, Armstrong B, Hardiman G, King J et al (1999) High-throughput isolation of Caenorhabditis elegans deletion mutants. Genome Res 9:859–867
Mak HY, Nelson LS, Basson M, Johnson CD, Ruvkun G (2006) Polygenic control of Caenorhabditis elegans fat storage. Nat Genet 38:363–368
Maricq AV, Peckol E, Driscoll M, Bargmann C (1995) glr-1, a C.elegans glutamate receptor that mediates mechanosensory signaling. Nature 378:78–81
McKay RM, McKay JP, Avery L, Graff JM (2003) c. elegans: A model for exploring the genetics of fat storage. Devel Cell 4:131–142
McKay JP, Raizen DM, Gottschalk A, Schafer WR, Avery L (2004) eat-2 and eat-18 are required for nicotinic transmission in the Caenorhabditis elegans pharynx. Genetics 166:161–169
McKay RM, McKay JP, Suh JM, Avery L, Graff JM (2007) Tripeptidyl peptidase II promotes fat formation in a conserved fashion. EMBO Reports 8:1183–1189
Mellem JE, Brockie PJ, Zheng Y, Madsen DM, Maricq AV (2002) Decoding of polymodel sensory stimuli by postsynaptic glutamate receptors in C. elegans. Neuron 36:933–944
Mukhopadhyay A, Deplancke B, Walhout AJM, Tissenbaum HA (2005) C. elegans tubby regulates life span and fat metabolism by two independent mechanisms. Cell Metabol 2:35–42
Mullaney BC, Ashrafi K (2009) C. elegans fat storage and metabolic regulation. Biochim Biophys Acta 1791:474–478
Nakazato M, Murakami N, Date Y, Kojima M, Matsuo H, Kangawa K, Matsukura S (2001) A role for ghrelin in the central regulation of feeding. Nature 409:194–198
Nass R, Merchant KM, Ryan T (2008) Caenorhabditis elegans in Parkinson’s disease drug discovery: addressing an unmet clinical need. Mol Interv 8:284–293
Nathoo AN, Moeller RA, Westlund BA, Hart AC (2001) Identification of neuro-peptide-like gene families in Caenorhabditis elegans and other species. Proc Natl Acad Sci USA 98:14000–14005
Niacaris T, Avery L (2003) Serotonin regulates repolarization of the C. elegans pharyngeal muscle. J Exp Biol 206:223–231
Noben-Trauth K, Naggert JK, North MA, Nishina PM (1996) A candidate gene for the mouse mutation tubby. Nature 380:534–538
O’Rahilly S, Farooqi IS (2008) Human obesity: a heritable neurobehavioural disorder that is highly sensitive to environmental conditions. Diabetes 57:2905–2910
Packham R, Walker RJ, Holden-Dye L (2010) The effect of a selective octopamine antagonist, epinastine, on pharyngeal pumping in Caenorhabditis elegans. Invert Neurosci 10:47–52
Panula P, Aarnisalo AA, Wasowicz K (1996) Neuropeptide FF, a mammalian neuropeptide with multiple functions. Prog Neurobiol 48:461–487
Papaioannou S, Marsden D, Franks CJ, Walker RJ, Holden-Dye L (2005) Role of a FMRFamide-like family of neuropeptides in the pharyngeal nervous system of Caenorhabditis elegans. J Neurobiol 65:304–319
Papaiouannou S, Holden-Dye L, Walker RJ (2008) The actions of Caenorhabditis elegans neuropeptde-like peptides (NLPs) on body wall muscle of Ascaris suum and pharyngeal muscle of C. elegans. Acta Biol Hung 59(Suppl):189–197
Pellerone FI, Archer CA, Behm CA, Grant WN, Lacey MJ, Somerville AC (2003) Trehalose metabolism genes in Caenorhabditis elegans and filarial nematodes. Int J Parasitol 33:1195–1206
Pierce SB, Costa M, Wisotzkey R, Devadhar S, Homburger SA, Ferguson KC, Heller J, Platt DM, Pasquinelli AA, Liu LX, Doberstein SK, Ruvkun G (2001) Regulation of DAF-2 receptor signaling by human insulin and ins-1, a member of the unusually large and diverse C. elegans insulin gene family. Genes Dev 15:672–686
Rand JB, Duerr JS, Frisby DL (2000) Neurogenetics of vesicular transporters in C. elegans. FASEB J 14:2414–2422
Ranganathan R, Cannon SC, Horvitz HR (2000) MOD-1 is a serotonin-gated chloride channel that modulates locomotory behaviour in C. elegans. Nature 408:470–475
Ren P, Lim CS, Johnsen R, Albert PS, Pilgrim D, Riddle DL (1996) Control of C. elegans larval development by neuronal expression of a TGF-β homolog. Science 274:1389–1391
Rogers CM, Franks CJ, Walker RJ, Burke JF, Holden-Dye L (2001) Regulation of the pharynx of Caenorhabditis elegans by 5-HT, octopamine and FMRFamide-like neuropeptides. J Neurobiol 49:235–244
Rogers C, Reale V, Kim K, Chatwin H, Li C, Evans P, de Bono M (2003) Inhibition of Caenorhabditis elegans social feeding by FMRFamide-related peptide activation of NPR-1. Nat Neurosci 6:1178–1185
Rosen E, Walkey C, Pulgserver P, Splegelman B (2000) Transcriptional regulation of adipogenesis. Genes Dev 14:1293–1307
Sawin ER, Ranganathan R, Horvitz HR (2000) C. elegans locomotion rate is modulated by the environment through a dopaminergic pathway and by experience through a serotonergic pathway. Neuron 26:619–631
Seymour MK, Wright KA, Doncaster CC (1983) The action of the anterior feeding apparatus of Caenorhabditis elegans (Nematoda: Rhabditida). J Zool 201:527–539
Shtonda BB, Avery L (2006) Dietary choice behaviour in C. elegans. J Exp Biol 209:89–102
Speese S, Petri M, Schuske K, Ailon M, Ann K, Iwasaki K, Jorgensen EM, Martin TFJ (2007) UNC-31 (CAPS) is required for dense-core vesicle but not synaptic vesicle exocytosis in Caenorhabditis elegans. J Neurosci 27:6150–6162
Srinivasan S (2008) Serotonin regulates C. elegans fat and feeding through independent molecular mechanisms. Cell Metabol 7:533–544
Steger KA, Avery L (2004) The GAR-3 muscarinic receptor cooperates with calcium signals to regulate muscle contraction in the Caenorhabditis elegans pharynx. Genetics 167:633–643
Suo S, Kimura Y, Van Tol HHM (2006) Starvation induces cAMP response element-binding protein-dependent gene expression through Octopamine-Gq signaling in Caenorhabditis elegans. J Neurosci 26:10082–10090
Suo S, Culotti JG, Van Tol HHM (2009a) Dopamine counteracts octopamine signalling in a neural circuit mediating food response in C. elegans. EMBO J 28:2437–2448
Suo S, Culotti JG, Van Tol HHM (2009b) Dopamine suppresses octopamine signalling in C. elegans: possible involvement of dopamine in the regulation of lifespan. Aging 1:870–874
Sze JY, Victor M, Loer C, Shi Y, Ruvkun G (2000) Food and metabolic signaling defects in a Caenorhabditis elegans serotonin-synthesis mutant. Nature 403:560–564
Tsalik EL, Hobert O (2003) Functional mapping of neurons that control locomotory behavior in Caenorhabditis elegans. J Neurobiol 56:178–197
Tsalik EL, Niacaria T, Wenick AS, Pau K, Avery L, Hobert O (2003) LIM homeobox gene-dependent expression of biogenic amine receptors in restricted regions of the C. elegans nervous system. Develop Biol 263:81–102
van Gilst MR, Hadjivassiliou H, Jolly A, Yamamoto KR (2005) Nuclear hormone receptor NHR-49 controls fat consumption and fatty acid composition in C. elegans. PLos Biol 3:e53
Walker DS, Gower NJD, Ly S, Bradley GL, Baylis HA (2002) Regulated disruption of Inositol 1, 4, 5-Triphosphate signalling in Caenorhabditis elegans reveals new functions in feeding and embryogenesis. Mol Biol Cell 13:1329–1337
Watts JL (2008) Fattening up without overeating. Cell Metabolism 8:95–96
Watts JL (2009) Fat synthesis and adiposity regulation in Caenorhabditis elegans. Trends Endocrin Metabol 20:58–65
Wenick AS, Hobert O (2004) Genomic cis-regulatory architecture and trans-acting regulators of a single interneuron-specific gene battery in C. elegans. Develop Cell 6:757–770
White J, Southgate E, Thomson JN, Brenner S (1986) The structure of the nervous system of the nematode Caenorhabditis elegans. Phil Trans Roy Soc Lond B 314:1–340
Wicks SR, Roehrig CJ, Rankin CH (1996) A dynamic network simulation of the nematode tap withdrawal circuit: predictions concerning synaptic function using behavioural criteria. J Neurosci 16:4017–4031
Wolkow CA, Kimura KD, Lee MS, Ruvkun G (2000) Regulation of C. elegans lifespan by insulin-like signaling in the nervous system. Science 290:147–150
Xiao H, Hapiak VM, Smith KA, Lin L, Hobson RJ, Plenefisch J, Komuniecki R (2006) SER-1, a Caenorhabditis elegans 5-HT2-like receptor, and a multi-PDZ domain containing protein (MPZ-1) interact in vulval muscle to facilitate serotonin-stimulated egg-laying. Develop Biol 298:379–391
You Y-J, Kim J, Cobb M, Avery L (2006) Starvation activates MAP kinase through the muscarinic acetylcholine pathway in Caenorhabditis elegans pharynx. Cell Metabol 3:237–245
Acknowledgments
Sarah Luedtke was supported by a studentship from the Gerald Kerkut Charitable Trust.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Luedtke, S., O’Connor, V., Holden-Dye, L. et al. The regulation of feeding and metabolism in response to food deprivation in Caenorhabditis elegans . Invert Neurosci 10, 63–76 (2010). https://doi.org/10.1007/s10158-010-0112-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10158-010-0112-z