
Abstract
Renal tubular potassium (K+) channels play important roles in the formation of cell-negative potential, K+ recycling, K+ secretion, and cell volume regulation. In addition to these physiological roles, it was reported that changes in the activity of renal tubular K+ channels were involved in exacerbation of renal cell injury during ischemia and endotoxemia. Because ischemia and endotoxemia stimulate production of cytokines in immune cells and renal tubular cells, it is possible that cytokines would affect K+ channel activity. Although the regulatory mechanisms of renal tubular K+ channels have extensively been studied, little information is available about the effects of cytokines on these K+ channels. The first report was that tumor necrosis factor acutely stimulated the single channel activity of the 70 pS K+ channel in the rat thick ascending limb through activation of tyrosine phosphatase. Recently, it was also reported that interferon-γ (IFN-γ) and interleukin-1β (IL-1β) modulated the activity of the 40 pS K+ channel in cultured human proximal tubule cells. IFN-γ exhibited a delayed suppression and an acute stimulation of K+ channel activity, whereas IL-1β acutely suppressed the channel activity. Furthermore, these cytokines suppressed gene expression of the renal outer medullary potassium channel. The renal tubular K+ channels are functionally coupled to the coexisting transporters. Therefore, the effects of cytokines on renal tubular transporter activity should also be taken into account, when interpreting their effects on K+ channel activity.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Potassium (K+) channels in renal tubular epithelia greatly contribute to the homeostasis of salts and water [1, 2]. One of their crucial roles is the formation of the cell-negative potential. This potential difference serves as a driving force for the electrogenic passive transport of solutes, such as apical sodium (Na+) entries through the Na+-glucose cotransporter (SGLT) in proximal tubule cells and the epithelial Na+ channel (ENaC) in principal cells of the cortical collecting duct (CCD) [1, 2]. The apical K+ channels in the CCD provide a pathway of K+ secretion [1, 2]. The renal tubular K+ channels are also important K+ recycling pathways for the apical sodium–potassium−chloride (Na+–K+–2Cl−) cotransporter (NKCC) in the thick ascending limb (TAL) and the basolateral sodium–potassium adenosine triphosphatase (Na+−K+-ATPase) along the nephron [1, 2]. Furthermore, they are involved in regulatory volume decrease of tubular cells [1, 2]. To date, many types of renal tubular K+ channels have been identified and characterized in each nephron segment, by using electrophysiological and molecular biological techniques [1, 2]. The activity of these K+ channels is subjected to regulation by a variety of factors, which include intracellular pH, ATP, calcium ion (Ca2+), membrane stretch, voltage, cyclic nucleotides, protein kinases/phosphatases, nitric oxide, arachidonic acid metabolites, etc. [1, 2].
The still expanding cytokine family consists of a variety of multifunctional proteins, which act as the key molecules in immunomodulation and inflammation [3]. They exert various effects on almost all organs. The nervous system, cardiovascular system, respiratory system, gastrointestinal tracts, and kidney are also targets of cytokines [4–6]. Although cytokines are secreted mainly by immune cells in response to microbial infection, other cell types are known to secrete cytokines [3]. In the kidney, proximal tubule cells are thought to act as the proinflammatory cells, being involved in renal dysfunction in patients suffered from chronic renal rejection or ischemia [7]. There are reports demonstrating that proximal tubule cells produced cytokines in response to lipopolysaccharide (LPS) or albumin [8, 9]. In addition, LPS and angiotensin II stimulated cytokine production in the TAL cells [10, 11].
It has been reported that several proinflammatory cytokines, such as interferon-γ (IFN-γ), interleukin-1β (IL-1β) and tumor necrosis factor-α (TNF-α) modulated Na+ reabsorption in renal tubular epithelia [12–17]. Since the driving force of the transepithelial Na+ reabsorption is dependent on the K+ channel activity, as well as Na+–K+-ATPase [1, 2], it is important to know whether cytokines would affect the K+ channels; however, there are few reports, regarding the effects of cytokines on K+ channel activity in renal tubular epithelia. This is in sharp contrast to the accumulating evidence for the effects of cytokines on neuronal ion channels [18]. In this review, we focus on how cytokines affect renal tubular K+ channels, and discuss their physiological and pathological significances.
Effects of cytokines on K+ channels in renal tubular epithelia
The first report addressing the effects of cytokines on renal tubular K+ channels was presented by Wei et al. [19]. They demonstrated that TNF acutely stimulated activity of an apical 70 pS K+ channel in rat TAL, using the cell-attached mode of the patch clamp technique. The stimulatory effect of TNF was highly likely mediated by tyrosine dephosphorylation, since an inhibitor of protein tyrosine phosphatase (PTP) abolished the TNF-induced activation of the channel. The finding that TNF increased PTP activity in the TAL also supported this notion. Although the structure of the 70 pS K+ channel in the TAL is not fully understood, Lu et al. [20] reported that the renal outer medullary potassium channel (ROMK) would be a critical subunit of the 70 pS channel. It is well known that protein tyrosine kinase (PTK) and PTP modulate the expression of ROMK in the cytoplasmic membrane [21]. The PTK-mediated tyrosine phosphorylation causes endocytosis of ROMK, whereas the PTP-mediated dephosphorylation blocks it [21]. Thus, the balance between PTK and PTP is one of the key factors determining the membrane expression of ROMK at a given time. However, the stimulatory effect of the TNF/PTP pathway on the 70 pS K+ channel seems not to be accounted for by the expression of the channel. The effect of tyrosine phosphorylation on the 70 pS K+ channel is different from that on ROMK in that PTK inhibits the 70 pS K+ channel in cell-free inside-out patches [22]. This observation suggests that the PTK-mediated tyrosine phosphorylation might directly suppress the activity of this K+ channel rather than enhance its endocytosis. The effect of PTP on the TAL’s 70 pS K+ channel in inside-out patches is still unknown. Nonetheless, it is possible that the TNF/PTP-mediated tyrosine dephosphorylation of the 70 pS K+ channel was engaged in modulation of the channel activity itself.
A ROMK-like inwardly rectifying K+ channel with an inward conductance of 30 pS was also present in the apical membrane of the TAL [19, 23]. In contrast to the 70 pS K+ channel, the activity of the ROMK-like K+ channel was not affected by TNF [19]. The ROMK channel consists of several isoforms (ROMK1, 2 and 3), all of which conserve the consensus phosphorylation site for PTK at the C-terminus [2]. However, only ROMK1 is subjected to the modulation by PTK and PTP [24]. The different sensitivity to PTK and PTP seems to arise from structural differences in the N-termini of ROMK isoforms. The N-termini of ROMK2 and 3 isoforms are different from that of ROMK1 and lacking in one PKC phosphorylation site [2, 25]. It was reported that the PKC site at the N-terminus would play a role in mediating the endocytosis of ROMK induced by tyrosine phosphorylation [25]. ROMK1 localizes in the CCD and outer medullary collecting duct, whereas the isoforms expressed in the TAL are ROMK2 and 3 [2]. Such distribution pattern of ROMK isoforms is consistent with the different responsiveness of the native ROMK-like K+ channels to PTK and PTP in each nephron segment. Membrane expression of the ROMK-like K+ channel in the principal cells of CCD, but not in the TAL, is reduced by tyrosine phosphorylation and increased by tyrosine dephosphorylation [21, 22, 25]. The absence of ROMK1 in the TAL well explains the ineffectiveness of TNF on the TAL’s ROMK-like K+ channel.
Although the activity of the TAL’s ROMK-like K+ channel was not acutely affected by TNF, the ROMK gene expression in the whole kidney was reduced by cytokines [17]. Schmidt et al. [17] reported that intraperitoneal injection of IL-1β, IFN-γ, or TNF-α into the mouse depressed the mRNA level of ROMK in the kidney. Because the systemic administration of these cytokines caused hypotension [17], it was possible that the suppressive effects of cytokines on the ROMK gene expression would have been indirectly mediated by the reduced blood flow in the kidney. However, they also confirmed that IL-1β, IFN-γ, or TNF-α reduced the ROMK gene expression in a mouse CCD cell line, M1, to an extent similar to those observed in the in vivo experiments [17]. Thus, the direct actions of these cytokines on the ROMK gene expression were strongly suggested.
In addition to the K+ channels in distal nephron segments, those in the proximal tubule cells have recently been reported to be affected by cytokines [26, 27]. In cultured human renal proximal tubule epithelial cells (RPTECs) isolated from the normal kidney, an inwardly rectifying K+ channel with an inward conductance of 40 pS is most frequently observed under the control condition [28]. Although no information is currently available, with regard to its molecular property and localization along the nephron, its open probability was generally high, with the mean value being about 0.8, and independent of membrane potential [28]. It was also reported that the activity of the 40 pS K+ channel was modulated by intracellular ATP [28], pH [28] and protein phosphorylation processes [28–30].
IFN-γ exerts a delayed suppressive effect and an acute stimulatory effect on the activity of this 40 pS K+ channel [26, 27]. The IFN-γ-induced suppression and stimulation of the channel are highly likely dependent on the IFN-γ receptor and its closely related janus kinase (JAK), since the JAK inhibitors blocked both effects [27]. The delayed suppression of channel activity was mediated, at least in part, by nitric oxide (NO) [26, 27]. It was reported that NO itself possessed a dose-dependent biphasic effect on the activity of the 40 pS K+ channel; a protein kinase G (PKG)-dependent stimulatory effect at micromolar concentrations and a peroxynitrite-dependent suppressive effect at higher millimolar concentrations [29]. Incubation of RPTECs with IFN-γ for 24 h augments the expression of inducible NO synthase (iNOS), which results in production of a large amount of NO [26, 27]. The excessive NO then reacts with superoxide to form peroxynitrite, which in turn impairs K+ channel activity through oxidation or nitrosylation of the channel and/or its related proteins [27]. On the other hand, the mechanisms involved in the acute stimulatory effect of IFN-γ are still unknown. The activity of the 40 pS K+ channel is stimulated by protein kinase A (PKA) [28] and phosphatidylinositol-3-kinase [27], as well as PKG [30]. However, IFN-γ seems to circumvent these pathways to stimulate the channel activity [27].
IL-1β is another cytokine that modulates the activity of the 40 pS K+ channel in RPTECs [26]. Although it is reported that IL-1β enhanced iNOS expression in some tissues, this cytokine does not seem to affect it in human proximal tubule cells, including RPTECs [26, 31]. Therefore, IL-1β is devoid of the NO-dependent delayed suppressive effect observed with IFN-γ [26, 27]; however, IL-1β acutely suppressed channel activity [26]. This suppressive effect is abolished by an inhibitor of protein kinase C (PKC) and is not reversed by activation of PKA and PKG (our unpublished observations). Kubokawa et al. [32] reported that at least two distinct phosphorylation sites would exist in the 40 pS K+ channel and/or its related proteins. One is involved in the channel activation by PKA and PKG, whereas the other contributes to the channel suppression by PKC and Ca2+/calmodulin-dependent protein kinase II [32]. Even though the phosphorylation of the former site opens the channel, subsequent phosphorylation of the latter one by PKC closes it. They also demonstrated that a PKC inhibitor had little effect on the basal channel activity, suggesting that the PKC activity would be relatively low under the control condition in RPTECs [32]. Hippocampal Ca2+ channel [33], a middle ear Na+ channel [34], and the intestinal cystic fibrosis transmembrane conductance regulator [35] are also affected by IL-1β through activation of PKC.
Implication of the cytokine-mediated K+ channel modulation in the transport of solutes in renal tubular epithelia
It is generally thought that K+ channels and transporters in renal tubular epithelia are functionally coupled [1, 2]. In proximal tubule cells, basolateral K+ channels cooperate with the basolateral Na+–K+-ATPase to provide a driving force for Na+ entry through the apical transporters, such as SGLT [1, 2]. The cooperation of basolateral K+ channels with the Na+–K+-ATPase is also observed in distal nephron segments [1, 2]. In the TAL, apical K+ channels serve as the K+ recycling pathways for the apical NKCC, facilitating transport of Na+ and Cl− [1, 2]. In principal cells of the CCD, apical Na+ entry through the ENaC is partly dependent on the cell-negative potential generated by apical and basolateral K+ channels [1, 2]. Therefore, the transport activity of solutes should change in parallel with the altered K+ channel activity.
Compared to the renal tubular K+ channels, effects of cytokines on renal transporters are relatively well documented [10, 12–17, 36–38]. Most of these investigations revealed the suppressive effects of cytokines on renal tubular transport [10, 12–14, 16, 17, 36–38]. IL-1β reduced 86Rb+ uptake [10], Na+ reabsorption [12], and the activity and protein expression of Na+–K+-ATPase [13, 14]. TNF-α similarly reduced 86Rb+ uptake [10], the activity and protein expression of Na+–K+-ATPase [16]. It also reduced protein expression of NKCC [16]. IFN-γ, as well as IL-1β and TNF-α, suppressed gene expression of various transporters, including Na+–K+-ATPase [17, 36, 37], sodium−proton (Na+–H+) exchanger [17], Na+–Cl− cotransporter [36], NKCC [17], SGLT [37], glucose transporters [37], and urea transporters [38]. Such suppressive effects of cytokines on renal transport of solutes are consistent with their suppressive effects on ROMK gene expression [17] in the kidney and the K+ channel activity in RPTECs [26, 27]. However, there are two exceptions.
Escalante et al. [10] reported that TNF and IL-1 reduced ouabain-sensitive 86Rb+ uptake in the TAL. This result suggested that both cytokines inhibited the basolateral Na+–K+-ATPase, which might well result in decreased apical Na+ entry. According to the report by Wei et al. [19], however, TNF stimulated the activity of the apical 70 pS K+ channel in the TAL. Because the apical 70 pS K+ channel, as well as the 30 pS channel, serves as a recycling pathway for the apical NKCC [1, 2], the TNF-induced increase in channel activity should be related to increased apical Na+ entry. It was suggested that such a discrepancy would partly be due to a difference in the temporal factor. TNF reduced 86Rb+ uptake after a 24-h incubation period [10], whereas stimulation of channel activity by TNF occurred in a few minutes [19]. In addition to the acute channel stimulation, TNF had been shown to induce expression of cyclooxygenase (COX) and the subsequent generation of prostaglandin E2 (PGE2) which suppressed the activity of the 70 pS K+ channel [39]. Wei et al. [19] speculated that the effects of TNF on channel activity would be time-dependently biphasic and that the chronic COX/PGE2-dependent channel suppression would be consistent with the TNF-induced inhibition of 86Rb+ uptake. Nonetheless, the significance of the acute stimulatory effect of TNF on channel activity is obscure. The acute stimulation of K+ channel activity by IFN-γ shown in RPTECs is also a matter of controversy.
As described above, many lines of evidence indicate that cytokines suppress the renal tubular transport of solutes [10, 12–14, 16, 17, 36–38]; however, these suppressive effects were generally observed after prolonged treatment with cytokines, ranging from 1 to 24 h. This indicates that the early phase effects of cytokines on tubular transport are still poorly understood, although a few reports exist. Vinciguerra et al. [15] reported that TNF-α rapidly (~15 min) increased the amiloride-sensitive short circuit current, which was accompanied by cell-surface recruitment of Na+–K+-ATPase, in cultured mpkCCDcL4 cells. It was also reported that LPS-injected rats exhibited immediate antinatriuresis in the first few hours after injection, which was later followed by natriuresis [40]. Thus, it is possible that some cytokines might also possess time-dependent biphasic effects on renal tubular transporters, like the case with K+ channels, depending on the experimental systems employed. It would be the best way to simultaneously examine the effects of cytokines on the K+ channels and transporters in the renal tubular epithelia.
Relationship between effects of cytokines on K+ channel activity and renal cell injury
It is well known that proinflammatory cytokines play pivotal roles in promoting cell injury in many organs during inflammatory diseases [3]. Endotoxemia-induced acute renal failure is a typical clinical incident that highlights the harmful effects of proinflammatory cytokines on the kidney [41]. Furthermore, therapeutic use of cytokines, such as IFNs, in renal cell carcinoma or viral hepatitis sometimes results in undesirable severe renal dysfunction [42]. In various experimental models, proinflammatory cytokines are reported to cause glomerulonephritis accompanied by proteinuria, tubulointerstitial fibrosis, and apoptosis/necrosis of tubular cells [43, 44]. These effects are generally thought to be mediated by activation of caspases and various transcription factors, such as NFκB, AP-1, and STAT [3]. The activated transcription factors induce synthesis of many effector proteins, including other cytokines, chemokines, matrix metalloproteases, iNOS and adhesion molecules [3], all of which could participate in renal cell injury. It is possible that the renal cell injury by cytokines would be mediated by modulation of K+ channel activity. In fact, it was reported that TNF induced apoptotic cell death in a rat liver cell line through activation of K+ channels [45].
In support of this notion, there are also reports suggesting that changes in activity of renal tubular K+ channels would be involved in renal cell injury during ischemia or endotoxemia [46–51]. Ischemia, as well as endotoxemia, is known to stimulate production of cytokines [17]. These cytokines may enhance renal cell injury by modulating K+ channel activity. However, there is still controversy about the causal relationship between the direction of the change in K+ channel activity and the severity of renal cell injury. Some investigators reported that inhibiting K+ channel activity by glibenclamide reduced hypoxia- or ischemia/reperfusion-induced renal tubular cell injury in isolated rat proximal tubules [46], isolated perfused rat kidneys [47], and rats in vivo [48]. These observations suggest that an increase in K+ channel activity leads to exacerbation of renal cell injury. In contrast, other investigators reported that glibenclamide enhanced renal cell injury in isolated perfused rat kidney [49], a human proximal tubule cell line (HK2) [50], isolated mouse proximal tubules [50], and a porcine proximal tubule cell line (LLC-PK1) [51], suggesting that a decrease in K+ channel activity leads to renal cell injury. Differences in experimental systems and cell conditions may partly account for the discrepant results. In any reports described above, the precise mechanisms linking the K+ channel activity to renal cell injury are not fully clarified. One explanation is that intracellular K+ loss through the activated K+ channel facilitates cell shrinkage, which triggers apoptosis [52]. In addition, changes in K+ channel activity alter the driving force for Ca2+ entry through Ca2+-permeable channels [53]. Thus, intracellular Ca2+-dependent factors may be involved in the K+ channel-mediated renal cell injury [53]. It should also be clarified whether the cytokine-mediated modulation of K+ channel activity would actually be involved in renal cell injury.
Conclusion
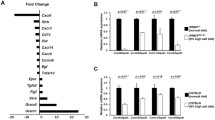
It has recently been revealed that several proinflammatory cytokines affect the activity and gene expression of renal tubular K+ channels. As shown in Fig. 1, their effects on K+ channels may be involved in alterations of tubular transport activity or exacerbation of renal cell injury. However, our understanding is still insufficient, with regard to their physiological and pathological significances in modulating renal tubular K+ channels. Multiple cytokines are produced during inflammation. Some cytokines might possess biphasic effects on their target molecules. Furthermore, different cytokines would exert opposite effects on the same targets. Such complicated modes of action give us profound difficulties in interpreting the final outcome of the effects of proinflammatory cytokines. Further studies are required to solve these problems.

Schematic representation showing the current knowledge of the effects of proinflammatory cytokines on the activity of the renal tubular K+ channels. The suppressive effects of cytokines on K+ channel gene expression are omitted. Dotted lines indicate putative effects. Circled plus and circled minus denote stimulation and suppression, respectively
References
Giebisch G, Krapf R, Wagner C. Renal and extrarenal regulation of potassium. Kidney Int. 2007;72:397–410.
Hebert SC, Desir G, Giebisch G, Wang W. Molecular diversity and regulation of renal potassium channels. Physiol Rev. 2005;85:319–71.
Feghali CA, Wright TM. Cytokines in acute and chronic inflammation. Front Biosci. 1997;2:d12–26.
Schäfers M, Sorkin L. Effect of cytokines on neuronal excitability. Neurosci Lett. 2008;437:188–93.
Allan SM, Tyrrell PJ, Rothwell NJ. Interleukin-1 and neuronal injury. Nat Rev Immunol. 2005;5:629–40.
Eisenhut M. Changes in ion transport in inflammatory disease. J Inflamm. 2006;3:5.
Daha MR, van Kooten C. Is the proximal tubular cell a proinflammatory cell? Nephrol Dial Transplant. 2000;15(Suppl 6):41–3.
Rangan GK, Wang Y, Tay YC, Harris DC. Differential effects of albumin on cytokine gene expression in proximal tubular epithelial cells. Nephrol Dial Transplant. 2005;20:1013–4.
Zager RA, Johnson AC, Geballe A. Gentamicin suppresses endotoxin-driven TNF-alpha production in human and mouse proximal tubule cells. Am J Physiol Renal Physiol. 2007;293:F1373–80.
Escalante BA, Ferreri NR, Dunn CE, McGiff JC. Cytokines affect ion transport in primary cultured thick ascending limb of Henle’s loop cells. Am J Physiol Cell Physiol. 1994;266:C1568–76.
Ferreri NR, Escalante BA, Zhao Y, An SJ, McGiff JC. Angiotensin II induces TNF production by the thick ascending limb: functional implications. Am J Physiol. 1998;274:F148–55.
Sakairi Y, Ando Y, Tabei K, Kusano E, Asano Y. Interleukin-1 inhibits sodium and water transport in rabbit cortical collecting duct. Am J Physiol. 1994;266:F674–80.
Kreydiyyeh SI, Al-Sadi R. Interleukin-1β increases urine flow rate and inhibits protein expression of Na+/K+-ATPase in the rat jejunum and kidney. J Interferon Cytokine Res. 2002;22:1041–8.
Kreydiyyeh SI, Al-Sadi R. The signal transduction pathway that mediates the effect of interleukin-1β on the Na+-K+-ATPase in LLC-PK1 cells. Pflügers Arch. 2004;448:231–8.
Vinciguerra M, Hasler U, Mordasini D, Roussel M, Capovilla M, Ogier-Denis E, et al. Cytokines and sodium induce protein kinase A-dependent cell-surface Na, K-ATPase recruitment via dissociation of NF-κB/IκB/protein kinase A catalytic subunit complex in collecting duct principal cells. J Am Soc Nephrol. 2005;16:2576–85.
Kreydiyyeh SI, Markossian S. Tumor necrosis factor α down-regulates the Na+-K+ ATPase and the Na+–K+–2Cl− cotransporter in the kidney cortex and medulla. Cytokine. 2006;33:138–44.
Schmidt C, Höcherl K, Schweda F, Kurtz A, Bucher M. Regulation of renal sodium transporters during severe inflammation. J Am Soc Nephrol. 2007;18:1072–83.
Viviani B, Gardoni F, Marinovich M. Cytokines and neuronal ion channels in health and disease. Int Rev Neurobiol. 2007;82:247–63.
Wei Y, Babilonia E, Pedraza PL, Ferreri NR, Wang WH. Acute application of TNF stimulates apical 70-pS K+ channels in the thick ascending limb of rat kidney. Am J Physiol Renal Physiol. 2003;285:F491–7.
Lu M, Wang T, Yan Q, Wang W, Giebisch G, Hebert SC. ROMK is required for expression of the 70-pS K channel in the thick ascending limb. Am J Physiol Renal Physiol. 2004;286:F490–5.
Wang WH. Regulation of ROMK (Kir1.1) channels: new mechanisms and aspects. Am J Physiol Renal Physiol. 2006;290:F14–9.
Gu RM, Wei Y, Falck JR, Krishna UM, Wang WH. Effects of protein tyrosine kinase and protein tyrosine phosphatase on apical K+ channels in the TAL. Am J Physiol Cell Physiol. 2001;281:C1188–95.
Wang WH. Two types of K+ channel in thick ascending limb of rat kidney. Am J Physiol. 1994;267:F599–605.
Lin DH, Sterling H, Yang B, Hebert SC, Giebisch G, Wang WH. Protein tyrosine kinase is expressed and regulates ROMK1 location in the cortical collecting duct. Am J Physiol Renal Physiol. 2004;286:F881–92.
Sterling H, Lin DH, Chen YJ, Wei Y, Wang ZJ, Lai J, et al. PKC expression is regulated by dietary K intake and mediates internalization of SK channels in the CCD. Am J Physiol Renal Physiol. 2004;286:F1072–8.
Nakamura K, Komagiri Y, Kojo T, Kubokawa M. Effects of cytokines on activity of an inwardly rectifying K+ channel in cultured human proximal tubule cells. J Iwate Med Assoc. 2007;59:375–85.
Nakamura K, Komagiri Y, Kojo T, Kubokawa M. Delayed and acute effects of interferon-γ on activity of an inwardly rectifying K+ channel in cultured human proximal tubule cells. Am J Physiol. 2009;296:F46–53.
Nakamura K, Hirano J, Kubokawa M. An ATP-regulated and pH-sensitive inwardly rectifying K+ channel in cultured human proximal tubule cells. Jpn J Physiol. 2001;51:523–30.
Nakamura K, Hirano J, Kubokawa M. Regulation of an inwardly rectifying K+ channel by nitric oxide in cultured human proximal tubule cells. Am J Physiol Renal Physiol. 2004;287:F411–7.
Nakamura K, Hirano J, Itazawa S, Kubokawa M. Protein kinase G activates inwardly rectifying K+ channel in cultured human proximal tubule cells. Am J Physiol Renal Physiol. 2002;283:F784–91.
Chatterjee PK, Hawksworth GM, McLay JS. Cytokine-stimulated nitric oxide production in human renal proximal tubule and its modulation by natriuretic peptides: a novel immunomodulatory mechanism? Exp Nephrol. 1999;7:438–48.
Kubokawa M, Kojo T, Komagiri Y, Nakamura K. Role of calcineurin-mediated dephosphorylation in modulation of an inwardly rectifying K+ channel in human proximal tubule cells. J Membr Biol. 2009;231:79–92.
Plata-Salaman CR, ffrench-Mullen JM. Interleukin-1 β inhibits Ca2+ channel currents in hippocampal neurons through protein kinase C. Eur J Pharmacol. 1994;266:1–10.
Choi JY, Choi YS, Kim SJ, Son EJ, Choi HS, Yoon JH. Interleukin-1β suppresses epithelial sodium channel β-subunit expression and ENaC-dependent fluid absorption in human middle ear epithelial cells. Eur J Pharmacol. 2007;567:19–25.
Cafferata EG, Gonzalez-Guerrico AM, Giordano L, Pivetta OH, Santa-Coloma TA. Interleukin-1β regulates CFTR expression in human intestinal T84 cells. Biochim Biophys Acta. 2000;1500:241–8.
Schmidt C, Höcherl K, Schweda F, Bucher M. Proinflammatory cytokines cause down-regulation of renal chloride entry pathways during sepsis. Crit Care Med. 2007;35:2110–9.
Schmidt C, Höcherl K, Bucher M. Regulation of renal glucose transporters during severe inflammation. Am J Physiol Renal Physiol. 2007;292:F804–11.
Schmidt C, Höcherl K, Bucher M. Cytokine-mediated regulation of urea transporters during experimental endotoxemia. Am J Physiol Renal Physiol. 2007;292:F1479–89.
Liu HJ, Wei Y, Fererri NR, Nasjletti A, Wang WH. Vasopressin and PGE2 regulate activity of apical 70 pS K+ channel in thick ascending limb of rat kidney. Am J Physiol Cell Physiol. 2000;278:C905–13.
Jonassen TE, Graebe M, Promeneur D, Nielsen S, Christensen S, Olsen NV. Lipopolysaccharide-induced acute renal failure in conscious rats: effects of specific phosphodiesterase type 3 and 4 inhibition. J Pharmacol Exp Ther. 2002;303:364–74.
Schrier RW, Wang W. Acute renal failure and sepsis. N Engl J Med. 2004;351:159–69.
Jonasch E, Haluska FG. Interferon in oncological practice: review of interferon biology, clinical applications, and toxicities. Oncologist. 2001;6:34–55.
Vesey DA, Cheung C, Endre Z, Gobé G, Johnson DW. Role of protein kinase C and oxidative stress in interleukin-1beta-induced human proximal tubule cell injury and fibrogenesis. Nephrology. 2005;10:73–80.
Phillips TM. Interferon-α induces renal dysfunction and injury. Curr Opin Nephrol Hypertens. 1996;5:380–3.
Nietsch HH, Roe MW, Fiekers JF, Moore AL, Lidofsky SD. Activation of potassium and chloride channels by tumor necrosis factor alpha. Role in liver cell death. J Biol Chem. 2000;275:20556–61.
Reeves WB, Shah SV. Activation of potassium channels contributes to hypoxic injury in proximal tubules. J Clin Invest. 1994;94:2289–94.
Engbersen R, Moons MM, Wouterse AC, Dijkman HB, Kramers C, Smits P, et al. Sulphonylurea drugs reduce hypoxic damage in the isolated perfused rat kidney. Br J Pharmacol. 2000;130:1678–84.
Pompermayer K, Souza DG, Lara GG, Silveira KD, Cassali GD, Andrade AA, et al. The ATP-sensitive potassium channel blocker glibenclamide prevents renal ischemia/reperfusion injury in rats. Kidney Int. 2005;67:1785–96.
Rahgozar M, Willgoss DA, Gobé GC, Endr ZH. ATP-dependent K+ channels in renal ischemia reperfusion injury. Ren Fail. 2003;25:885–96.
Zager RA, Johnson AC, Lund S, Hanson SY, Abrass CK. Levosimendan protects against experimental endotoxemic acute renal failure. Am J Physiol Renal Physiol. 2006;290:F1453–62.
Assad AR, Delou JM, Fonseca LM, Villela NR, Nascimento JH, Verçosa N, et al. The role of KATP channels on propofol preconditioning in a cellular model of renal ischemia-reperfusion. Anesth Analg. 2009;109:1486–92.
Penning LC, Denecker G, Vercammen D, Declercq W, Schipper RG, Vandenabeele P. A role for potassium in TNF-induced apoptosis and gene-induction in human and rodent tumour cell lines. Cytokine. 2000;12:747–50.
Lang F, Shumilina E, Ritter M, Gulbins E, Vereninov A, Huber SM. Ion channels and cell volume in regulation of cell proliferation and apoptotic cell death. Contrib Nephrol. 2006;152:142–60.
Author information
Authors and Affiliations
Corresponding author
About this article
Cite this article
Nakamura, K., Komagiri, Y. & Kubokawa, M. Effects of cytokines on potassium channels in renal tubular epithelia. Clin Exp Nephrol 16, 55–60 (2012). https://doi.org/10.1007/s10157-011-0490-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10157-011-0490-5