
Abstract
In this study, we used an automated segmentation of regions of interest and co-registration to diffusion tensor imaging (DTI) images to investigate whether microstructural abnormalities occur in gray structures of the frontal-subcortical circuits in patients with amyotrophic lateral sclerosis (ALS). Twenty-four patients with probable or definite sporadic ALS and 22 healthy controls were enrolled in the study. Thirteen out of 24 ALS patients and all of the control subjects underwent a detailed neuropsychological evaluation. DTI was performed to measure mean diffusivity (MD) and fractional anisotropy in the frontal cortex, caudate, putamen, globus pallidus, thalamus, amygdala and hippocampus. MD values of ALS patients were significantly higher in the frontal cortex (P = 0.023), caudate (P = 0.01), thalamus (P = 0.019), amygdala (P = 0.012) and hippocampus (P = 0.002) compared to controls. MD of these structures significantly correlated to a variable degree with neurological disability and neuropsychological dysfunctions. The increased MD values in several cortical and subcortical gray structures and their correlations with neuropsychological variables substantiate a multisystemic degeneration in ALS and suggest that dysfunctions of frontal–subcortical circuits could play a pivotal role in frontal impairment and behavioral symptoms in ALS patients.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Amyotrophic lateral sclerosis (ALS) is caused by a degeneration of lower motor and pyramidal neurons [1], leading to loss of voluntary muscle movements. The diagnosis of ALS is based on clinical features, findings on electrodiagnostic testing and exclusion of other health conditions. Although ALS has traditionally been considered a paradigm of a pure motor neuron disorder, previous pathological studies have described extra-motor alterations [2, 3] as a potential contributor of the disease. Several studies have demonstrated dysfunctions of the non-motor cortex (prefrontal and temporal cortices) [4, 5] and a widespread neuronal degeneration in many subcortical gray matter (GM) structures (thalamus, subthalamic nucleus and cerebellum) [6]. Therefore, the presence of multisystemic neurodegenerative processes in ALS may explain why many ALS patients display, along with the typical motor deficits, neuropsychological dysfunctions. Diffusion tensor imaging (DTI) is one of the most sensitive methods for detecting alterations of cerebral tissues [7]. The diffusion behavior of water molecules can be evaluated via indices such as mean diffusivity (MD), which measures the magnitude of diffusion, and fractional anisotropy (FA), which quantifies the preferential direction of water diffusion along fiber tracts, reflecting the degree of alignment of cellular structures within white matter (WM) [8]. The majority of previous DTI studies in ALS have focused on the assessment of WM damage, especially of the corticospinal tract [9–11] and corpus callosum [12–14]. Few DTI studies have investigated alterations of cortical GM (precentral gyrus, inferior frontal gyrus, middle temporal gyrus, temporal pole, postcentral gyrus, angular gyrus, supramarginal gyrus) [15] and subcortical GM structures (basal ganglia, thalamus) [16] in ALS patients using DTI.
However, in vitro studies [2, 3, 17–19] on ALS patients have demonstrated a degeneration of GM structures that are functionally implicated in frontal-subcortical circuits [20], such as the frontal cortex, basal ganglia, thalamus, hippocampus and amygdala. Consequently, the in vivo investigation of these circuits could help clarify the pathophysiology of ALS-related cognitive impairment that is still unclear.
The main objective of this study is to evaluate the involvement of cortical and subcortical GM structures of the frontal-subcortical circuits in ALS patients using DTI. Our primary hypothesis is that DTI measurements in these GM structures are different in patients with ALS than in age-matched controls; the secondary hypothesis is that DTI alterations correlate with clinical variables and frontal dysfunctions.
Methods
Subjects
From February 2010 to May 2012, we enrolled 26 consecutive patients with sporadic ALS and 23 age-matched healthy controls. Two of the patients and one of the healthy controls showed structural abnormalities (such as vascular lesions) upon conventional magnetic response imaging (MRI) and were, therefore, not included in the study. The data of the remaining 24 patients (13 males and 11 females; mean age 61.7 ± 11.4 years, range 42–88, median 62; disease duration 23.0 ± 18.5 months, range 10–76, median 12; 15 with limb onset, four with bulbar onset and five with limb and bulbar onset) and 22 age-matched healthy controls (10 males and 12 females; mean age 60.1 ± 9.8 years; range 41–82; median 59.5) were analyzed. All participants gave written informed consent, which was approved by the ethical committee of the University “Magna Graecia” of Catanzaro, Italy.
Clinical assessments
Clinical diagnosis of ALS was made by one of the authors who was blinded to the MRI results. A detailed medical history and clinical examination were conducted in all patients. The patients were classified as follows: 14 as definite ALS and 10 as probable ALS according to the revised El Escorial research diagnostic criteria [21]. We considered as exclusion criteria the presence of multifocal motor neuropathy and paraneoplastic neuropathy, using nerve conduction studies. The disease severity was evaluated using the ALS functional rating scale-revised (ALSFRS-R) [22].
Cognitive functions were evaluated in 13 out of 24 ALS patients (seven males and six females; mean age 59.9 ± 13.5 years, range 42–88, median 61; disease duration 25.3 ± 18.2 months, range 12–65, median 24) and in all of control subjects by the following tests: Mini Mental State Examination (MMSE) [23]; Beck Depression Inventory-II (BDI-II) [24]; Rey Auditory Verbal Learning Test-Immediate Recall (RAVLT-IR); Rey Auditory Verbal Learning Test-Delayed Recall (RAVLT-DR) [25]; Controlled Oral Word Association Test (COWAT) [26]; Modified Card Sorting Test (MCST) [27] and Frontal Assessment Battery (FAB) [28].
The cognitive evaluation of 11 ALS patients (six males and five females; mean age 63.8 ± 8.4 years, range 48–78, median 65; disease duration 20.4 ± 19.5 months, range 10–76, median 12) was not included in the study: the motor disabilities (severe dysarthria and upper limbs weakness) of these patients affected neuropsychological scores and did not allow us to perform reliable tests.
MRI protocol and image processing
Subjects were examined using a 3-Tesla MR750 GE MRI scanner with an eight channel head coil. All participants underwent the same MRI protocol, including conventional T1-weighted, T2-weighted and FLAIR scanning. Whole-brain 3D T1-weighted SPGR (BRAVO) images were obtained in the sagittal plane with a voxel size of 1 × 1 × 1 mm3. Diffusion-weighted volumes were acquired using spin-echo planar imaging (matrix size 128 × 128; 80 axial slices, voxel size 2 × 2 × 2 mm3, 2 NEX) with 27 isotropically distributed orientations for the diffusion-sensitizing gradients at a b value of 1.000 s × mm2 and four b = 0 images. Image processing was performed with FSL 5.0 (http://www.fmrib.ox.ac.uk/fsl/) using a protocol described previously [29]. After correction for image distortions and head motion, a diffusion tensor model was fit at each voxel, generating FA and MD maps. For registering DTI data to the T1-weighted anatomic image, we calculated a full-affine (correlation ratio cost function) transformation between FA maps and brain-extracted whole-brain volumes from T1-weighted images. The calculated transformation matrix was then applied to the MD maps with identical resampling options.
Anatomic T1-weighted images were processed with the segmentation tool FIRST 5.0 integrated within the FSL software. For each subject and each hemisphere, the following regions were identified: caudate, putamen, globus pallidus, thalamus, hippocampus, amygdala, frontal cortex. Results of regions of interest (ROI) segmentation were superimposed on anatomic images and visually inspected by a trained radiologist to exclude misregistration or erroneous ROI identification. The segmented regions defined the binary masks where mean values of MD and FA were calculated for each individual.
Statistical analysis
The difference in sex distribution between patients with ALS and control subjects was evaluated with χ 2 test. The differences in continuous clinical and imaging variables between the study groups were assessed using two-tailed, two-sample t test. Pearson’s correlation analysis was used to evaluate the relationship between DTI parameters and clinical and cognitive parameters. Statistical analysis for clinical and imaging data was performed with Statistical Package for Social Science Software (SPSS, version 12.0, Chicago, IL, USA) for Windows. Consistent with the primary and secondary hypotheses, the P value corrected for multiple comparisons (Bonferroni correction) was set to <0.025 for identifying significant differences in MD and FA between the groups and for evaluating significant correlations of DTI alterations with clinical variables (disease duration and ALSFRS-R score) and frontal dysfunctions (MCST and FAB scores).
Results
Clinical features
The sex (χ 2 = 0.55) and age (P = 0.60) distributions did not differ between groups. In ALS patients, we evaluated the degree of motor disability with ALSFRS-R (score 27.4 ± 6.3, range 17–39, median 28). When we compared the group of 13 ALS patients with cognitive evaluation and control subjects, we found no significant differences in sex (χ 2 = 0.63), age (P = 0.60), education (P = 0.15), MMSE (8.9 ± 4.0 vs. 10.8 ± 3.5, P = 0.17), RAVLT-DR (5.9 ± 3.4 vs. 7.9 ± 2.7, P = 0.076), or BDI-II (12.6 ± 7.1 vs. 8.0 ± 6.5, P = 0.055), and significant differences in RAVLT-IR (34.3 ± 11.4 vs. 41.0 ± 6.6, P = 0.035), COWAT (17.6 ± 9.6 vs. 26.3 ± 9.2, P = 0.012), MCST (4.4 ± 1.8 vs. 5.9 ± 0.3, P = 0.001) and FAB (13.3 ± 3.4 vs. 15.6 ± 1.8, P = 0.016). The patients with cognitive assessment did not statistically differ from those without cognitive assessment in terms of demographics and clinical features.
Diffusion tensor imaging
Table 1 shows DTI mean values. No left versus right asymmetry of DTI measures was found in either controls or ALS patients. Therefore, data from the left and right sides in each group were averaged for all the subsequent group comparisons. MD was significantly higher in patients than in controls in the frontal cortex (P = 0.023), caudate (P = 0.01), thalamus (P = 0.019), hippocampus (P = 0.002) and amygdala (P = 0.012). No significant differences were found in MD values of the putamen or globus pallidus. No significant differences in FA values were found in any of the structures investigated.
Correlation analyses between the clinical features of 24 ALS patients and MD values (Table 2) revealed a significant positive correlation between disease duration and MD of the caudate (r = 0.56, P = 0.004), and thalamus (r = 0.48, P = 0.02) and frontal cortex (r = 0.49, P = 0.01). The ALSFRS-R score correlated negatively with the MD of the thalamus (r = −0.47, P = 0.02), amygdala (r = −0.48, P = 0.02) and frontal cortex (r = −0.62, P = 0.001).
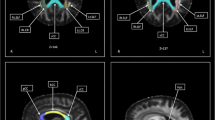
Correlation analyses between neuropsychological test scores of 13 ALS patients and MD measures revealed several negative correlations (Table 3). The MCST scores correlated with the MD values of the caudate (r = −0.71, P = 0.007; Fig. 1), hippocampus (r = −0.81, P < 0.001), amygdala (r = −0.66, P = 0.015; Fig. 1) and frontal cortex (r = −0.78, P = 0.002; Fig. 1). The FAB scores correlated with the MD values of the caudate (r = −0.64, P = 0.017; Fig. 1), hippocampus (r = −0.81, P = 0.001), amygdala (r = −0.77, P = 0.002; Fig. 1) and frontal cortex (r = −0.69, P = 0.009; Fig. 1).

Correlations between performances at the neuropsychological tests (MCST, and FAB) and MD of the frontal cortex, amygdala and caudate in patients with ALS. For illustrative purposes, the left column shows the reconstructions of the GM structures in a single healthy control (the ROIs are superimposed onto the T1-weighted images). The right column shows the scatterplots of the correlations; MD values are reported on the x-axis, frontal functions scores are reported on the y-axis (r correlation coefficient, P = p value, corrected for multiple comparisons with cut-off set to <0.025)
Discussion
In the present study, we evaluated brain areas implicated in frontal-subcortical circuits, such as the frontal cortex, basal ganglia, thalamus, hippocampus and amygdala. In general, brain pathological processes that modify tissue integrity reduce the barriers that restrict the movement of water molecules. As a consequence, MD increases and FA decreases [30].
ALS patients showed significant increases of MD values compared to healthy controls in the cortical (frontal cortex and hippocampus) and subcortical (caudate, thalamus and amygdala) GM structures, without significant differences in FA values. FA is not a satisfactory marker to evaluate alterations of GM, while it is more effective for analyzing WM [8]. FA is intrinsically higher in white matter than in gray matter, where water diffusion shows less directional dependence [31]. The low amount of anisotropic structures (i.e., WM fibers) in the considered ROIs may contribute to making FA a less sensitive indicator of neurodegeneration than MD in these GM structures.
The degeneration of the frontal cortex found in our ALS patients has been widely demonstrated by precedent neuroimaging studies [15, 32, 33], suggesting a continuum between ALS and frontotemporal dementia (FTD). Also DTI abnormalities observed in the caudate and thalamus of our patients support the results from previous in vitro pathological investigations [2, 3] and in vivo imaging studies [16, 34], suggesting involvement of these subcortical structures in ALS. Finally, we showed a microstructural damage of the hippocampus and amygdala, using an advanced DTI technique, confirming precedent pathological studies [17, 18]. High MD values may be the consequence of underlying pathological changes that may include the presence of ubiquitin-immunoreactive neural inclusions [19, 35] and pathological TDP-43 lesions [36, 37]. We speculate that diffuse neurodegeneration of these cortical and subcortical regions, including hippocampus, basal ganglia and amygdala, may modify tissue integrity and contribute to the damage of structural barriers at cellular and sub-cellular levels, increasing the local diffusivity of water molecules [38].
Regarding the correlation that we found between disease duration and MD values of the frontal cortex, thalamus and caudate, and between ALSFRS-R and MD values of the frontal cortex, thalamus and amygdala, our study supports the view that ALS is a degenerative multi-systemic pathology. In agreement with some previous results [32, 39, 40], our correlation analysis showed that a microstructural damage of frontal cortex was significantly related to disease duration and disability. Moreover, we found that MD values of several subcortical structures were related with disease duration (i.e., thalamus and caudate) and disability (i.e., thalamus and amygdala). These results are a further evidence of a possible role of extra-motor subcortical degeneration as marker of disease progression in ALS.
We hypothesize that the microstructural alterations of these extra-motor structures might have implications on the pathophysiology of behavior changes and cognitive impairment found in ALS [4, 41, 42], through a dysfunction of frontal-subcortical circuits. Each of these circuits shares a common structure, linking specific areas of the frontal cortex (dorsolateral, anterior cingulated and orbitofrontal cortex) to the striatum, basal ganglia and thalamus (closed-loops) [20]. In particular, the dorsolateral circuit mediates executive functions, and the orbitofrontal circuit is involved in emotional and behavioral inhibition. Both circuits have afferent and efferent connections with the amygdala, which play an important role in functional integration [43]. Although each frontal-subcortical circuit constitutes a closed loop of anatomically segregated dedicated neurons, “open”-loop elements are incorporated into the functional connectivity of these circuits [44]. The connections with other cortical structures (e.g., parietal and temporal cortices) and other deep nuclei (e.g., amygdala and hippocampus) [45] are dedicated to memory and language.
Previous DTI studies have already highlighted microstructural damage of the frontal WM tracts that link the subcortical nuclei to the frontal cortex (e.g., anterior corpus callosum [12, 13], uncinate fasciculus [12, 46–48], prefrontal WM regions [12, 49, 50] and bilateral frontal WM/cingulate gyrus [48, 51]). These precedent results support our observations, because we demonstrated the involvement of both subcortical gray nuclei and the frontal cortex in ALS patients.
According to recent studies that pointed out the features of cognitive impairment in ALS [52, 53], our patients showed a significant difference with respect to control subjects in terms of verbal fluency (COWAT scores) [54, 55] and frontal functions (MCST and FAB scores) [56, 57]. In particular, the MCST and FAB tests were used to evaluate cognitive and behavioral domains under the control of the frontal lobes (executive functions, behavioral regulation and response initiation) [58].
Furthermore, the negative correlations between the measures of the frontal functions (MCST and FAB scores) and MD values of the frontal cortex, caudate and amygdala suggest that dysfunctions of frontal-subcortical circuits could have a pivotal function in ALS-related disinhibition and dysexecutive syndrome [59].
Using voxel-based morphometry (VBM), recent studies [60, 61] showed a relationship between cortical density and cognitive/behavioral dysfunctions in ALS, establishing that cortical atrophy in ALS is highly dependent on cognitive changes [60] and demonstrating a neural damage (anterior cingulate cortex and right inferior frontal gyrus) in a limbic prefrontal network [61]. In our study, we used an automated segmentation of ROIs and coregistration to DTI images to investigate not cortical density changes, but the presence of microstructural abnormalities in subcortical structures, namely basal ganglia, thalamus, hippocampus and amygdala. The correlation between MD values and the measures of the frontal functions confirms the hypothesis of microstructural damage that involve not only the frontal cortex, but also the deep nuclei of the frontal-subcortical networks.
This hypothesis of dysfunction within frontal-subcortical circuits of ALS patients was confirmed in the case of the limbic system, in a recent fMRI-study [62]. Relative to healthy controls, ALS patients showed greater activation in several prefrontal areas (ventral and dorsal anterior cingulated cortex and dorsolateral prefrontal cortex) and altered connectivity between left amygdala and prefrontal cortex. Moreover, the authors reported that altered connectivity of the left amygdala and supplementary motor area was related to greater disease severity in ALS patients, substantiating limbic-motor interface abnormalities in ALS.
There is disagreement about memory deficits in ALS. Studies of cognition have shown that memory impairments in patients with ALS usually involve immediate recall [55, 63]. Deficits in delayed recall are highly variable [53, 55]. According to these precedent studies, we found significant differences only in RAVLT-IR scores, and not in RAVLT-DR scores.
Given that mood disturbances, such as depression, could have affected the results of neuropsychological tests, we compared BDI-II mean scores of ALS patients and control group, but we found no significant differences. These results are consistent with recent reports [64–66] that claim that prevalence rates of depression are low in ALS and exclude an influence of depression on tests that evaluate the frontal functions.
Furthermore, alongside the dysexecutive symptoms, apathy appears to be a common behavioral abnormality in ALS patients [67, 68]; recent neuroimaging studies suggest that apathy could be correlated with a disruption of cortical-basal ganglia circuits in both ALS [69] and FTD [70]. Further studies that would require detailed neuropsychological assessment of apathy and neuroimaging examination of frontal-subcortical circuits are needed to more clearly delineate the anatomical and functional correlates of apathy in ALS.
The major limitation of the present study lies in the limited number of patients who underwent complete neuropsychological tests. In fact, the motor disabilities of some patients did not allow us to perform exhaustive and detailed tests. Additional studies with larger sample sizes are necessary to evaluate the degree, extent and pattern of degeneration of these structures. Other structural and functional neuroimaging studies could be necessary to confirm our results.
However, the present study highlights the large potential of DTI of the brain to provide in vivo markers of cortical and subcortical involvement in ALS. Our findings of diffusion abnormalities in the frontal cortex, caudate, thalamus, hippocampus and amygdala of patients with ALS suggest that there is degeneration or dysfunction of neurons in these extra-motor structures. This novel aspect is particularly promising for understanding the pathophysiology of executive impairment found in ALS and for studying what the underlying neuroanatomical changes are from the point of establishing a common framework for ALS and FTD.
References
Rowland LP (1998) Diagnosis of amyotrophic lateral sclerosis. J Neurol Sci 160(Suppl 1):6–24
Smith MC (1960) Nerve fibre degeneration in the brain in amyotrophic lateral sclerosis. J Neurol Neurosurg Psychiatry 23:269–282
Bigio EH, Lipton AM, White CL 3rd, Dickson DW, Hirano A (2003) Frontotemporal and motor neurone degeneration with neurofilament inclusion bodies: additional evidence for overlap between FTD and ALS. Neuropathol Appl Neurobiol 29:239–253
Ringholz GM, Appel SH, Bradshaw M, Cooke NA, Mosnik DM, Schulz PE (2005) Prevalence and pattern of cognitive impairment in sporadic amyotrophic lateral sclerosis. Neurology 65:586–590
Talbot PR, Goulding PJ, Lloyd JJ, Snowden JS, Neary D, Testa HJ (1995) Inter-relation between “classic” motor neuron disease and frontotemporal dementia: neuropsychological and single photon emission computed tomography study. J Neurol Neurosurg Psychiatry 58:541–547
Thivard L, Pradat PF, Lehéricy S, Lacomblez L, Dormont D, Chiras J, Benali H, Meininger V (2007) Diffusion tension imaging and voxel based morphometry study in amyotrophic lateral sclerosis: relationships with motor disability. J Neurol Neurosurg Psychiatry 78:889–892
Chenevert TL, Brunberg JA, Pipe JG (1990) Anisotropic diffusion in human white matter: demonstration with MR techniques in vivo. Radiology 177:401–405
Basser PJ, Pierpaoli C (1996) Microstructural and physiological features of tissues elucidated by quantitative-diffusiontensor MRI. J Magn Reson B 111:209–219
Ellis CM, Simmons A, Jones DK, Bland J, Dawson JM, Horsfield MA, Williams SC, Leigh PN (1999) Diffusion tensor MRI assesses corticospinal tract damage in ALS. Neurology 53:1051–1058
Graham JM, Papadakis N, Evans J, Widjaja E, Romanowski CA, Paley MN, Wallis LI, Wilkinson ID, Shaw PJ, Griffiths PD (2004) Diffusion tensor imaging for the assessment of upper motor neuron integrity in ALS. Neurology 63:2111–2119
Toosy AT, Werring DJ, Orrell RW, Howard RS, King MD, Barker GJ, Miller DH, Thompson AJ (2003) Diffusion tensor imaging detects corticospinal tract involvement at multiple levels in amyotrophic lateral sclerosis. J Neurol Neurosurg Psychiatry 74:1250–1257
Ciccarelli O, Behrens TE, Johansen-Berg H, Talbot K, Orrell RW, Howard RS, Nunes RG, Miller DH, Matthews PM, Thompson AJ, Smith SM (2009) Investigation of white matter pathology in ALS and PLS using tract-based spatial statistics. Hum Brain Mapp 30:615–624
Filippini N, Douaud G, Mackay CE, Knight S, Talbot K, Turner MR (2010) Corpus callosum involvement is a consistent feature of amyotrophic lateral sclerosis. Neurology 75:1645–1652
Cirillo M, Esposito F, Tedeschi G, Caiazzo G, Sagnelli A, Piccirillo G, Conforti R, Tortora F, Monsurrò MR, Cirillo S, Trojsi F (2012) Widespread microstructural white matter involvement in amyotrophic lateral sclerosis: a whole-brain DTI study. AJNR Am J Neuroradiol 33:1102–1108
Canu E, Agosta F, Riva N, Sala S, Prelle A, Caputo D, Perini M, Comi G, Filippi M (2011) The topography of brain microstructural damage in amyotrophic lateral sclerosis assessed using diffusion tensor MR imaging. AJNR Am J Neuroradiol 32:1307–1314
Sharma KR, Sheriff S, Maudsley A, Govind V (2013) Diffusion tensor imaging of basal ganglia and thalamus in amyotrophic lateral sclerosis. J Neuroimaging 23:368–374
Kawashima T, Doh-ura K, Kikuchi H, Iwaki T (2001) Cognitive dysfunction in patients with amyotrophic lateral sclerosis is associated with spherical or crescent-shaped ubiquitinated intraneuronal inclusions in the parahippocampal gyrus and amygdala, but not in the neostriatum. Acta Neuropathol 102:467–472
Takeda T, Uchihara T, Arai N, Mizutani T, Iwata M (2009) Progression of hippocampal degeneration in amyotrophic lateral sclerosis with or without memory impairment: distinction from Alzheimer disease. Acta Neuropathol 117:35–44
Anderson VE, Cairns NJ, Leigh PN (1995) Involvement of the amygdala, dentate and hippocampus in motor neuron disease. J Neurol Sci 129(Suppl):75–78
Alexander GE, DeLong MR, Strick PL (1986) Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci 9:357–381
Brooks BR, Miller RG, Swash M, Munsat TL, World Federation of Neurology Research Group on Motor Neuron Diseases (2000) El Escorial revisited: revised criteria for the diagnosis of amyotrophic lateral sclerosis. Amyotroph Lateral Scler Other Motor Neuron Disord 1:293–299
Cedarbaum JM, Stambler N, Malta E, Fuller C, Hilt D, Thurmond B, Nakanishi A (1999) The ALSFRS-R: a revised ALS functional rating scale that incorporates assessments of respiratory function. BDNF ALS Study Group (Phase III). J Neurol Sci 169:13–21
Folstein MF, Folstein SE, McHugh PR (1975) “Mini-mental state”. A practical method for grading the cognitive state of patients for the clinician. J Psychiatr Res 12:189–198
Beck AT, Steer RA, Brown GK (1996) Manual for beck depression inventory-II. Psychological Corporation, San Antonio
Rey A (1958) Memorisation d’une serie de 15 mots en 5 repetitions. Presses Universitaries des France, Paris
Benton A, Hamsher KD (1989) Multilingual aphasia examination. AJA Associates, Iowa City
Nelson HE (1976) A modified card sorting test sensitive to frontal lobe defects. Cortex 12:313–324
Dubois B, Slachevsky A, Litvan I, Pillon B (2000) The FAB: a frontal assessment battery at bedside. Neurology 55:1621–1626
Cherubini A, Péran P, Caltagirone C, Sabatini U, Spalletta G (2009) Aging of subcortical nuclei: microstructural, mineralization and atrophy modifications measured in vivo using MRI. Neuroimage 48:29–36
Nicoletti G, Lodi R, Condino F, Tonon C, Fera F, Malucelli E, Manners D, Zappia M, Morgante L, Barone P, Barbiroli B, Quattrone A (2006) Apparent diffusion coefficient measurements of the middle cerebellar peduncle differentiate the Parkinson variant of MSA from Parkinson’s disease and progressive supranuclear palsy. Brain 129:2679–2687
Pierpaoli C, Jezzard P, Basser PJ, Barnett A, Di Chiro G (1996) Diffusion tensor MR imaging of the human brain. Radiology 201:637–648
Senda J, Kato S, Kaga T, Ito M, Atsuta N, Nakamura T, Watanabe H, Tanaka F, Naganawa S, Sobue G (2011) Progressive and widespread brain damage in ALS: MRI voxel-based morphometry and diffusion tensor imaging study. Amyotroph Lateral Scler 12:59–69
Lillo P, Mioshi E, Burrell JR, Kiernan MC, Hodges JR, Hornberger M (2012) Grey and white matter changes across the amyotrophic lateral sclerosis-frontotemporal dementia continuum. PLoS ONE 7:e43993
Sharma KR, Saigal G, Maudsley AA, Govind V (2011) 1H MRS of basal ganglia and thalamus in amyotrophic lateral sclerosis. NMR Biomed 24:1270–1276
Al-Sarraj S, Maekawa S, Kibble M, Everall I, Leigh N (2002) Ubiquitin-only intraneuronal inclusion in the substantia nigra is a characteristic feature of motor neurone disease with dementia. Neuropathol Appl Neurobiol 28:120–128
Geser F, Martinez-Lage M, Robinson J, Uryu K, Neumann M, Brandmeir NJ, Xie SX, Kwong LK, Elman L, McCluskey L, Clark CM, Malunda J, Miller BL, Zimmerman EA, Qian J, Van Deerlin V, Grossman M, Lee VM, Trojanowski JQ (2009) Clinical and pathological continuum of multisystem TDP-43 proteinopathies. Arch Neurol 66:180–189
Geser F, Brandmeir NJ, Kwong LK, Martinez-Lage M, Elman L, McCluskey L, Xie SX, Lee VM, Trojanowski JQ (2008) Evidence of multisystem disorder in whole-brain map of pathological TDP-43 in amyotrophic lateral sclerosis. Arch Neurol 65:636–641
Le Bihan D, Breton E, Lallemand D, Grenier P, Cabanis E, Laval-Jeantet M (1986) MR imaging of intravoxel incoherent motions: application to diffusion and perfusion in neurologic disorders. Radiology 161:401–407
Verstraete E, Veldink JH, Hendrikse J, Schelhaas HJ, van den Heuvel MP, van den Berg LH (2012) Structural MRI reveals cortical thinning in amyotrophic lateral sclerosis. J Neurol Neurosurg Psychiatry 83:383–388
D’Ambrosio A, Gallo A, Trojsi F, Corbo D, Esposito F, Cirillo M, Monsurró MR, Tedeschi G (2013) Frontotemporal cortical thinning in amyotrophic lateral sclerosis. AJNR Am J Neuroradiol. doi:10.3174/ajnr.A3753
DeLong MR, Wichmann T (2007) Circuits and circuit disorders of the basal ganglia. Arch Neurol 64:20–24
Strong MJ, Grace GM, Freedman M, Lomen-Hoerth C, Woolley S, Goldstein LH, Murphy J, Shoesmith C, Rosenfeld J, Leigh PN, Bruijn L, Ince P, Figlewicz D (2009) Consensus criteria for the diagnosis of frontotemporal cognitive and behavioural syndromes in amyotrophic lateral sclerosis. Amyotroph Lateral Scler 10:131–146
Cummings JL (1995) Anatomic and behavioral aspects of frontal-subcortical circuits. Ann N Y Acad Sci 15(769):1–13
Bonelli RM, Cummings JL (2007) Frontal-subcortical circuitry and behavior. Dialogues Clin Neurosci 9:141–151
Tekin S, Cummings JL (2002) Frontal-subcortical neuronal circuits and clinical neuropsychiatry: an update. J Psychosom Res 53:647–654
Agosta F, Pagani E, Petrolini M, Caputo D, Perini M, Prelle A, Salvi F, Filippi M (2010) Assessment of white matter tract damage in patients with amyotrophic lateral sclerosis: a diffusion tensor MR imaging tractography study. AJNR Am J Neuroradiol 31:1457–1461
Sato K, Aoki S, Iwata NK, Masutani Y, Watadani T, Nakata Y, Yoshida M, Terao Y, Abe O, Ohtomo K, Tsuji S (2010) Diffusion tensor tract-specific analysis of the uncinate fasciculus in patients with amyotrophic lateral sclerosis. Neuroradiology 52:729–733
Sarro L, Agosta F, Canu E, Riva N, Prelle A, Copetti M, Riccitelli G, Comi G, Filippi M (2011) Cognitive functions and white matter tract damage in amyotrophic lateral sclerosis: a diffusion tensor tractography study. AJNR Am J Neuroradiol 32:1866–1872
Sage CA, Peeters RR, Görner A, Robberecht W, Sunaert S (2007) Quantitative diffusion tensor imaging in amyotrophic lateral sclerosis. Neuroimage 34:486–499
Tsujimoto M, Senda J, Ishihara T, Niimi Y, Kawai Y, Atsuta N, Watanabe H, Tanaka F, Naganawa S, Sobue G (2011) Behavioral changes in early ALS correlate with voxel-based morphometry and diffusion tensor imaging. J Neurol Sci 307:34–40
Li J, Pan P, Song W, Huang R, Chen K, Shang H (2012) A meta-analysis of diffusion tensor imaging studies in amyotrophic lateral sclerosis. Neurobiol Aging 33:1833–1838
Phukan J, Pender NP, Hardiman O (2007) Cognitive impairment in amyotrophic lateral sclerosis. Lancet Neurol 6:994–1003
Christidi F, Zalonis I, Smyrnis N, Evdokimidis I (2012) Selective attention and the three-process memory model for the interpretation of verbal free recall in amyotrophic lateral sclerosis. J Int Neuropsychol Soc 18:809–818
Ludolph AC, Langen KJ, Regard M, Herzog H, Kempo B, Kurwert T (1992) Frontal lobe function in amyotrophic lateral sclerosis: a neuropsychologic and positron emission tomography study. Acta Neurol Scand 85:81–89
Gallassi R, Montagna P, Ciardulli C, Lorusso S, Mussuto V, Stracciari A (1985) Cognitive impairment in motor neuron disease. Acta Neurol Scand 71:480–484
Moretti R, Torre P, Antonello RM, Carraro N, Cazzato G, Bava A (2002) Complex cognitive disruption in motor neuron disease. Dement Geriatr Cogn Disord 14:141–150
Ahn SW, Kim SH, Kim JE, Kim SM, Kim SH, Sung JJ, Lee KW, Hong YH (2011) Frontal assessment battery to evaluate frontal lobe dysfunction in ALS patients. Can J Neurol Sci 38:242–246
Stukovnik V, Zidar J, Podnar S, Repovs G (2010) Amyotrophic lateral sclerosis patients show executive impairments on standard neuropsychological measures and an ecologically valid motor-free test of executive functions. J Clin Exp Neuropsychol 32:1095–1109
Achi EY, Rudnicki SA (2012) ALS and frontotemporal dysfunction: a review. Neurol Res Int 2012:806306
Mioshi E, Lillo P, Yew B, Hsieh S, Savage S, Hodges JR, Kiernan MC, Hornberger M (2013) Cortical atrophy in ALS is critically associated with neuropsychiatric and cognitive changes. Neurology 80:1117–1123
Cerami C, Dodich A, Canessa N, Crespi C, Iannaccone S, Corbo M, Lunetta C, Consonni M, Scola E, Falini A, Cappa SF (2013) Emotional empathy in amyotrophic lateral sclerosis: a behavioural and voxel-based morphometry study. Amyotroph Lateral Scler Frontotemporal Degener. doi:10.3109/21678421.2013.785568
Passamonti L, Fera F, Tessitore A, Russo A, Cerasa A, Gioia CM, Monsurrò MR, Migliaccio R, Tedeschi G, Quattrone A (2013) Dysfunctions within limbic-motor networks in amyotrophic lateral sclerosis. Neurobiol Aging 34:2499–2509
Massman PJ, Sims J, Cooke N, Haverkamp LJ, Appel V, Appel SH (1996) Prevalence and correlates of neuropsychological deficits in amyotrophic lateral sclerosis. J Neurol Neurosurg Psychiatry 61:450–455
Averill AJ, Kasarskis EJ, Segerstrom SC (2007) Psychological health in patients with amyotrophic lateral sclerosis. Amyotroph Lateral Scler 8:243–254
Atassi N, Cook A, Pineda CM, Yerramilli-Rao P, Pulley D, Cudkowicz M (2011) Depression in amyotrophic lateral sclerosis. Amyotroph Lateral Scler 12:109–112
Jelsone-Swain L, Persad C, Votruba KL, Weisenbach SL, Johnson T, Gruis KL, Welsh RC (2012) The relationship between depressive symptoms, disease state, and cognition in amyotrophic lateral sclerosis. Front Psychol 3:542
Grossman AB, Woolley-Levine S, Bradley WG, Miller RG (2007) Detecting neurobehavioral changes in amyotrophic lateral sclerosis. Amyotroph Lateral Scler 8:56–61
Gibbons ZC, Richardson A, Neary D, Snowden JS (2008) Behaviour in amyotrophic lateral sclerosis. Amyotroph Lateral Scler 9:67–74
Woolley SC, Zhang Y, Schuff N, Weiner MW, Katz JS (2011) Neuroanatomical correlates of apathy in ALS using 4 Tesla diffusion tensor MRI. Amyotroph Lateral Scler 12:52–58
Eslinger PJ, Moore P, Antani S, Anderson C, Grossman M (2012) Apathy in frontotemporal dementia: behavioral and neuroimaging correlates. Behav Neurol 25:127–136
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Barbagallo, G., Nicoletti, G., Cherubini, A. et al. Diffusion tensor MRI changes in gray structures of the frontal-subcortical circuits in amyotrophic lateral sclerosis. Neurol Sci 35, 911–918 (2014). https://doi.org/10.1007/s10072-013-1626-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10072-013-1626-z