
Abstract
In recent years, the use of operant conditioning procedures has shown that species as diverse as chimpanzees, honeybees, and mosquitofish can be trained to discriminate between sets containing different numbers of objects. However, to succeed in this task, subjects can use two different strategies: either select the array containing a specific number of items (an absolute numerosity rule), or select the set containing the larger (or smaller) quantity of items (a relative numerosity rule). In the latter case, subjects need not only be able to judge whether two numerosities are equal or different but also be able to order numerosities. Here, in two experiments, we address whether fish can perform both kinds of judgment by training them with specific numerosities and testing their generalization to new numerosity contrasts. In Experiment 1, subjects were initially trained to select between visual arrays of 6 and 12 shapes, and were then tested with a contrast pairing the previously trained numerosity (either 6 or 12) with a novel numerosity (respectively, 3 or 24). Spontaneously, subjects selected the novel numerosity, in accordance with a relative numerosity rule. The second experiment tested whether guppies can also learn to select one specific number against all others, if appropriately trained. Fish trained to select an array of 4 shapes against several alternatives (4 vs. 1, 4 vs. 2, 4 vs. 8, 4 vs. 10) learned to recognize the number 4 against all alternatives and proved able to generalize their discrimination to novel, more difficult contrasts (4 vs. 3 and 4 vs. 6 items). In summary, although guppies preferentially opt for relative comparisons, they can flexibly learn either relative or absolute decision criteria on numerosity stimuli, depending on the context.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Comparative research in the last decades demonstrated the existence of rudimentary numerical abilities in non-human animals. The capacity to use numerical information has been found not only in mammals (rhesus monkeys: Beran 2006; elephants: Perdue et al. 2012; horses: Miletto Petrazzini 2014; dogs: West and Young 2002), but also in birds (Pepperberg 2006; Rugani et al. 2008), fish (Buckingham et al. 2007; Agrillo et al. 2011, 2012b; Gómez-Laplaza and Gerlai 2013a, b), as well as in invertebrates (Gross et al. 2009; Nelson and Jackson 2012). One methodology most commonly adopted in this research topic consists of training subjects to discriminate between two visual stimuli differing in numerosity in order to obtain a food reward (e.g., Cantlon and Brannon 2007; Rugani et al. 2008; Gross et al. 2009; Agrillo et al. 2011; Vonk and Beran 2012). In these studies, the size of the items presented and their spatial configurations are varied in order to control for non-numerical perceptual cues such as density or overall area. Provided that no other information can be used as a discriminative cue, subjects will reach the learning criterion in these tasks only if they are sensitive to numerosity.
When subjects learn to discriminate between two values of numerosity, they may be applying two kinds of numerical strategies. First, they might have learned a rule about a specific value of numerosity: “select four objects”—hereafter called the “absolute numerosity” rule. Alternatively, they might have learned to select the larger (or smaller) group regardless of the absolute numerosity of the arrays, a strategy hereafter referred to as the “relative numerosity” rule. In both cases, the behavioral output is the same, thus making it impossible to decide which rule subjects applied in a traditional operant conditioning procedure.
There is evidence in the comparative literature that non-human animals use a relative rule in some cognitive domains. For instance, in a study on relative size transposition, one parrot trained to respond to the smaller or the larger of the two stimuli responded on a relative basis in test trials when presented with novel-sized objects, larger or smaller than those used during training (Pepperberg and Brezinsky 1991). Similar abilities to use a relative rule in a size discrimination task have been reported in horses (Hanggi 2003). In numerical cognition studies, the picture is less clear. For instance, in a study by Brannon and Terrace (1998), rhesus monkeys were initially trained to discriminate between groups of two-dimensional figures presented on a monitor in the range 1–4; subsequently, the subjects proved able to generalize the numerical rule also to novel numerosities (range 5–9), thus excluding the possibility that subjects were using an absolute numerosity rule. However, during the training phase, the subjects were required to learn a relative numerosity rule, as different combinations of numerical contrasts between 1 and 4 were presented, making impossible to focus on a specific numerosity as positive stimulus. In contrast, Davis (1984) reported that a raccoon successfully learned to select a set containing three items from alternative arrays containing one, two, four, and five items after extensive training. In this task, it is likely that the raccoon was reinforced to use an absolute numerosity rule as it was required to always select a specific number of objects against different alternatives containing a smaller or larger number of objects. These examples raise the possibility that the training procedure employed may influence the type of rule applied by the subjects. However, to our knowledge, no study has addressed whether a relative or an absolute responding rule is spontaneously preferred in an animal species, nor whether animals from the same species can be successfully trained to use both strategies.
In the present study, we tackled these issues in fish. In the first experiment, guppies (Poecilia reticulata) were trained to discriminate between 6 and 12 objects in order to obtain a food reward. After reaching the learning criterion, they were subjected to probe trials in which a novel numerical contrast was presented (e.g., 12 vs. 24 if they had previously been trained to choose 12 over 6 objects). If guppies spontaneously apply an absolute numerosity rule, they should select the numerosity reinforced in the previous training phase (12), even though 12 represents the smaller group in comparison with 12 versus 24. On the contrary, if they use a relative numerosity rule, they should select the larger group composed of 24 items, even though the numerosity previously reinforced is also available (12).
As guppies proved to use a relative numerosity rule in Experiment 1, we set up Experiment 2 in order to assess whether they can learn to select a specific number of objects against all alternatives (absolute numerosity rule). For this purpose, guppies were trained to choose a group containing four objects against both a smaller group of objects (1 or 2) and a larger group of objects (8, 10). They were then tested in probe trials for their ability to generalize to novel contrasts (4 vs. 3 and 4 vs. 6 items).
Experiment 1
In the present experiment, we tested whether guppies, trained to discriminate between two numerosities (e.g., 6 vs. 12), generalize to new numerosity contrasts by using an absolute numerosity rule (e.g., select number 12) or by using a relative numerosity rule (select the larger group).
Subjects
Thirty-two adult female guppies (Poecilia reticulata) were tested. The fish used in this experiment were tenth-generation descendants of wild caught fishes collected from the Tacarigua River in Trinidad. Subjects were maintained at the Department of General Psychology in 150 one-stock aquaria containing mixed-sex groups (15 individuals with approximately a 1:1 sex ratio). Aquaria were provided with natural gravel, an air filter, and live plants. Both stock aquaria and experimental tanks were maintained at a constant temperature of 25 ± 1 °C and a 14- to 10-h light/dark (L/D) photoperiod with an 18-W fluorescent light. Before the experiment, fish were fed twice daily to satiation with commercial food flakes and live brine shrimps (Artemia salina).
Apparatus and stimuli
We used the apparatus previously adopted in two studies investigating numerical abilities in fish (Agrillo et al. 2012b, 2014). The experimental apparatus was composed of a 50 × 19 × 32 cm tank (Fig. 1) filled with gravel and 24 cm of water. The short walls were covered with white plastic material, while the long ones were covered with green plastic material. Two mirrors (29 × 5 cm) were placed in the middle of the tank, 3 cm away from the long walls, in order to reduce the potential effects of social isolation (Miletto Petrazzini et al. 2012). In addition, an artificial leaf (9 × 8 cm) was placed in the middle to provide some shelter for the subjects. Two “choice areas” were defined by white rectangles (14 × 12 cm) covered by a green net in correspondence with the sides in which stimuli were presented.

Experimental apparatus used in Experiment 1. Guppies were housed in the experimental tank for the entire experiment. Two mirrors were placed in front of each other in the middle of the tank. Two choice areas were defined by white rectangles covered by a green net in correspondence with the short walls. At the beginning of each trial, stimuli (two groups of figures differing in numerosity: 6 vs. 12 in the training phase, 12 vs. 24 or 3 vs. 6 in the test phase) were hanged on the short walls (color figure online)
Stimuli were printed within a 6 × 6 cm square area and were presented at the bottom of a 6 × 32 transparent plexiglass panel that was vertically inserted into water against the short walls. Stimuli were thus displayed in the lower part of the wall, 10 cm above the floor. They consisted of groups of black geometric figures (circles, ovals, triangles, crosses, stars, squares, and rectangles) differing in size on a white background. In the training phase, we presented the 6 versus 12 numerical contrast. In the test phase, we presented either 12 versus 24 (for subjects trained toward the larger numerosity as positive, see procedure) or 3 versus 6 (for subjects trained toward the smaller numerosity). Stimuli selected for the experiment were extracted from a pool of 24 different pairs for each numerical contrast. The overall configuration (i.e., the spatial arrangement of the figures within the square) of the stimuli was changed across sets (i.e., pairs of stimuli) to avoid the possibility of fish using pattern recognition of the arrays instead of numerical information as the discriminative cue.
Numerosity covaries with other physical attributes, such as cumulative surface area, overall space encompassed by the stimuli, or density of the arrays. These non-numerical cues are commonly referred to as “continuous quantities” (Feigenson et al. 2002; Gebuis and Reynvoet 2012; Gómez-Laplaza and Gerlai 2012, 2013a, b). To control for continuous quantities, we used a procedure previously adopted in other studies on guppies (Agrillo et al. 2014; Bisazza et al. 2014). In particular, for one-third of the stimuli, the two numerosities were 100 % equated for cumulative surface area. However, a by-product of equating the cumulative surface area was that smaller-than-average objects would be more frequent in the larger groups, and guppies might use this information instead of number. To reduce this possibility, cumulative surface area was controlled to 85 % in another third of the stimuli during the training phase, and in the remaining one-third of the stimuli, it was controlled to 70 %. As a consequence, in the 70 % condition, the biggest object within each stimulus pair was shown in the larger set; in the 85 % condition, the biggest object was shown in the larger set in half of the trials and in the smaller set in the other half of trials. In probe trials, cumulative surface area was always matched to 100 %. In this way, should guppies discriminate between the two arrays in probe trials, neither cumulative item area, matched to 100 %, nor the individual item size, an unreliable cue in the training phase, could have played a key role. Furthermore, as density is negatively correlated to the overall surface occupied by the array, half of the set was controlled for overall surface, whereas the second half was controlled for density.
Sixteen identical experimental tanks, placed close to each other on the same table, were used. A video camera was suspended about 1 m above the experimental tanks and used to record the position of the guppies during the tests.
Procedure
The experiment was divided into three different steps: familiarization, training, and test. Familiarization was set up to familiarize the fish with the experimental apparatus. Subsequently, subjects were singly trained to discriminate between 6 and 12. The fish that reached the learning criterion were subsequently tested in extinction with a novel numerical contrast (either 12 vs. 24 or 3 vs. 6). Should the fish have used an absolute numerical rule, they were expected to select the numerosity reinforced during the previous training phase.
Familiarization
During the 6 days preceding the beginning of training, guppies were gradually familiarized with the experimental apparatus. On the first 2 days of the familiarization phase, four subjects were inserted in the apparatus for a total of 4 h and then returned to their aquarium; in the following 2 days, two subjects were inserted in the apparatus for 4 h; then, the fish stayed singly in the apparatus for the following 2 days. In this latter period, guppies were fed twice a day with Artemia salina nauplii that were inserted in the morning and in the afternoon near the two short walls.
Training
All subjects were presented with the same numerical contrast: 6 versus 12. Half of the fish received food near the larger numerosity, while the other half received food near the smaller numerosity, in order to reinforce an association between the positive numerosity (larger or smaller) with the reward (food). More specifically, on days 1–3, fish received four trials per day (three consecutive days, for a total of 12 trials). Each trial consisted of inserting two stimuli panels on the short walls soon followed by the release of a food reward (nauplii of Artemia salina) near the reinforced stimulus. Food consisted of live brine shrimps released with a Pasteur pipette; an identical syringe released pure water near the non-reinforced numerosity. Subjects were left free to feed for 7 min after which stimuli were removed from the tank. The intertrial interval lasted 3 h. We counterbalanced the position of the stimuli (left–right) over trials. Subjects were moved from one tank to another at the end of each day in order to avoid the possibility of using the local/spatial cues of the tank.
From day 4, subjects continued to receive four trials per day with the same stimuli, but they obtained a reward only if they chose the correct stimulus. In these trials, the two stimuli were introduced, while the fish was outside the two choice areas. We consider the first choice area entered by the subject as an indicator of its choice. In particular, the subject was considered as selecting a stimulus when its head was inside the choice area associated with that stimulus. If the choice was correct, we released the food near that stimulus; if the choice was incorrect, both stimuli were removed and the trial ended. If guppies did not select a choice area within 2 min, the trial was considered null. This procedure continued until the subject reached a learning criterion of 10 correct trials in 12 consecutive trials (83 % accuracy, corresponding to a statistically significant choice with the binomial test). If subjects did not reach the learning criterion within 20 trials, the training phase was interrupted and the fish did not start the test phase.
To date, studies on discrimination learning in fish have used both the proportion of correct choices (Gierszewski et al. 2013; Bisazza et al. 2014) and the association time near the positive stimulus (Agrillo et al. 2012b, c) as measures of learning. While it has been shown, in rats, that response choice and response time are correlated and both are reliable indicators of discrimination learning (Olton and Samuelson 1974), no study has currently investigated whether these variables could be indifferently used as an index of learning in fish. A recent study showed that numerical acuity of goldbelly topminnows is partially different when response choice is used instead of association time (Agrillo and Dadda 2007). However, the study was not specifically designed to investigate this issue, as different apparatuses and procedures were also used in the two tests. To shed light on this topic, at the end of the training phase, all fish (including those not admitted to the test phase) were subjected to two probe trials (intermingled with a reinforced trial) in which stimuli were inserted for 4 min in the tank without any reinforcement, and we measured the proportion of time spent near the positive numerosity. The proportion of correct choices was then correlated with the proportion of time.
Test
In test phase, three probe trials were presented each day for four consecutive days. Fish trained to select the larger numerosity (12) as positive were presented with 12 and 24 objects; fish trained to select the smaller numerosity (6) as positive were presented with 3 and 6 objects. The intertrial interval lasted 3 h. Two reinforced trials presenting the numerical contrast of the training (6 vs. 12) were alternated in the probe trials. As dependent variable, we considered the proportion of time spent in the “choice areas” (accuracy) during probe trials.
Proportions were arcsine (square-root)-transformed (Sokal and Rohlf 1995). Mean ± SD are provided. Statistical tests were carried out using SPSS 20.0.
Results
Training (6 vs. 12)
Sixteen subjects out of 32 (50 %) reached the learning criterion within 20 trials. Eight of these subjects were trained toward the larger numerosity as positive and eight toward the smaller numerosity as positive. There was no difference in the proportion of correct choices between fish trained toward the larger numerosity as positive (0.694 ± 0.121) and those trained with the smaller numerosity as positive (0.668 ± 0.151, independent t test, t(30) = 0.506, P = 0.617). The same result was obtained when we analyzed the performance of the 16 subjects who reached the learning criterion (larger numerosity as positive: 0.800 ± 0.048, smaller numerosity as positive: 0.798 ± 0.048, t(14) = 0.103, P = 0.919).
One-sample t tests conducted on all 32 fish evidenced a significant discrimination of the two numerosities (6 vs. 12), both considering the proportion of correct choices during training [0.68 ± 0.14, t(31) = 7.419, P < 0.001] and the proportion of time spent near the correct stimulus in the probe trials [0.67 ± 0.25, t(31) = 3.803, P = 0.001]. A significant positive correlation between these two measures of learning was found (Pearson’s test, r = 0.566, P = 0.001, Fig. 2).

Results of Experiment 1: training. A positive correlation between the two dependent variables was found (proportion of correct choices: X-axis, proportion of time spent near the reinforced numerosity: Y-axis), meaning that the proportion of time spent near the reinforced numerosity is a reliable predictor of the proportion of correct choices
Test (12 vs. 24 or 3 vs. 6)
In the probe trials of the test phase, no difference was found between fish trained toward the larger numerosity as positive (0.67 ± 0.12) and those trained toward the smaller numerosity as positive (0.74 ± 0.13) in the proportion of time spent near the previously non-reinforced numerosity [independent t test, t(14) = 1.027, P = 0.322]. We accordingly pooled together the two groups in subsequent analyses.
On the whole, guppies exhibited a significant preference for the non-reinforced numerosity [0.71 ± 0.13, t(15) = 5.740, P < 0.001, Fig. 3]. No difference was found when we contrasted the overall accuracy in the first half of the test phase (trials 1–6: 0.68 ± 0.16) with the overall accuracy in the second half [trials 7–12: 0.74 ± 0.15, paired t test, t(15) = 1.33, P = 0.203], meaning that the performance observed is not influenced by any form of learning during the test phase.

Results of Experiment 1: training and test. Y-axis refers to the proportion of time spent near the reinforced numerosity in the training phase (16 subjects admitted to the test phase), and the proportion of time spent near the novel numerosity (smaller or larger than the reinforced numerosity) in the test phase. While subjects spent more time near the reinforced numerosity during the training, they switched their preference in the test phase and spent more time near the stimulus that presented a larger/smaller number of objects. Bars represent the standard errors. Asterisks denote a significant departure from chance level
Discussion
The results of this experiment show that fish that had learned to discriminate between two numerosities will later generalize to a relative numerosity rule, selecting a novel numerosity over the previously reinforced numerosity. Relative numerosity rules are thus preferred over absolute numerosity rules in fish, despite the fact that applying a relative rule might be cognitively more challenging because it requires the ordering of numerosities presented. In other cognitive tasks, learning to respond on the basis of a relative rather than an absolute rule has been considered more complex because it requires the subject to compare the stimuli and then derive a general rule instead of forming a single association (Thomas 1980; Pepperberg and Brezinsky 1991). Similarly, in the case of numbers, same–different judgments (absolute rule) are logically simpler than more–less judgments (relative rule). Being able to assess whether a numerosity is larger/smaller than another implies the capacity to represent the absolute numerosity of both sets, in order to compare these two numerosities. Alternatively, it is possible to hypothesize that the peak shift phenomenon described in several species in other cognitive tasks may explain our data (Dougherty and Lewis 1991; Cheng et al. 1997). In peak shifts, subjects are initially trained to respond to a positive stimulus (S+) and withhold responding to a negative stimulus (S−) that is similar, although not identical, to S+ [see Purtle (1973) for a review]. In probe trials, subjects are tested in the absence of reward on a continuum of stimuli including S+ and S−: maximal response is not commonly observed with S+ but to a stimulus similar to S+; the novel stimulus selected is also more distinct from S−. This phenomenon seems to occur as a consequence of learning different outcomes for two partially similar stimuli and is supposed to involve basic associative processes (Perry et al. 2013).
In our experiment, guppies were simultaneously trained to reach a positive stimulus (e.g., S+: 12) and avoid a negative one (S−: 6); in test phase, subjects did not select S+ but a novel stimulus (24) more distinct from S−, a pattern of response that resembles peak shift phenomenon. However, it is worth noting that in test phase, we presented only two alternative stimuli instead of a continuum of stimuli from S+ to S−, a methodological difference that prevents us a fine comparison with the existing literature on this effect. In addition, the size of peak shift was found to be inversely related to the magnitude of the difference between S+ and S− (Hanson 1959). In our study, the ratio between S+ and S− was equal to 2:1, a large numerical ratio easily discriminated by guppies (Agrillo et al. 2014) that should have led to a reduced peak shift. In contrast, guppies showed a strong generalization to the novel numerosity, thus suggesting that the pattern of data here reported could be only partially ascribed to the simple associative models underlying peak shifts.
Because we measured the fish discrimination using two different dependent variables within individuals, Experiment 1 also provided useful information on the relation between the dependent variables used in training studies. Usually, traditional operant conditioning studies use the proportion of correct choices in many consecutive probe trials as a measure of learning. Here, we found that this variable correlates positively with the time spent near the reinforced numerosity, meaning that the association time is a reliable predictor of the proportion of correct choices commonly observed with a traditional procedure [reviewed in Agrillo and Bisazza (2014)].
One may argue that only half of the subjects met the learning criterion in the training phase, thus making it difficult to understand whether our results can be widely generalized to guppies’ populations. The proportion of fish starting the test phase in our study is similar to the proportion reported in previous studies that used this method of operant conditioning (Agrillo et al. 2012b, c; Bisazza et al. 2014). With respect to this topic, it is important to notice that fish that do not meet the learning criterion after a few trials do not usually improve their performance even after extensive training (e.g., Agrillo et al. 2012b); hence, these individuals apply neither an absolute nor a relative numerosity rule.
In sum, Experiment 1 showed that trained guppies, when given the possibility to use two alternative solutions, an absolute rule (select a given number of objects) or a relative rule (select the largest or smallest number of objects), spontaneously favor the latter strategy. However, this result does not necessarily imply that they are not equipped with the cognitive ability to learn an absolute rule if tested in different conditions. We addressed this issue in Experiment 2.
Experiment 2
In this experiment, we trained guppies to select a fixed number of objects (4) against both larger and smaller quantities, and studied whether fish were able to generalize the learned rule when presented with novel numerical contrasts. For this experiment, we used a novel, recently developed setup for operant conditioning in guppies (Bisazza et al. 2014; Lucon-Xiccato and Bisazza 2014).
Subjects
Ten adult guppies, one male and nine females of a domestic strain, participated in this experiment. Five subjects (one male and four females) have been previously trained to dislodge discs in a shape discrimination task, but they had no previous experience in numerical training (subgroup 1). The other five fish were naïve (subgroup 2). The group was initially composed of fourteen fish, but four subjects (two males and one female from subgroup 1 and one female from subgroup 2) were excluded from the experiment because they ceased to participate in the early stages of the experiment.
Apparatus and stimuli
The experimental apparatus was similar to that used in two recent studies that investigated discrimination learning in guppies (Lucon-Xiccato and Bisazza 2014; Bisazza et al. 2014). It consisted of a 60 × 40 cm tank, filled with gravel and 30 cm of water maintained at a temperature of 25 ± 2 °C (Fig. 4a). The apparatus was divided into two compartments: the back compartment, with live plants, and the front “experimental compartment.” A start box was inserted between the compartments and was provided with a transparent guillotine door. Each compartment was lit by a 15-W fluorescent light. The apparatus was provided with two air filters. A green plastic panel (20 × 15 cm) with 83 holes (Ø 1 cm, depth 0.5 cm) was placed on the gravel substratum of the experimental compartment (30 × 40 cm), close to the front wall. The bottom of the panel was covered with a green net to allow the smell of the food reward to pervade the whole compartment and so to reduce the possibility for the subject to identify the correct choice using olfactory cues. Four social companions were kept in the apparatus to avoid social isolation and were temporarily removed at the beginning of each session.

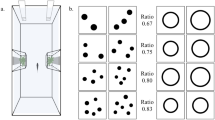
Experimental apparatus used in Experiment 2 (a). The back compartment was provided with live plants, and a green plastic plate was placed on the gravel in the experimental compartment. The start box was inserted between the compartments. Stimuli were placed on the green plate, and a pair of yellow discs was used to completely cover the holes in front of each stimulus during both training and control tests (b) (color figure online)
Stimuli consisted of laminated panels placed orthogonally to the green panel (Fig. 4b). Stimuli were groups of black dots on a white background (5 × 5 cm). During training, we presented four different numerical contrasts: 4 versus 1, 4 versus 2, 4 versus 8 and 4 versus 10. In test 1, fish were tested with the same comparisons but with novel spatial configurations. In test 2, fish were presented with two novel numerosities: 4 versus 6 and 4 versus 3. The size of the dots and their spatial distribution varied across pairs to avoid pattern recognition. In addition, stimuli were controlled for continuous quantities as described in Experiment 1. More specifically, in one-third of the stimuli, the cumulative surface area was matched to 100 %; in another third, it was matched to 75 %; and in the remaining third, the cumulative surface area was not controlled (i.e., the ratio between the cumulative surface area within each pair was congruent with the numerical ratio). In probe trials of test 2, cumulative surface area was always matched to 100 %. Stimuli were extracted from a pool of 18 different pairs for each numerical contrast (six pairs for each control condition).
Two yellow plastic discs (Ø 1.2 cm, height 0.2 cm) were used during the experiment to cover the holes of the panel.
Procedure
We used a modification of the procedure recently employed to study learning abilities in guppies (Bisazza et al. 2014; Lucon-Xiccato and Bisazza 2014).
The procedure consisted of four phases: familiarization, pre-training, training, and control tests.
Familiarization
Two subjects were introduced into the apparatus 4 days before the beginning of training in order to familiarize them with the experimental apparatus. On the first 2 days, fish received food twice daily in the presence of a pair of stimuli (the same numerical contrasts used later in the training phase) randomly presented in order to habituate them to their presence. Commercial food flakes were released through a Pasteur pipette in correspondence with the stimulus showing four items. From the third day, fish were familiarized with the experimental procedure. Four times a day for two consecutive days, fish were gently encouraged to enter the start box using a transparent plastic panel. Subsequently, the transparent guillotine was closed from a remote location, and a green plastic barrier was placed in front of it to prevent the fish from seeing the experimental compartment. Dry food was delivered in five holes close to the stimulus showing four items, and two yellow discs were placed nearby to familiarize fish with their presence. Subsequently, fish were allowed to enter the experimental compartment and eat the food. Each numerical contrast was presented twice. On the evening of day 4, fish were individually housed in an experimental apparatus. The training phase began the next day.
Pre-training
In this phase, fish were trained to dislodge the discs. A total of 18 trials were administered in two consecutive days. At the beginning of each trial, the subject was gently ushered into the start box, the guillotine was closed, and the green panel was placed in front of it. A pair of stimuli was placed on the green plate, and a yellow disc was used to partially cover the hole in front of each stimulus. On day 1, fish received a total of eight trials, subdivided into two sessions of four each (one in the morning and one in the afternoon). The intersession interval lasted 5 h. In trials 1–3, the discs covered 25 % of the holes; in trials 4–6, they covered 50 %; in trials 7–8, they covered 75 %. On day 2, fish received a total of 10 trials, subdivided into two sessions of five each. In trials 1–2 and 6–7, the discs covered 75 % of the holes; in trials 3–5 and 8–10, the discs completely covered the holes. The same four numerical contrasts of the successive training phase (4 vs. 1, 4 vs. 2, 4 vs. 8, 4 vs. 10) were presented in random order, and a food reward was placed each time under the disc in front of the stimulus with four items. These trials were not included in the statistical analyses.
Training
In this phase, the subjects received 12 trials per day, subdivided into two sessions of six each.
Each trial started with the subjects in the start box and the green barrier in place to block the view of the experimental compartment. A pair of stimuli was placed on the plate, and yellow discs covered the corresponding holes entirely. Subsequently, the barrier was removed, and after 30 s, the guillotine was opened to allow the subject to enter the experimental compartment. The choice was defined as the first disc dislodged by the fish. In order to keep the subject motivated, we adopted a correction procedure, and when the fish opened the wrong disc, it was allowed to open the correct one and eat the food. If the subject did not dislodge any disc within 5 min, the trial was considered invalid and repeated later. The distance from the corridor and the position of the stimuli on the panel was determined with a pseudorandom rule. To avoid the side bias, the positive stimulus was never presented more than twice in a row on the same side, and the left–right position of the stimuli was counterbalanced over trials. Each numerical contrast was presented three times a day, once for each control condition.
The learning criterion was set at 70 % correct trials over two consecutive days (corresponding to a statistically significant preference with the Chi-square test calculated on the 2 days).
Once the criterion was reached, the fish entered the test phase.
Test
The test phase consisted of two tests. In test 1, four probe trials intermingled with 12 standard trials were presented for four consecutive days (for a total of 16 daily trials subdivided into two sessions of eight trials each). In probe trials, fish were presented with the same numerical contrasts (4 vs. 1, 4 vs. 2, 4 vs. 8, 4 vs. 10) used during training but with completely novel spatial configurations (e.g., dots were aligned, a pattern never presented during training). There were four presentations for each numerical comparison. Subjects were non-differentially reinforced for their selections on these trials with a probability of 0.50 for reward and a probability of 0.50 for no reward. Standard trials were normally reinforced.
In test 2, four probe trials intermingled with 12 standard trials were presented for four consecutive days (for a total of 16 daily trials subdivided into two sessions of eight trials each). In these probe trials, fish were presented with two novel numerical contrasts with closer numerical ratio: 4 versus 6 (0.67 ratio) and 4 versus 3 (0.75 ratio), eight presentations for each numerical comparison. Half of the probe trials was randomly rewarded, and the other half was not as in test 1; standard trials were normally reinforced. Two fish (both females from subgroup 1) ceased to participate in this test after 2 days and one fish (a female from subgroup 2) after 3 days.
As during training, the distance from the corridor and the left–right position of the stimuli was pseudorandomly determined. In both tests, we considered the proportion of choices for the number 4 during probe trials as the dependent variable. Proportions were arcsine square-root-transformed (Sokal and Rohlf 1995). Mean ± SD are provided.
Results
Training
On average, all subjects reached the learning criterion in 81.6 ± 30.88 trials with no difference in learning rate between subgroups 1 and 2 [independent t test, t(8) = 0.981, P = 0.355]. None of the subjects exhibited a side bias (all Ps > 0.05).
Test 1
Fish showed a statistically significant preference for the trained stimulus even when they were presented with novel spatial configurations [0.687 ± 0.065, t(9) = 8.441, P < 0.001, Fig. 5]. We found no statistical difference in the proportion of correct choices between subgroup 1 (0.725 ± 0.071) and subgroup 2 [0.650 ± 0.034; t(8) = 2.119, P = 0.067].

Results of Experiment 2. In test 1, fish significantly selected the number 4 in the presence of completely novel pattern configurations. In test 2, fish generalized the rule to novel numerosities and successfully selected the number 4 with novel numerical contrasts: 4 versus 3 and 4 versus 6. Bars represent the standard errors. Asterisks denote a significant departure from chance level
Test 2
Fish showed a statistically significant preference for the trained stimulus even when they were presented with novel, more difficult numerical ratios [0.629 ± 0.073; t(9) = 5.475, P < 0.001, Fig. 5]. We found no statistical difference in the proportion of correct choices between subgroup 1 (0.650 ± 0.071) and subgroup 2 [0.608 ± 0.077; t(8) = 0.890, P = 0.399]. When the numerical contrasts were examined separately, a significant preference for the number 4 was observed both in 4 versus 3 [0.629 ± 0.103, t(9) = 3.931, P = 0.003] and in 4 versus 6 [0.629 ± 0.132, t(9) = 3.097, P = 0.013] with no significant difference between the two numerical contrasts [paired t test, t(9) = 0.015, P = 0.988].
When we compared the tests, fish showed to be more accurate in test 1 than in test 2 [paired t test, t(9) = 3.109, P = 0.013].
Discussion
The second experiment was designed to test whether guppies can learn to use an absolute numerosity rule if appropriately trained. Fish learned this task in a relatively small number of trials in comparison with other studies where fish underwent more trials to learn shape discriminations (approximately 100 trials: Sovrano and Bisazza 2008; Siebeck et al. 2009) or numerical discriminations (approximately 150/200; Agrillo et al. 2009, 2010).
However, since guppies and their relatives possess sophisticated learning skills and can discriminate complex patterns such as the individual features of another guppy (Bisazza 2010), they could have hypothetically solved the task by learning all 72 different pairs of stimuli or at least some of them. To rule out this possibility, in the first control test, we presented the same numerical comparisons but with novel, very different configurations and found that guppies retain their ability to choose the stimulus containing four objects. Thus, we can exclude that pattern recognition took place over training trials. A second possibility is that guppies have simultaneously learned to avoid quantities that approximate 1 or 2 units and quantities that approximate 8–10 units. To test for this possibility, in test 2, we proposed probe trials in extinction with novel numerical comparisons, 4 versus 3 and 4 versus 6. Even in this case, fish selected the number 4 more often than chance. One may argue that if fish learned to avoid quantities both smaller and larger than 4, results in test 2 might be due to fish avoidance to the novel numerosities because 6 is closer to 8–10 and 3 is closer to 1–2 than 4. Though we cannot exclude this hypothesis, it is worth noting that the question whether they learned to avoid 1, 2, 3, 6, 8, 10 or whether they learned to select 4 is irrelevant to the absolute–relative rule issue because both strategies rely on an absolute rule and not on a relative rule.
The performance in test 2 was significantly lower than with the easier numerical contrasts of test 1, but it must be pointed out that, in terms of ratio, these contrasts are close to or even beyond the limits reported for most mammals and birds (Agrillo and Bisazza 2014). Actually, the numerical acuity seen in test 2 appears to be higher than previously reported for guppies and fish in general. Previously, guppies were found to fail in discriminating 4 from 6 conspecifics (Agrillo et al. 2012a) and did not learn to discriminate 4 from 3 objects with two-dimensional stimuli, although they could discriminate these quantities when stimuli were in motion (Agrillo et al. 2014). In these studies, guppies were very likely to use an approximate relative numerosity rule to solve their quantity tasks. In Experiment 2, this strategy was not available; therefore, our results suggest the possibility that learning an absolute numerosity rule had a refining effect on guppies’ representations of numerosity, perhaps by providing an anchor in memory.
It is worth noting, however, that in this experiment, guppies were trained with more trials compared to previous studies using operant conditioning procedures in fish (Agrillo et al. 2012b, 2014) and had a longer training experience when faced with the more difficult discriminations. As a consequence, the possibility exists that the extensive training increased their numerical acuity, as recently reported in the same species (Bisazza et al. 2014).
General discussion
Discrimination learning between two quantities of objects is probably the most frequently used technique in the investigation of numerical abilities of non-human species. Although this technique has been widely employed in non-human primates, birds, fish, and insects, in many studies, the subjects could have solved the task either by applying an absolute numerosity rule (select that exact numerosity) or by applying a relative numerosity rule (select the larger/smaller array). It is thus not clear whether animals can apply both or only one of these rules—and, in this case, which rule. In the present study, we investigated this issue in guppies in two experiments where subjects were required to make numerosity judgments between groups of two-dimensional figures.
In Experiment 1, we trained guppies to perform a numerical discrimination, 6 versus 12 elements until criterion. We then tested them in extinction in a critical numerical contrast, where the previously trained numerosity was pitted against a new numerosity, which was either twice as large (24, for the fish previously trained to select 12 over 6), or twice as small (3, for the fish previously trained to select 6 over 12). Overall, fish spontaneously responded according to a relative numerosity rule: they selected the new numerosity over the numerosity that had been previously reinforced.
However, the fact that guppies do not exhibit any preference for using an absolute rule does not necessarily imply that they are not equipped with the cognitive ability to learn an absolute rule. To test this hypothesis, in Experiment 2, guppies were subjected to a task that could not be solved using a relative numerical rule. They were first trained to select a group containing four objects both when paired with smaller (4 vs. 1 or 4 vs. 2 objects) or with larger quantities (4 vs. 8 or 4 vs. 10 objects) and then tested with numerosity 4 paired against familiar and novel numerosity values. In this experiment, fish succeeded in applying an absolute numerosity rule, thus proving that both types of rules are within their repertoire, but which is chosen depends on the training context.
To date, the literature about the use of absolute and relative numerical representation is restricted to a few mammals tested after extensive training (e.g., Davis 1984; Brannon and Terrace 1998). This study represents the first evidence showing that also a species distantly related to humans, such as guppies, can use both strategies when required to make relative numerosity judgments. In other words, while previous studies have shown that numerical acuity of non-human animals can largely vary according to the experimental procedure adopted [e.g., training procedures vs. spontaneous choice tasks, food as stimuli vs. two-dimensional figures as stimuli, reviewed in Agrillo and Bisazza (2014)], the present work extends these methodological findings by showing that the type of training procedure used influences not only the numerical acuity but also the cognitive strategy adopted by subjects involved in numerosity judgments.
Although fish can use both relative and absolute numerosity rules, they spontaneously favor the relative rule. Applying a relative rule is supposed to require a greater level of abstraction than an absolute rule (Thomas 1980; Pepperberg and Brezinsky 1991) because subjects need to abstract away the absolute values of numerosities and instead decide based on a relation between the two numerosities. However, relative rules are also more adaptive. In nature, animals have to repeatedly face relative numerosity judgments, such as selecting the largest quantity of food items to optimize food intake (Royama 1970; Bodmer 1990) or selecting the largest group of social companions in order to dilute the chance to be captured by predators (Foster and Treherne 1981; Cresswell 1994). In these circumstances, it is not the absolute number that counts, as normally there is not an optimal quantity of food items or an optimal shoal size to select. Instead, in all these situations, selecting the most advantageous option requires applying a relative numerosity rule, regardless of the exact numerosities involved. Because these problems are faced by most species, we suggest that the relative numerosity rule must be the most common strategy in many species.
In conclusion, our study provides evidence that trained guppies flexibly learn to apply different numerical criteria depending on the context, although they spontaneously use a relative numerical rule, probably because of its higher adaptive value. Future studies are now required to assess whether these cognitive abilities are unique in guppies or instead are shared by other fish and, more broadly, other non-human animals.
References
Agrillo C, Bisazza A (2014) Spontaneous versus trained numerical abilities. A comparison between the two main tools to study numerical competence in non-human animals. J Neurosci Methods 234:82–91
Agrillo C, Dadda M (2007) Discrimination of the larger shoal in the poeciliid fish Girardinus falcatus. Ethol Ecol Evol 19(2):45–157
Agrillo C, Dadda M, Serena G, Bisazza A (2009) Use of number by fish. PLoS ONE 4(3):e4786
Agrillo C, Piffer L, Bisazza A (2010) Large number discrimination by fish. PLoS ONE 5(12):e15232
Agrillo C, Piffer L, Bisazza A (2011) Number versus continuous quantity in numerosity judgments by fish. Cognition 119:281–287
Agrillo C, Piffer L, Bisazza A, Butterworth B (2012a) Evidence for two numerical systems that are similar in humans and guppies. PLoS ONE 7(2):e31923
Agrillo C, Miletto Petrazzini ME, Piffer L, Dadda M, Bisazza A (2012b) A new training procedure for studying discrimination learning in fishes. Behav Brain Res 230:343–348
Agrillo C, Miletto Petrazzini ME, Tagliapietra C, Bisazza A (2012c) Inter-specific differences in numerical abilities among teleost fish. Front Psychol 3:483
Agrillo C, Miletto Petrazzini ME, Bisazza A (2014) Numerical acuity of fish is improved in the presence of moving targets, but only in the subitizing range. Anim Cogn 17:307–316
Beran MJ (2006) Quantity perception by adult humans (Homo sapiens), chimpanzees (Pan troglodytes), and rhesus macaques (Macaca mulatta) as a function of stimulus organization. Int J Comp Psychol 19:386–397
Bisazza A (2010) Cognition. In: Evans J, Pilastro A, Schlupp I (eds) Ecology and evolution of poeciliid fishes. Chicago University Press, Chicago, pp 165–173
Bisazza A, Agrillo C, Lucon-Xiccato T (2014) Extensive training extends numerical abilities of guppies. Anim Cogn 17(6):1413–1419
Bodmer RE (1990) Fruit patch size and frugivory in the lowland tapir (Tapirus terrestris). J Zool 222(1):121–128
Brannon EM, Terrace HS (1998) Ordering of the numerosities 1 to 9 by monkeys. Science 282:746–749
Buckingham JN, Wong BBM, Rosenthal GG (2007) Shoaling decision in female swordtails: how do fish gauge group size? Behaviour 144:1333–1346
Cantlon JF, Brannon EM (2007) How much does number matter to a monkey (Macaca mulatta)? J Exp Psychol Anim Behav Proc 33:32–41
Cheng K, Spetch ML, Johnston M (1997) Spatial peak shift and generalization in pigeons. J Exp Psychol Anim Behav Proc 23(4):469–481
Cresswell W (1994) Flocking is an effective anti-predation strategy in redshanks, Tringa totanus. Anim Behav 47(2):433–442
Davis H (1984) Discrimination of the number three by a raccoon (Procyon lotor). Anim Learn Behav 12:409–413
Dougherty DM, Lewis P (1991) Stimulus generalization, discrimination learning, and peak shift in horses. J Exp Anal Behav 56(1):97–104
Feigenson L, Carey S, Spelke ES (2002) Infants discrimination of number vs. continuous extent. Cogn Psychol 44:33–66
Foster WA, Treherne JE (1981) Evidence for the dilution effect and the selfish herd from fish predation on a marine insect. Nature 293:466–467
Gebuis T, Reynvoet B (2012) The role of visual information in numerosity estimation. PLoS ONE 7(5):e37426
Gierszewski S, Bleckmann H, Schluessel V (2013) Cognitive abilities in Malawi cichlids (Pseudotropheus sp.): matching-to-sample and image/mirror-image discriminations. PLoS ONE 8(2):e57363
Gómez-Laplaza LM, Gerlai R (2012) Activity counts: the effect of swimming activity on quantity discrimination in fish. Front Psychol 3:484
Gómez-Laplaza LM, Gerlai R (2013a) Quantification abilities in angelfish (Pterophyllum scalare): the influence of continuous variables. Anim Cogn 16:373–383
Gómez-Laplaza LM, Gerlai R (2013b) The role of body surface area in quantity discrimination in angelfish (Pterophyllum scalare). PLoS ONE 8(12):e83880
Gross HJ, Pahl M, Si A, Zhu H, Tautz J et al (2009) Number-based visual generalisation in the honeybee. PLoS ONE 4(1):e4263
Hanggi EB (2003) Discrimination learning based on relative size concepts in horses (Equus caballus). Appl Anim Behav Sci 83:201–213
Hanson HM (1959) Effects of discrimination training on stimulus generalization. J Exp Psychol 58:321–334
Lucon-Xiccato T, Bisazza A (2014) Discrimination reversal learning reveals greater female behavioural flexibility in guppies. Biol Lett 10(6):20140206
Miletto Petrazzini ME (2014) Trained quantity abilities in horses (Equus caballus): a preliminary investigation. Behav Sci 4:213–225
Miletto Petrazzini ME, Agrillo C, Piffer L, Dadda M, Bisazza A (2012) Development and application of a new method to investigate cognition in newborn guppies. Behav Brain Res 233:443–449
Nelson XJ, Jackson RR (2012) The role of numerical competence in a specialized predatory strategy of an araneophagic spider. Anim Cogn 15:699–710
Olton DS, Samuelson R (1974) Decision making in the rat: response-choice and response-time measures of discrimination reversal learning. J Comp Physiol Psychol 87(6):1134–1147
Pepperberg IM (2006) Grey parrot numerical competence: a review. Anim Cogn 9:377–391
Pepperberg IM, Brezinsky MV (1991) Acquisition of a relative class concept by an African gray parrot (Psittacus erithacus): discriminations based on relative size. J Comp Psychol 105:286–294
Perdue BM, Talbot CF, Stone A, Beran MJ (2012) Putting the elephant back in the herd: elephant relative quantity judgments match those of other species. Anim Cogn 15:955–961
Perry CJ, Barron AB, Cheng K (2013) Invertebrate learning and cognition: relating phenomena to neural substrate. Wiley Interdiscip Rev Cogn Sci 4(5):561–582
Purtle RB (1973) Peak shift: a review. Psychol Bull 80:408–421
Royama T (1970) Factors governing the hunting behaviour and selection of food by the great tit (Parus major L.). J Anim Ecol 39(3):619–668
Rugani R, Regolin L, Vallortigara G (2008) Discrimination of small numerosities in young chicks. J Exp Psychol Anim Behav Proc 34:388–399
Siebeck UE, Litherland L, Wallis GM (2009) Shape learning and discrimination in reef fish. J Exp Biol 212(13):2113–2119
Sokal RR, Rohlf FJ (1995) Biometry. Freeman and Company, New York
Sovrano VA, Bisazza A (2008) Recognition of partly occluded objects by fish. Anim Cogn 11:161–166
Thomas RK (1980) Evolution of intelligence: an approach to its assessment. Brain Behav Evol 17:454–472
Vonk J, Beran MJ (2012) Bears “count” too: quantity estimation and comparison in black bears (Ursus americanus). Anim Behav 84:231–238
West R, Young RJ (2002) Do domestic dogs show any evidence of being able to count? Anim Cogn 5:183–186
Acknowledgments
The authors would like to thank Laura Piffer for her help in testing the subjects and two anonymous reviewers for their useful comments. The study complies with all laws of the country (Italy) in which it was performed and was supported by “FIRB grant 2013” (prot.: RBFR13KHFS) to C. Agrillo from Ministero dell’Istruzione, Università e Ricerca (MIUR, Italy).
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Miletto Petrazzini, M.E., Agrillo, C., Izard, V. et al. Relative versus absolute numerical representation in fish: Can guppies represent “fourness”?. Anim Cogn 18, 1007–1017 (2015). https://doi.org/10.1007/s10071-015-0868-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10071-015-0868-y