
Abstract
Humans and other animals use the global geometry of the surrounding environment in order to orient and determine which direction they are facing. Accordingly, the impact of environment geometry on spatial behavior is reflected in the paths of progression in the environment. When the perception of the global geometry is limited, such as in large or dark environments, the global geometry of the environment has to be constructed gradually as the accumulated geometry of locales. In the present study, we progressively altered the form of a dark square test arena by means of local alterations to its corners and walls, in order to differentiate the impact of the global arena geometry from that of the local arena geometry sectors on spatial behavior in rats. We found that as long as the local alterations did not distort the global square geometry of the dark environment, the rats’ behavior did not significantly change. In contrast, distortion of the square shape of the arena resulted in significant changes in the spatial distribution of the rats’ activity. Accordingly, we suggest that the perceived global geometry affects spatial behavior, overriding the impact of the local geometry.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Humans and other animals construct a spatial representation of their surrounding environment and use this representation to orient (Lynch 1960; Tolman 1948); that is, to determine a heading direction (Gallistel 1990), the direction they are facing in the environment (Cheng and Newcombe 2005) and to become organized in time and space (Gallistel 1989). This process rests, at least partially, on the perception of the direction and distance between the environment and self (Gauzzelli et al. 1999). Information in the environment that may assist the navigator includes, for example, cues provided by the distal panorama (Pecchia and Vallortigara 2010; Stürzl et al. 2008), cues in the layout of the environment (Collett 1996; Etienne et al. 2000; Golledge 1999), and the geometry of the surrounding environment (Cheng and Newcombe 2005; Vallortigara 2008). Navigators may also use internal cues generated by their self-movement (e.g. vestibular and kinesthetic cues) in order to update their position in reference to a specific location; for example, the starting point of travel (Alyan 1996; Mittelstaedt and Mittelstaedt 1980). Together, external and internal cues are utilized in navigating and constructing spatial representation (Etienne et al. 1999; Shettleworth 1998, 2005; Shettleworth and Sutton 2005; Sovrano et al. 2007). After a representation of the environment has been acquired, spatial behavior may then comprise path-planning, which is based on identifying origin, destination, directions, and distances (Golledge 1999). In the present study, we set out to determine which geometric properties of the environment may influence the spatial organization of behavior.
Environmental cues are classified into geometric cues, such as the shape of the test environment, and featural cues, such as the color or texture of sectors of the environment (Cheng 1986). It was previously postulated that in a dedicated geometric module, geometric cues are preferentially encoded over featural cues (Cheng 1986). However, recent studies have demonstrated that featural cues can sometimes hinder the learning of geometric cues (see Cheng 2008 for review). Whether its effect is primary or secondary, it is clear that environment geometry is involved in shaping spatial behavior. Indeed, previous studies have revealed that navigators may exploit the boundaries of the test environment in order to estimate distances (Avni et al. 2008; Hamilton et al. 2007, 2008; Hartley et al. 2004; Horev et al. 2007; Maurer and Derivaz 2000; Yaski et al. 2009) and that boundaries are a discrete component in spatial memory (Doeller et al. 2008). In rats, the impact of boundaries is notable in the establishment of the home base at the arena corner or at the location where they were introduced into the arena (Nemati and Whishaw 2007), and their spatial behavior is then organized in relation to that home base, which is the terminal for roundtrips in the arena (Eilam and Golani 1989). When objects are placed in the arena, rats will travel to the objects regardless of their location, but retain the location of the home base as a terminal for travel (Zadicario et al. 2007). Accordingly, it was concluded that spatial behavior is primarily anchored at the start corner and from there extended to the objects, regardless of their location (Zadicario et al. 2007). Similarly, in a square arena with a corner-shaped object placed against one of the arena walls, rats preserved the home base at the start corner while extensively traveling from it to the corner-shaped object (Yaski and Eilam 2008). In the present study, we altered the local geometry of the arena by gradually modifying its corners and walls, in order to ascertain what shapes spatial behavior: the global arena geometry, or the local geometry of the corners and walls.
The extent to which animals act in response to spatial changes in experimental settings is probably influenced by variables such as the size of the test environment (Chiandetti et al. 2007; Chiandetti and Vallortigara 2008; Sovrano et al. 2005, 2007), the extent of the change relative to the enclosure (Yaski and Eilam 2007), and past experience in the setting (Zadicario et al. 2007). Most studies on spatial behavior are based on training subjects to reach a goal while relying on specific cues, and then altering these cues (Maaswinkel and Whishaw 1999; Stackman and Herbert 2002). Indeed, while in laboratory experimental settings animals may be able to encompass visually the entire surrounding environment, in nature they may need to navigate over greater areas and at any given time encounter only a limited part of the environment. Therefore, we tested the rats in a dark environment, since even in the relatively limited scale of a laboratory setting, a dark environment does not enable the animals to visually capture the entire surrounding, requiring them, at least initially, to explore limited sectors of the arena. In the dark arena, we tracked freely moving rats and, as in past studies, we drew conclusions regarding their spatial behavior from their movements and routes (Tchernichovski et al. 1996, 1998; Wallace et al. 2002). Specifically, we tested the rats in a dark square arena while gradually altering the geometry of the arena corners and walls, in order to determine what affects spatial behavior: global or local geometric changes. We asked the following three questions: (i) considering that corners are attractors for rats, what is the impact of adding corners; (ii) what is the impact of altering the local geometry of corners on spatial behavior; and (iii) how is spatial behavior affected by changes in the global geometry of the environment, such as distorting its symmetry and its simple square geometry.
Materials and methods
Animals
Twenty adult male Wistar rats (3 months old; weight 300–400 g) were housed in groups of 3–4 individuals in metal cages (60 cm × 40 cm × 25 cm) with sawdust bedding, in a temperature-controlled room (21°C) under 12/12 dark/light cycle (lights on 19:00–07:00). Standard rat pellets and water were provided ad lib. Rats were individually marked by colored ink on the tail and were each acclimated to handling for 10 min/day during the 5 days before testing.
Apparatus
Testing was carried out in a square open-field arena (200 cm × 200 cm), enclosed with 50-cm-high opaque Plexiglas walls. The arena was placed in a temperature-controlled room (21°C). During testing, the room was in complete darkness; all light sources (e.g. indicator lights in the DVD player, camera and air conditioner) were sealed with opaque tape or covered in dark cloth. The room was illuminated with an infrared light source, with an 830-nm filter that emits light not visible to the rats (Tracksys, UK). A video camera (Ikegami B/W ICD-47E) was placed 2.5 m above the center of the arena, providing a top view that was recorded onto a DVD (Sony 169 GB). The geometry of the arena was then altered in some tests as follows. One, two, or three of the arena corners were duplicated by two vertical Plexiglas plates (30 cm wide and 50 cm high), transforming the arena corner into two adjacent inner corners with a protruding corner between them (Fig. 1). In two subsequent tests, one of the arena sides was extended outward by a 30 × 60 cm sector (Fig. 1), and then one corner was triplicated into three adjacent inner corners, with two protruding corners separating them by means of four vertical Plexiglas plates (30 × 50 cm each; Fig. 1).

Experimental design. Rows represent the two tested groups of rats; columns represent, from left to right, the repeated tests. Each square represents a top view of the arena walls. Each asterisk marks the location at which a rat was placed at the beginning of each test
Procedure
Rats were randomly assigned to one of two groups (n = 10 each), and each rat was tested five times over consecutive days (one test per day). Rats in group 1 were exposed repeatedly to the same arena, in order to assess the effect of repetitive exposures (Fig. 1, top row). Rats in group 2 were exposed to an arena that underwent geometrical changes, by duplicating an arena corner over the first three tests, extending the arena outward in the fourth test, and triplicating one arena corner for the fifth test (Fig. 1, bottom row).
At the beginning of each test, a rat was placed in the same corner (‘start corner’), facing the center of the arena (Fig. 1, marked with an asterisk). The behavior of the rat was then recorded for 20 min, after which it was returned to its cage. After each test, the arena was cloth-wiped with detergent. Testing took place during the dark period of the dark/light cycle, since rats are nocturnal and their activity is higher at this time.
Data acquisition and analysis
Spatial behavior of the rats was extracted from the video files by means of a tracking system (Ethovision XT). Data were extracted for the entire arena, for four equal quadrants of the arena, and for 15 × 15 cm zones in the arena. For each such division, we measured the time spent there, the number of visits, and the distance traveled by each rat in each of the arena zones during the 20 min of testing. Data were compared by a two-way ANOVA with repeated measures, followed by a Tukey post hoc test. Alpha level was set to 0.05.
Results
The impact of the global arena geometry on spatial behavior
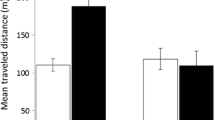
In both test groups, rats did not differ in the overall traveled distance (F 1,18 = 0.64; P = 0.435, Fig. 2). However, there was an effect of the repeated exposures (F 4,72 = 4.66; P = 0.002) and a significant interaction between groups and tests (F 4,72 = 2.57; P = 0.045). Tukey test revealed that when rats were exposed repeatedly to the same arena (group 1), there was no change in the traveled distance, whereas when the arena underwent geometric changes (group 2), the traveled distance increased in tests 4 and 5 compared to test 1. These findings indicate that the substantial alteration to the square shape of the arena (tests 4 and 5) resulted in a significant increase in the traveled distance.

The mean (±SEM) total distance traveled in group 1 (left) and group 2 (right). Repeated tests are depicted on the x-axis along with the respective arena shape. As shown, there was no significant difference in group 1, in which the arena was not altered. In group 2, there was no significant change in tests 1–3, where changes were local at the arena corners and the global square shape of the arena was preserved. However, there was a significant change (marked by *) in tests 4 and 5, when the arena underwent changes that distorted its square geometry. * Indicates a significant difference compared to test 1 of the same group
The impact of the global arena geometry is also evident in the rats’ paths (Fig. 3). In group 1 (Fig. 3a), where rats were exposed to the same arena, there was no apparent effect of the repetitive tests. This was not the case in group 2, for which arena geometry underwent progressive changes (Fig. 3b). As shown, the progressive increase in the number of corners in tests 1–3 does not seem to have affected the travel paths. However, in tests 4 and 5, in which the geometry of the arena underwent a major alteration that distorted its global square geometry, there is a notable change in the paths of the rats in the vicinity of the extended corner (Fig. 3b).

Spatial behavior in arena with fixed (group 1) compared with altered (group 2) geometry. Paths of locomotion are depicted for five exemplary rats (rows) over the five tests (columns) in group 1 (a) and group 2 (b). For each group, the respective arena shapes and its division into quadrants are depicted on the top row. As shown, differences in paths occurred mainly in tests 4 and 5 of group 2, where the square arena geometry underwent significant distortion
In order to quantify the impact of the alterations to the arena geometry on the rats’ spatial behavior, we divided the arena into four quadrants (dashed lines in the top row of Fig. 3a, b). For each rat, we then calculated the traveled distance and the time spent in each quadrant in each test. There was no significant difference in the distance traveled in the quadrants between the two groups (F 1,18 = 0.64; P = 0.435, Fig. 4). There was a significant difference between tests and between quadrants (F 4,72 = 4.66, P = 0.002; F 3,54 = 18.05, P < 0.001; respectively). In group 1, the traveled distance in each quadrant was steady over the repeated tests to the same arena geometry (Fig. 4, left). In contrast, there were marked changes in group 2, when arena geometry was modified over tests (Fig. 4, right). These changes were reflected in a significant interaction between groups, tests and quadrants (F 12,216 = 7.36; P < 0.001). Specifically, Tukey test revealed that in quadrants 1–3, where corners were successively duplicated, the traveled distance did not significantly change across tests 1–5. This is especially striking in the triplicated corner of quadrant 3 in test 5. For quadrant 4, where the arena was extended outward in tests 4 and 5, the distance traveled during these tests was significantly greater than in tests 1–3. These changes in the traveled distance were replicated for the time spent at each quadrant (data not shown).

The increase in traveled distance in the quadrant with distorted geometry. The mean (±SEM) traveled distance in each arena quadrant over repeated tests (x-axis) is shown for the arena with no geometric changes (group 1, left) and arena with altered corner and wall geometry (group 2, right). The division of the arena into quadrants is depicted at the top between the two graphs, and each line represents the change in the traveled distance in the quadrant with the respective color. As shown, in group 1, there was no change in the traveled distance over repeated tests, but this differed among quadrants, being greatest in the start quadrant ( ). In group 2, there was a significant change only in the traveled distance in quadrant 4 (
). In group 2, there was a significant change only in the traveled distance in quadrant 4 ( ), where the arena was extended outward
), where the arena was extended outward
The impact of the local arena geometry on spatial behavior
In order to analyze the effect of the local geometry, the arena perimeter was divided into 15 × 15 cm squares. Using these squares, it was possible to quantify the behavior at each of the arena corners (blue squares in Fig. 5) and compare it with the changes in corner geometry. For each corner, we calculated the mean number of visits (entries) to these squares, thus quantifying activity per area unit. We found that there was no significant difference between the two groups in the number of visits to each of the corners (F 1,18 = 0.28; P = 0.600). However, there was a significant difference between tests and between corners (F 4,72 = 3.32, P = 0.015; F 3,54 = 21.33, P < 0.001; respectively). Nevertheless, there was no significant interaction between groups, tests and corners (F 12,216 = 1.66; P = 0.077). Tukey test revealed that in group 1, the repeated tests did not significantly affect the number of visits to each of the corners. Similarly, in group 2, none of the progressive changes in the arena geometry affected the number of visits per square to each of the corners. The only significant difference was in the outward extension of corner 3 in test 5, where a higher number of visits per square were scored, compared to test 1. Note, however, that this corner was not changed in test 5 compared with test 4, and therefore this higher number of visits per square was not a mere reflection of the geometry of corner 3. Implicit in these results is that the overall increased activity when the arena shape was changed (tests 4 and 5) was not due to higher activity per square, but to the inclusion of additional such squares in the extended corner (corner 3).

The division of the corners and wall into comparable 15 × 15 cm squares. The division of the corners (dark squares) and wall (open squares) is shown for group 1 (top row) and group 2 (bottom row) for each of the tests (columns). This division enabled a comparison of the mean number of visits to a square in a corner or along a wall between the different groups and tests. As detailed in the text, the activity per area unit at the corners did not differ between the groups and the tests with the exception of group 2 that displayed a higher number of visits to corner 3 in test 5. The activity per area unit at the walls was not affected by the repeated tests in group 1. In contrast, in group 2, when corner 3 was extended outwards in tests 4 and 5 the number of visits per square to the right wall was higher compared to all other tests
The division of the arena perimeter into 15 × 15 cm squares was also utilized to assess travel along the arena walls (green open squares in Fig. 5), where another major component of locomotion took place. The mean number of visit per square at the top, left, right, and bottom walls was calculated for each test in groups 1 and 2. There was no significant difference between the groups in the number of visits per square at each of the arena walls (F 1,18 = 1.03; P = 0.325). There was also no significant difference between the tests (F 4,72 = 1.55; P = 0.197). However, there was a significant difference in the number of visits per square between the walls (F 3,54 = 21.96; P < 0.001), and a significant interaction between groups, tests, and walls (F 12,216 = 5.76; P < 0.001). Tukey test revealed that in group 1 there was no significant difference in the number of visits per square at each of the walls over repeated tests. In group 2, there was no significant difference in the visits per square at the top wall, which was not changed over tests. Surprisingly, there was also no difference in the number of visits per square at the left wall, where corner 2 was triplicated in test 5. The number of visits per square at the bottom wall was higher in test 5 compared to test 1. The number of visits per square at the right wall was also higher in tests 4 and 5 compared to all other tests; i.e., when corner 3 was extended outwards and the global shape of the arena was altered, there was an increase in activity at one of the walls that led to that corner.
Discussion
In the present study, we progressively altered the form of a square test arena by means of local changes at its corners and walls, in order to differentiate the impact of the global arena geometry from that of the local geometry of arena sectors on the rats’ spatial behavior. We found that as long as the global geometry of the environment did not undergo substantial changes and retained a square shape, the rats’ behavior did not significantly change. In contrast, distortion of the square shape of the arena resulted in significant changes in the spatial distribution of the rats’ activity. Specifically, activity increased at those specific arena sectors that, following arena alteration, were more closely surrounded by walls on three sides. In the following discussion, we suggest that these results support the notion that the perceived global geometry affects spatial behavior and overrides the impact of local geometry.
Home base at the start corner: the anchor for traveling in an unfamiliar homogenous (ambiguous) environment
When rats are introduced into an unfamiliar environment, they establish a ‘home base’ where they spend longer duration, and from where they set out on round trips in the environment (Eilam and Golani 1989). The home base is established at the start location, where rats are introduced into the arena (Nemati and Whishaw 2007), and in an empty square arena, the home base is typically established at one of the corners, probably because these are easy to recognize, and are relatively protected areas (Avni et al. 2006; Eilam 2004). Once a home base is established, paths of progression in the environment converge upon it (Avni et al. 2006; Geyer et al. 1986) and round trips in the environment are performed with a typical structure of a limited number of stops (Golani et al. 1993) and specific kinematics (Drai and Golani 2001; Dvorkin et al. 2008, 2010; Horev et al. 2007). In accordance with the aforementioned studies, rats in the present study established their home base at the start corner, as reflected in the longer duration of staying and the greater travel distance in the quadrant containing the start corner, as well as the high incidence of visits to that quadrant. These characteristics of the start quadrant were preserved over all tests regardless of the alterations to the arena shape. Accordingly, the home base at the start corner of the dark test arena was the anchor of spatial behavior in the tested rats, as also revealed in past studies.
The role of environment geometry in exploration and navigation
Studies on the spontaneous spatial behavior of several rodent species have revealed that a metric change in arena size does not affect open-field behavior (Eilam 2003; Eilam et al. 2003; Golani et al. 1993), as if the corners and walls of the open field are perceived as one geometric entity and not as a set of discrete walls and corners (Wang and Spelke 2002). For example, when rats were exposed to an ‘elastic’ arena expansion, in which the same rats were first exposed to a 1 × 1 m arena with four equispaced objects, and then exposed to a 2 × 2 m arena with four more distantly equispaced objects, they preserved in the large arena the same overall preference for certain objects that they had shown before arena expansion (Zadicario et al. 2007). Thus, past studies on spontaneous spatial behavior well fit the concept of a global geometric representation, following the prediction that “a given sketch will be recognized as such in spite of changes in size, although this might induce a general activity response” (Jacobs and Schenk 2003). Nevertheless, while it is well-accepted that humans and other animals encode and rely on the geometry of the environment (Cheng and Newcombe 2005; Cheng 2008; Vallortigara 2008), several concerns were raised regarding the relative importance of global geometry compared with local information, be it geometric (e.g. angles of corners), or non-geometric (featural, metric), and regarding the context in which environment geometry predominates the manifestation of spatial behavior (Cheng 2008; Chiandetti and Vallortigara 2008; Tommasi and Polli 2004). Specifically, the above past studies questioned whether the predomination of global geometry of space, as opposed to local information, is restricted to the scenario of Cheng’s seminal study, which was based on a reorientation task in a small arena with simple geometry. In other words, it was not clear whether geometry also bears on spatial behavior in environments with complex or ambiguous geometry, and in large environments. Another question relates to what shapes spatial behavior: the geometry of the surrounding environment or the geometry of specific locales/objects? The present findings are pertinent to these questions, as detailed below.
Local and global geometry
The present results demonstrated that freely moving rats did not change their spatial behavior following alteration to the test arena that, one by one, altered the local geometry of the arena corners by ‘duplicating’ each corner to form a protruding corner enclosed between two inner corners. Although this change involved the shortening of two adjacent arena walls by 15%, the orthogonal position of the walls was preserved. We expected that the distinctness of the altered corner against the background of the homogenous global arena geometry would attract the rats, thereby resulting in increased visit incidence, extended duration of stay, and greater traveled distance in the vicinity of the altered corner (Yaski and Eilam 2007). However, the successive geometric alterations to three of the arena corners did not result in change in the rats’ spatial behavior. We refer to the duplication of an arena corner as a local change since the global square shape of the arena was preserved, with the position and most of the walls’ length unchanged. Indeed, even with such local changes at three corners, the behavior of rats resembled that of the rats that were tested in the unchanged arena, attesting to the local geometric changes at the corners not having affected spatial behavior. Thus, if there was an effect of geometry in the present study, it was that of the global, not the local geometry.
Is there an effect of environment geometry on spatial behavior?
The lack of a significant effect of the local geometry on the rats’ spatial behavior raises the question of whether environment geometry is at all a factor in shaping spatial behavior. Specifically, it was suggested that spatial behavior in rats is primarily directed to optimize security (Whishaw et al. 2006; see however, Eilam 2010). Accordingly, one could assume that rats in the present study settled at the start corner, since a corner is a relatively protected location that is enclosed by two walls, and that the only change in spatial behavior occurred when other arena sectors were altered to become enclosed within three walls, thus becoming even more secure. While we cannot preclude such an interpretation, it is not likely that security alone controlled the rats’ spatial behavior in the present study, for the reasons detailed below.
Wild rats typically live in densely structured environments where they can better elude predators (Barnett 1968). In laboratory settings, the organization of their locomotor behavior is affected by the structure and geometry of the environment (e.g. Paulus and Geyer 1997), with rodents avoiding open spaces and clinging to the relatively secure apparatus walls (Eilam 2004; Eilam et al. 2003). Rats in the present study were tested in a dark and quiet environment. They also clung to the security of the arena walls, but also moved throughout the environment, with numerous crosscuts through the arena center (Fig. 3). In the same vein, other studies have demonstrated that rodents may divert from optimal security by abandoning their shelter (Eilam 2010) or by frequently visiting a prominent object placed in an exposed location (Yaski and Eilam 2007). Notably, in the present experiment, in corner 3, which was altered to be “safer” by being enclosed by walls on three sides, no change in spatial behavior occurred when we altered that corner (test 4) but only in the subsequent test, when the same corner remained unaltered, but the arena geometry was distorted by altering another corner (corner 2 in test 5). We therefore suggest that spatial behavior of the rats in the present study was, at least partially, influenced by arena geometry. Moreover, since the local geometry of specific locales did not seem to shape the behavior of these rats, we conclude that it is the global geometry that is involved in shaping spatial behavior.
Global geometry: instantly perceived or gradually composed by integrating local geometries?
One drawback in laboratory test environments is that of their simple geometry. For example, the water maze is usually a round pool, and the open field is usually a square or rectilinear enclosure. Whereas simple geometry was critical in establishing the concept of ‘geometric module’ (Cheng 1986; Cheng and Newcombe 2005; Vallortigara 2008), enclosures do not necessarily reflect what animals and humans encounter in their natural environment. Indeed, habitats are likely to provide various arrays and shapes of landmarks. Moreover, the notion of geometric module has been refuted in studies with complex geometry (Cheng 2008; Pearce et al. 2004; Stürzl et al. 2008). In the present study, rats were tested in a square arena, which constitutes a simple yet an ambiguous, “over-symmetrical” environment shape; that is, a shape that is congruent with itself for rotations of 90° for a square (Benhamou and Poucet 1998). However, our test procedure breached this ambiguity and symmetry of the square shape. Specifically, the rats seemed to display a preference to travel along the left compared with the top wall, even when these were geometrically symmetrical (Fig. 2). In addition, placing the rats at the same start corner resulted in their preference for the start quadrant, and subsequently in differential activity at different quadrants (Fig. 4). Thus, environment ambiguity and symmetry do not seem to affect the present finding on the predominance of the global over the local geometry.
Environment size also matters in perceiving its geometry (Chiandetti et al. 2007; Sovrano et al. 2005, 2007). If the test environment is complex or large enough to prevent visual encompassing of its overall global shape, then it is virtually impossible to first encode the global geometry and then paste additional featural information onto it. In the present study, rats were tested in a dark arena, in which visual information was absent. This resembles testing in a large arena, since in both situations the navigators cannot immediately encode the global geometry, and therefore need to encode alternative information. One such alternative is that of space calibration. Specifically, we have demonstrated that in an unfamiliar environment, blindfolded humans and blind mole rats (Spalax ehrenbergi) first calibrate the environment by going through a buildup phase of constructing a perimeter path (Avni et al. 2008; Yaski et al. 2009). Humans in this phase also counted steps and were then able to accurately plot and describe the global geometry of the environment along with its dimensions (Yaski et al. 2009). After both the blindfolded humans and the blind mole rats had completed the perimeter path, they evidently integrated the local information into a global representation that they then utilized for crosscutting through the unexplored environment center (Avni et al. 2008; Yaski et al. 2009). Thus, when a view of the entire environment is not available, as in the case of large or dark environments, the navigator has first to acquire local geometries and then to compose them into a global geometric shape of the overall environment.
Constructing the perimeter path, which is a phase of acquiring the global shape and size of the environment (Avni et al. 2008; Yaski et al. 2009), is based on gaining local familiarity with the environment, sector by sector. Specifically, in a previous experiment with mole rats, they gradually extended their perimeter path by repeatedly retracing their previous paths. They gained local familiarity with the arena corners, and ultimately, when the perimeter path was completed, they integrated the local information into a global representation that they utilized for crosscutting through the unexplored environment center while slowing down when they reached to the vicinity of the opposite wall (Avni et al. 2008). Exploration by another rodent species, the jird (Meriones tristrami; a gerbil-like rodent), in a dark arena consisted of an initial period of traveling to a recently visited location in loop-like paths, and only then, after encompassing the entire arena, in establishing a home base (Avni et al. 2006; Zadicario et al. 2005). For the present discussion, these examples illustrate that the initial phase of exploration may involve the acquisition of local and non-geometric information, and only then a perception of the global geometry of the environment. This calls the attention to the exploration phase during which the sense of environment geometry is acquired and thereby affects spatial behavior. The present findings reinforce the predominance of the effect of global geometry on the behavior of the rats after 20 min of travel in the environment and over repeated exposures to that environment. While it is unlikely that these rats could have first perceived the entire global geometry of the dark arena, their spatial behavior later on seemed to be predominated by the global geometry.
Summary and conclusion
Rats in the present study were repeatedly introduced into a dark arena subjected to local geometric alterations across the repetitive exposures. Since the rats could not visually perceive the shape of the arena, they had to learn this by traveling and exploring the arena sector by sector. We found that the rats established their home base at the start corner in which they had been introduced into the arena. From the start corner, they then traveled spontaneously in the arena, in a pattern that remained constant as long as the global square and symmetrical shape of the arena was preserved. In other words, despite local geometrical alterations at the arena corners and an increase in the number of corners, the rats did not change their spatial behavior. Such behavioral change occurred only when the arena was altered to distort the square shape. It is thus suggested that spatial behavior is organized in relation to the global, not local geometry.
References
Alyan SH (1996) Evidence for resetting the directional component of path integration in the house mouse (Mus musculus). Ethology 102:629–638
Avni R, Zadicario E, Eilam D (2006) Exploration in a dark open field: a shift from directional to positional progression and a proposed model of acquiring spatial information. Behav Brain Res 171:313–323
Avni R, Tzvaigrach Y, Eilam D (2008) Exploration and navigation in the blind mole rat (Spalax ehrenbergi): global calibration as a primer of spatial representation. J Exp Biol 211:2817–2826
Barnett SA (1968) The rat: a study in behaviour. Aldine, Chicago
Benhamou S, Poucet B (1998) Landmark use by navigating rats (Rattus norvegicus): contrasting geometric and featural information. J Comp Psychol 112:317–322
Cheng K (1986) A purely geometric module in the rat’s spatial representation. Cognition 23:149–178
Cheng K (2008) Whither geometry? Troubles of the geometric module. Trends Cogn Sci 12:355–361
Cheng K, Newcombe NS (2005) Is there a geometric module for spatial orientation? Squaring theory and evidence. Psychon Bull Rev 12:1–23
Chiandetti C, Vallortigara G (2008) Spatial reorientation in large and small enclosures: comparative and developmental perspectives. Cogn Process 9:229–238
Chiandetti C, Regolin L, Sovrano VA, Vallortigara G (2007) Spatial reorientation: the effects of space size on the encoding of landmark and geometry information. Anim Cogn 10:159–168
Collett TS (1996) Insect navigation en route to the goal: multiple strategies for the use of landmarks. J Exp Biol 199:227–235
Doeller CF, King JA, Burgess N (2008) Parallel striatal and hippocampal systems for landmarks and boundaries in spatial memory. Proc Natl Acad Sci USA 105:5915–5920
Drai D, Golani I (2001) SEE: a tool for the visualization and analysis of rodent exploratory behavior. Neurosci Biobehav Rev 25:409–426
Dvorkin A, Benjamini Y, Golani I (2008) Mouse cognition-related behavior in the open-field: emergence of places of attraction. PLoS Comput Biol 4:e1000027
Dvorkin A, Szechtman H, Golani I (2010) Knots: attractive places with high path tortuosity in mouse open field exploration. PLoS Comput Biol 6:e1000638
Eilam D (2003) Open-field behavior withstands drastic changes in arena size. Behav Brain Res 142:53–62
Eilam D (2004) Locomotor activity in common spiny mice (Acomys cahirinuse): the effect of light and environmental complexity. BMC Ecol 4:16
Eilam D (2010) Is it safe? Voles in an unfamiliar dark open-field divert from optimal security by abandoning a familiar shelter and not visiting a central start point. Behav Brain Res 206:88–92
Eilam D, Golani I (1989) Home base behavior of rats (Rattus norvegicus) exploring a novel environment. Behav Brain Res 34:199–211
Eilam D, Dank M, Maurer R (2003) Voles scale locomotion to the size of the open-field by adjusting the distance between stops: a possible link to path integration. Behav Brain Res 141:73–81
Etienne AS, Maurer R, Georgakopoulos J, Griffin A (1999) Dead reckoning (path integration), landmarks, and representation of space in a comparative perspective. In: Golledge RG (ed) Wayfinding behavior: cognitive mapping and other spatial processes. The Johns Hopkins University Press, Baltimore, pp 197–228
Etienne AS, Boulens V, Maurer R, Rowe T, Siegrist C (2000) A brief view of known landmarks reorientates path integration in hamsters. Naturwissenschaften 87:494–498
Gallistel CR (1989) Animal cognition: the representation of space, time and number. Annu Rev Psychol 40:155–189
Gallistel CR (1990) The organization of learning. MIT Press, New York
Gauzzelli A, Bota M, Arbib MA (1999) Incorporating path integration capabilities in the TAM-WG model of rodent navigation. Neurocomputing 26–27:713–719
Geyer MA, Russo PV, Masten VL (1986) Multivariate assessment of locomotor behavior—pharmacological and behavioral analyses. Pharmacol Biochem Behav 25:277–288
Golani I, Benjamini Y, Eilam D (1993) Stopping behavior: constraints on exploration in rats (Rattus norvegicus). Behav Brain Res 53:21–33
Golledge RG (1999) Human cognitive maps and wayfinding. In: Golledge RG (ed) Wayfinding behavior: cognitive mapping and other spatial processes. Johns Hopkins University Press, Baltimore, pp 5–45
Hamilton DA, Akers KG, Weisend MP, Sutherland RJ (2007) How do room and apparatus cues control navigation in the Morris water task? Evidence for distinct contributions to a movement vector. J Exp Psychol Anim Behav Process 33:100–114
Hamilton DA, Akers KG, Johnson TE, Rice JP, Candelaria FT, Sutherland RJ, Weisend MP, Redhead ES (2008) The relative influence of place and direction in the Morris water task. J Exp Psychol Anim Behav Process 34:31–53
Hartley T, Trinkler I, Burgess N (2004) Geometric determinants of spatial memory. Cognition 94:39–75
Horev G, Benjamini Y, Sakov A, Golani I (2007) Estimating wall guidance and attraction in mouse free locomotor behavior. Genes Brain Behav 6:30–41
Jacobs LF, Schenk F (2003) Unpacking the cognitive map: the parallel map theory of hippocampal function. Psychol Rev 110:285–315
Lynch K (1960) The image of the city. MIT Press, Massachusetts
Maaswinkel H, Whishaw IQ (1999) Homing with locale, taxon, and dead reckoning strategies by foraging rats: sensory hierarchy in spatial navigation. Behav Brain Res 99:143–152
Maurer R, Derivaz V (2000) Rats in a transparent morris water maze use elemental and configural geometry of landmarks as well as distance to the pool wall. Spat Cogn Comp 2:135–156
Mittelstaedt ML, Mittelstaedt H (1980) Homing by path integration in a mammal. Naturwissenschaften 67:566–567
Nemati F, Whishaw IQ (2007) The point of entry contributes to the organization of exploratory behavior of rats on an open field: an example of spontaneous episodic memory. Behav Brain Res 182:119–128
Paulus MP, Geyer MA (1997) Environment and unconditioned motor behavior: influences of drugs and environmental geometry on behavioral organization in rats. Psychobiol 25:327–337
Pearce JM, Good MA, Jones PM, McGregor A (2004) Transfer of spatial behavior between different environments: implications for theories of spatial learning and for the role of the hippocampus in spatial learning. J Exp Psychol Anim Behav Process 30:135–147
Pecchia T, Vallortigara G (2010) View-based strategy for reorientation by geometry. J Exp Biol 213:2987–2996
Shettleworth SJ (1998) Cognition, evolution, and behavior. Oxford University Press, New York
Shettleworth SJ (2005) Taking the best for learning. Behav Processes 69:147–149
Shettleworth SJ, Sutton JE (2005) Multiple systems for spatial learning: dead reckoning and beacon homing in rats. J Exp Psychol Anim Behav Process 31:125–141
Sovrano VA, Bisazza A, Vallortigara G (2005) Animals’ use of landmarks and metric information to reorient: effects of the size of the experimental space. Cognition 97:121–133
Sovrano VA, Bisazza A, Vallortigara G (2007) How fish do geometry in large and in small spaces. Anim Cogn 10:47–54
Stackman RW, Herbert AM (2002) Rats with lesions of the vestibular system require a visual landmark for spatial navigation. Behav Brain Res 128:27–40
Stürzl W, Cheung A, Cheng K, Zeil J (2008) The information content of panoramic images I: the rotational errors and the similarity of views in rectangular experimental arenas. J Exp Psychol Anim Behav Process 34:1–14
Tchernichovski O, Benjamini Y, Golani I (1996) Constraints and the emergence of ‘free’ exploratory behavior in rat ontogeny. Behaviour 133:519–539
Tchernichovski O, Benjamini Y, Golani I (1998) The dynamics of long-term exploration in the rat—part I. A phase-plane analysis of the relationship between location and velocity. Biol Cybern 78:423–432
Tolman EC (1948) Cognitive maps in rats and men. Psychobiol Rev 55:189–208
Tommasi L, Polli C (2004) Representation of two geometric features of the environment in the domestic chick (Gallus gallus). Anim Cogn 7:53–59
Vallortigara G (2008) Animals as natural geometers. In: Tommasi L, Nadel L, Peterson M (eds) Cognitive biology: evolutionary and developmental perspectives on mind, brain and behavior. MIT press, London, pp 83–104
Wallace DG, Hines DJ, Whishaw IQ (2002) Quantification of a single exploratory trip reveals hippocampal formation mediated dead reckoning. J Neurosci Methods 113:131–145
Wang RF, Spelke ES (2002) Human spatial representation: insights from animals. Trends Cogn Sci 6:376–382
Whishaw IQ, Gharbawie OM, Clark BJ, Lehmann H (2006) The exploratory behavior of rats in an open environment optimizes security. Behav Brain Res 171:230–239
Yaski O, Eilam D (2007) The impact of landmark properties in shaping exploration and navigation. Anim Cogn 10:415–428
Yaski O, Eilam D (2008) How do global and local geometries shape exploratory behavior in rats? Behav Brain Res 187:334–342
Yaski O, Portugali J, Eilam D (2009) The dynamic process of cognitive mapping in the absence of visual cues: human data compared to animal studies. J Exp Biol 212:2619–2626
Zadicario P, Avni R, Zadicario E, Eilam D (2005) ‘Looping’—an exploration mechanism in a dark open field. Behav Brain Res 159:27–36
Zadicario P, Ronen S, Eilam D (2007) Modulation of quinpirole-induced compulsive-like behavior in rats by environmental changes: implications for OCD rituals and for exploration and navigation. BMC Neurosci 8:23
Acknowledgments
The present study complies with the current law in Israel and the institutional guidelines for animal care and use in research (permit #L-10-013). The authors declare that they have no conflict of interest. This study was supported by The Israel Science Foundation, Grant 177/09.
Author information
Authors and Affiliations
Corresponding author
Additional information
The authors D. Ben-Yehoshua and O. Yaski contributed equally to the present research.
Rights and permissions
About this article
Cite this article
Ben-Yehoshua, D., Yaski, O. & Eilam, D. Spatial behavior: the impact of global and local geometry. Anim Cogn 14, 341–350 (2011). https://doi.org/10.1007/s10071-010-0368-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10071-010-0368-z