
Abstract
The relationships between climate and forest dynamics can help us to interpret patterns of ecosystem carbon and to predict how forests react to climatic changes. We report mass dynamics of deadwood (necromass) from tropical forest ecosystems subject to some of the highest frequency of tropical cyclones in the world and to regular, persistent seasonal monsoon winds. Plots that are influenced by typhoons but exposed to different degrees of monsoon winds were monitored. We expected that stocks and inputs of necromass would reflect the seasonal intensity of wind events and be higher in the high wind exposure forest than in the low wind exposure forest, especially for fallen woody debris. The results showed that necromass input was indeed influenced by the magnitude of typhoons and aggravated by monsoon winds. However, while there was no significant difference in stock of necromass between plots, inputs of standing necromass were significantly higher in the high wind exposure plot; these were mostly derived from dead resprouts. Both our forests had very low values of total necromass stocks (3.47–4.32 Mg C ha−1) and inputs (2.1–2.5 Mg C ha−1 y−1) compared with tropical forests worldwide. Our results show that both monsoon and typhoon winds shape these tropical forests, favouring low stature individuals and trees with ability to resprout and that these strategies provide these forests with remarkable resistance and resilience to wind disturbances. Our findings from some of the most wind-affected forests in the world indicate how woody carbon dynamics and forest structure in other regions may respond to future changes in the frequency and intensity of winds.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Highlights
-
Locally, wind strength regulates the seasonal inputs of tropical forest necromass.
-
Globally, winds result in low-carbon stocks in both biomass and necromass in tropical forests.
-
Key resistance and resilience mechanisms to winds are short stature and resprouts.
Introduction
Tropical forests are vital ecosystems for storing and sequestering carbon (Pan and others 2011; Quéré and others 2018). The three major carbon pools in forest ecosystems are biomass (above- and below-ground living plants), necromass (litter and woody debris (deadwood)), and soil organic carbon. Although all pools contribute to carbon storage (stocks) and carbon sequestration (input and output fluxes), most attention is usually given to aboveground biomass (IPCC 2006; Köhl and others 2015; Clark and others 2017). The fluxes between each of these carbon pools are critical for determining the overall carbon storage and budgets in the ecosystem. Woody debris inputs into the necromass pool and losses of necromass are among these key carbon fluxes (Trumbore 2006; Palace and others 2012), and given the increased rates of tree mortality in some forests (for example, McDowell and others 2018) necromass stocks may also increase (Brienen and others 2015). Therefore, by neglecting to account for the necromass pool and associated fluxes, we incompletely assess the carbon balance in tropical forests and may not properly represent its true climate sensitivity (compare Sullivan and others 2020).
Although forest necromass stocks are in practice highly correlated with biomass (Chao and others 2009a), they should also reflect the balance between necromass fluxes—its input (necromass production) and its output (necromass decomposition). Necromass input is contributed by the quantity of woody debris produced by both tree death and branch fall (van der Meer and Bongers 1996; Palace and others 2008), with the causes of necromass input being primarily senescence, competition, stress, and disturbance (Franklin and others 1987). The output of necromass is mainly controlled by environmental factors, such as temperature (Chambers and others 2000; Berbeco and others 2012), and less so by the local biodiversity of decomposers and tree species traits (Pietsch and others 2019). As annual temperatures normally only change fractionally, the net balance between input and output of necromass is likely to be most strongly determined by the frequency and magnitude of the disturbances that can cause tree death and branch fall.
Examining different types of woody debris can indicate the causes of tree death (Chao and others 2009b). In general, the standing mode of death is related to senescence, competition, or stress, whereas the fallen mode of death is strongly associated with physical disturbance (Chao and others 2009b; Esquivel-Muelbert and others 2020). For example, typhoons are likely to increase the number of fallen branches and uprooted trees (Whigham and others 1991), and trees may also die snapped in wind events due to structural imbalances between root anchorage and stem strength (Soethe and others 2006). Therefore, understanding the types and magnitudes of woody debris present in forests can help indicate the major characteristics and mechanisms of woody debris dynamics.
Tropical cyclones are a major class of wind disturbance affecting many tropical forests. Although research on the effects of cyclones on tropical forests has been focussed strongly on the North Atlantic Basin, the much less studied north-western Pacific basin is the most active region on Earth for tropical cyclones (Lin and others 2020). As a tropical and subtropical island in the north-western Pacific, Taiwan experiences one of the highest typhoon disturbance rates of any landmass in the world (Lugo 2008; Lin and others 2010; Lin and others 2020) so forests here are likely to be among those most affected by tropical cyclones. As well as typhoons, though, the tropical north-western Pacific is also influenced by northeast monsoons in winter (Wang 2004). The prevailing monsoon winds are known to influence forest structure, such that monsoon-windward forests can be characterised by low canopy height and high stem density (Lawton 1982; Chao and others 2010b) (Appendix 1ab). Indeed, one patch of monsoon-windward forest that we monitor in Taiwan (Lanjenchi Plot) has one of the highest stem densities among forests globally (Lutz and others 2018). However, the effects of typhoons and monsoon winds on necromass dynamics here remain poorly understood, and by accounting for the effects of these two distinctive climatic processes on necromass dynamics, it will be possible to understand forest ecosystem carbon dynamics and their climate sensitivities better. Furthermore, as the patterns of typhoons and monsoons are changing (Xu and others 2006; Tu and others 2009; Jiang and Tian 2013; Lin and Chan 2015), evaluating the potential ecological effects of these changes on a global scale requires long-term monitoring of forest localities with pre-existing influences of typhoons and monsoons.
This study aims to investigate the patterns of necromass dynamics and balance in two forest ecosystems affected by both typhoons and monsoon winds: one located in the high-wind-exposed slopes of a coastal mountain (Appendix 1ab) and the other located in a relatively low-wind-exposed valley (Appendix 1 cd) (Figure 1a). At the landscape scale, the two forests experience a different degree of influence from the unidirectional, long-lasting, northeast winter monsoon (Figure 1). However, at the regional scale, both forests have a similar probability to being disturbed by short-period typhoons with unpredictable wind directions in summer (Figure 1). In the light of these differences and the fact that the study plots are close to each other (< 3 km apart, compared to the average 200 to 300 km radius of typhoons), we propose the following general hypothesis and specific predictions.

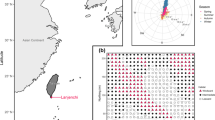
a Map and b sustained wind patterns of the study plots. a The study plots include high wind exposure (Lanjenchi plot) and low wind exposure plot (Nanjenshan plots I and II). The high wind exposure plot is on the first mountain range facing the northeast monsoon, whereas the low wind exposure plot is located on the valley between the first and second mountain ranges. Inset map on the right shows Taiwan, with a yellow rectangle indicating the location of the study landscape. b Sustained wind speed was measured at 2 m above ground of the high wind exposure plot and the low wind exposure plot, respectively. The typhoon season (mid-July to mid-October) is indicated in light grey and the monsoon season (mid-October to mid-February) in light blue. Lower and upper case letters indicate, respectively, significant differences between seasons and plots using Tukey’s HSD post hoc test.
Hypothesis: The combined effect of these winds results in unusually high accumulation (stocks) and production (inputs) of woody debris. Our expectations are:
-
1.
Necromass stock
-
1.1
Necromass stocks are greatest in localities exposed more to winds.
-
1.1
-
2.
Necromass input
-
2.1
Necromass inputs are higher in localities exposed more to winds.
-
2.1
Fallen woody debris inputs are mainly determined by the magnitude of winds, whereas standing woody debris inputs are unaffected by these.
Methods
Study Area
The first study plot, Lanjenchi plot (5.88 ha; 220 m by 240 to 300 m; 120° 51′ 38'' E, 22° 03′ 23'' N), is located on coastal mountain upper slopes and hilltops, which are exposed to northeast monsoon wind (Chao and others 2010b) (Figure 1a; hereafter the high wind exposure plot). The second set of study plots, Nanjenshan plots I (2.1 ha; 150 m by 140 m; 120° 50′ 51'' E, 22° 04′ 54'' N) and II (0.64 ha; 80 m by 80 m; 120° 50′ 36'' E, 22° 04′ 52'' N), is located on the valley of the same mountain range, which has low exposure to monsoon wind (Figure 1a) (Chao and others 2010b). The floristic composition of the Nanjenshan plots is very similar to one another (Chao and others 2010a), so results from Nanjenshan plot I and plot II are here treated together as one sample, denoted the ‘Nanjenshan’ or ‘low wind exposure plot’.
Although the two forests are close together and their elevations differ little, their topographic characteristics, average wind speed (sustained and gust), and vegetation types and structure are all significantly different (Figure 1; Table 1; Appendix 2). The high wind exposure plot (Lanjenchi) is situated on the upper slope of a low elevation coastal mountain range, directly facing the Pacific Ocean (Figure 1a). Due to its lacking any protection from other mountain ranges, the median sustained wind speed in the high wind exposure plot during the winter monsoon season is high (Figure 1b; methods please see Appendix 2), which was proposed to be the major causal factor of the high stem density and low aboveground biomass in the plot (Table 1) (Chao and others 2010b). The low wind exposure plot (Nanjenshan) is further inland situated in a valley protected by two mountain ranges. The slopes of the low wind exposure plot mostly face north-east and north-west, but are less influenced by the winter monsoon due to mountain ranges to the north-east (Figure 1a).
The wind regimes of the plots are a complex of summer typhoons and winter monsoon winds (Figure 1b). In the monsoon season, the high wind exposure plot has higher median sustained wind speed (Figure 1b). In typhoon season, because typhoon wind directions are unpredictable, both plots can be affected by typhoons. Therefore, in some years the maximum sustained wind speed can be high in typhoon season in the low wind exposure plot (Figure 1b). A preliminary analysis showed that there are significant differences in sustained wind speed between plots (two-way ANOVA using log10-transformed wind speed, F1,374256 = 47,180, p < 0.001) and between seasons (F3,374256 = 9567, p < 0.001). Similar patterns can also be found when examining gust wind speeds between months (Appendix 2).
Necromass Stock and Input
Woody debris was defined as those dead woody branches or trunks of plants with diameters ≥ 1 cm. In the high wind exposure plot, three east to west transects (200 m, 200 m, and 198 m) were established in February 2012, and two north-to-south transects (280 m and 190 m) were established in February 2013. In the low wind exposure plot, eight transects were established in February 2013 (five in plot I (111 m, 105 m, 105 m, 100 m, and 105 m) and three in plot II (64 m, 60 m, and 60 m)). Three of the transects are oriented east to west and five run north to south. A total of 1778 m were sampled in the study plots (Liao 2017). Since establishment, investigation of newly formed woody debris (necromass input) was conducted seasonally every three months until February 2018. In the meantime, necromass stocks were censused annually. The census times fall close to the beginning of February, April, July and October, but with adjustments of up to ± 30 days in some cases. Necromass stock and input were quantified and analysed for a total of 6 years in the high wind exposure plot and 5 years in the low wind exposure plot.
A line-intersect method (van Wagner 1968) was used to investigate fallen woody debris on the transect lines. A plot method (Harmon and Sexton 1996) was used to sample standing woody debris within 5 m on each side of the transect lines. For fallen woody debris pieces on the intersect lines, their diameters (d) were measured and void proportion was estimated. For those standing woody debris pieces (including dead resprouts of multi-stemmed individuals) located within 5 m on each side of the intersect lines, their diameters at the base (db) and top (dt), and their height (Ls (m)) were measured and void proportion was estimated. All measured woody debris pieces were numbered and tagged with nylon strings to identify samples of each census. Woody debris was classified into five decay classes, such that decay class 1 indicates a piece of intact wood and decay class 5 indicates a piece of rotten wood (Chao and others 2017).
Volumes and carbon mass of fallen and standing woody debris were estimated, respectively. (1) Volumes of fallen woody debris pieces (vf) were estimated by
based on the line-intersect method (van Wagner 1968), where vf is the volume at the unit area (m3 ha−1), d is the intercepted diameter (cm) of each fallen woody debris piece and L in the total length (m) of each transect. The equation assumes that each sample line will cross woody debris at various angles, making a set of vertical elliptical cross-sections. Once integrated, the cross-sectional area per unit length (cm2 m−1) can be used to estimate woody debris volume per unit area (m3 ha−1). If there is any void proportion noted for a particular piece of woody debris, its actual volume is multiplied by (1–void proportion). (2) Volumes of each standing woody debris piece were estimated by Smalian’s formula (Phillip 1994):
where vs is the volume (m3) of the target standing woody debris, db and dt (m) are the diameters at base and top, respectively, and LS (m) is the length of the target standing woody debris. If there is any void proportion noted for a particular piece of woody debris; its actual volume is multiplied by (1–void proportion). The standing woody debris volume per unit area (m3 ha−1) was computed as the sum of the total volume of standing woody debris sampled in a transect divided by the transect area and standardised to a per hectare value. The averages of the plot-level volumes were weighted by transect length (Keller and others 2004). Plot-level variance (σ2) values were also weighted by transect length as suggested by Keller and others (2004).
where Lj is the length of each transect; vij (m3 ha−1) is the measured volume (either standing or fallen woody debris) of each transect j at the decay class i; \(~\bar{v}_{i}\) is the weighted average of each plot at the decay class i; n is the number of sampled transects. Standard error of the mean (SE) was calculated as σ/\(\sqrt n\). Plot-level SE is the sum of each SE at each decay class (Chao and others 2017).
Field measurement of woody debris volume (vi) of woody debris (either standing or fallen) at decay class i can be converted to necromass carbon (NC, Mg C ha−1) by multiplying with woody debris density (ρ, g cm−3) and carbon concentration (c, g g−1 (carbon fraction)) at each decay class (i).
In this study, we applied the woody debris density and carbon concentration at each decay class reported in Chao and others (2017).
All newly encountered woody debris pieces were tagged and recorded as new input. Necromass input was calculated for each census based on the necromass carbon of newly recorded pieces of woody debris (either standing or fallen) at that census divided by the number of days between censuses (NCI=NC/number of days). As the number of days varied from 49 to 128 days, necromass input rates were standardised to annual equivalents (NCI, Mg C ha−1 y−1). Preliminary results showed that fine woody debris (diameter < 5 cm and ≥ 1 cm (FWD)) contributed disproportionally to the number of pieces of necromass but represented relatively small carbon stocks (Appendix 3). Thus, after July 2016, we only measured intermediate (≥ 5 cm; IWD) and coarse woody debris (≥ 10 cm; CWD). To account for the fine woody debris, estimates of total necromass after July 2016 were adjusted by the plot level and woody debris type average ratios of fine to other woody debris (intermediate and coarse), based on censuses between February 2012 and April 2016.
Necromass Decomposition and Net Fluxes
Decomposition constant of necromass (k) was investigated for one year in each forest (April 2012 to April 2013 in the high wind exposure plot; April 2013 to April 2014 in the low wind exposure plot). In each plot, we set up 16 quadrats (each 10 m × 10 m) separate from the woody debris transects and measured the diameter of each coarse woody debris piece (≥ 10 cm) at both ends. Additionally, decomposition class, length, and proportion of void space were recorded (Liao 2017). Each coarse woody debris piece (≥ 10 cm) was numbered and tagged with a nylon string. Within each quadrat, a subquadrat (2 m × 2 m in the high wind exposure plot; 1 m × 1 m in the low wind exposure plot) was set up in the south-west corner to investigate the fine and intermediate woody debris (≥ 1 cm, < 10 cm). The diameters, decomposition class, and length of each fine and intermediate woody debris were also recorded. To distinguish the remaining woody debris from newly fallen pieces at the next census, we covered these measured fine and intermediate woody debris pieces with fishnets. The necromass carbon of each woody debris pieces at the beginning of the census (Y0, Mg of carbon) in the selected quadrats is calculated using Eq. 2 and Eq. 4. Each woody debris piece was revisited a year later (t = 1 y), and each parameter measured again to calculate the necromass at the end of the census (Yt, Mg of carbon). The negative single-exponential decay equation was applied to calculate the decomposition constant (k, y−1) (Olson 1963):
Thereafter, with the annual necromass census data and the decomposition constant, the annual decomposition quantity of necromass carbon (NCD) of a specific year (t) can be estimated as
where NCS,t and NCS,t+1 (Mg ha−1) is the stock of necromass at time t and remaining stock at t+1. Note that the NCS,t+1 here was calculated based on the decomposition constant k, rather than from our direct field measurement at time t+1, because the direct field measurement at time t+1 would also include inputs of new woody debris.
The net flux of necromass is the difference between the annual input quantity (April, July, October of year t and February of year t+1) of necromass carbon and annual decomposition quantity of necromass carbon at time t (calculated based on measurement of year t).
Climate and Wind Disturbance
Climatic data, including the number of typhoons, sustained wind speed, and precipitation, were extracted from the records recorded in the Hengchun Station (No. 467590; about 20 km away from the study plots) in the Central Weather Bureau climate database (Central Weather Bureau 2019). We used the power dissipation index (PDI) (Emanuel 2005; Yu and Chiu 2012), including annual wind (PDIannual), seasonal wind (PDIseasonal) and typhoon (PDItyphoon), to evaluate the effect of winds. Only one weather station dataset was used for the PDI indices because the Hengchun station records the required maximum sustained wind speed data at 10 m above ground as proposed by Emanuel (2005). This provides a background regional magnitude of winds, rather than the local sustained wind speed data at 2 m height (those in Figure 1a and Appendix 2).
PDI (109 m3 s−2) was first proposed by Emanuel (2005) to estimate the power and magnitude of typhoon winds. The original equation is as follows:
Vmax (m s−1) is the maximum sustained wind speed at 10 m and integrated over lifetime τ (in units of second) of the typhoon. The Vmax in our study was the 6-hourly maximum sustained value of each typhoon (the Vmax in PDItyphoon) or daily-maximum sustained wind speed of each day (the Vmax in PDIannual and PDIseasonal). For PDItyphoon, we included each typhoon that the Central Weather Bureau had issued warning reports for Taiwan island (Central Weather Bureau 2019). Although warning reports for Taiwan may not always relate to a visit of typhoons to the study forests, it provides a consistent basis for extracting wind speed data that reflect the approximate magnitude of each typhoon on the study region.
In the literature, PDI is used only for evaluating the magnitude of typhoons. Here we applied the same index to evaluate the effects of prevailing winds throughout the year. In annual and seasonal PDI of this study, the Vmax was the daily-maximum sustained wind speed of each day (compare the 6-hourly maximum sustained wind data used in the typhoon PDI index) and integrated over each woody debris census study period. A trapezoidal rule was applied to approximate the results of annual and seasonal PDI (109 m3 s−2).
where ∆t represents the total time (in seconds) of the study period. Our calculations of PDIannual and PDIseasonal overestimate the magnitude of wind of each day due to only daily Vmax being used (compared to PDItyphoon). However, as 6-hourly data are not readily available in the study forests, the estimation can provide a consistent index of relative wind magnitude throughout our study period.
Data analysis
Linear mixed-effect models were used to examine the relationships between necromass and variables. The package lme4 (Douglas and others 2015) in the program R (R Core Team 2019) was applied. We use each transect as the random intercept effect (denoted as 1|transect) to account for repeat measurements of each transect. Other factors, including plot, PDI, number of typhoons, and precipitation, were used as fixed effects. If dependent or independent variables were not normally distributed when constructing the models, we transformed the variables to have an approximately normal distribution pattern. Where variables include the value 0, which cannot be log-transformed, we added a small fixed value of 0.1 or 0.01, depending on which leads to a better approximation to a normally distributed pattern. Sample-size corrected Akaike information criterion (AICc) values were used for model comparison (Burnham and Anderson 2002), such that the model with the lowest AICc value was considered to be the best model (that is, AICc differences (△AICc) = 0).
Result
Quantity of Necromass Carbon Stock
The total necromass stock was 3.47 ± 0.32 Mg C ha−1 (average ± SE) from 2012 to 2018 in the high wind exposure plot and 4.32 ± 0.43 Mg C ha−1 from 2013 to 2018 in the low wind exposure plot (Appendix 4). The annual variation of total stock reflects the patterns of the fallen stock rather than the standing stock (Figure 2). Standing stock was significantly higher in the low wind exposure plot than the high wind exposure plot (Mann–Whitney U test, p = 0.014; Appendix 5c), but plots were indistinguishable in terms of total and fallen stocks (p = 0.101, p = 0.836, respectively; Appendix 5ab). When controlling for the differences of transects, none of the investigated variables helped explain the patterns of the total, fallen, and standing stocks, as their AICc values were all higher than the null model (△AICc > 0; Appendix 6).

Stocks of necromass carbon and annual power dissipation index (PDIannual) in the high wind exposure (Lanjenchi) and low wind exposure (Nanjenshan) plots. a Total stock; b fallen stock; c standing stock (transect-length-weighted plot necromass average ± SE).
Quantity of Necromass Input
The quantity and variations of woody debris input showed that total necromass input (Figure 3a) was mainly contributed by fallen woody debris (Figure 3b). The quantity of standing necromass input is low (Figure 3c), and contributes less to the ‘total’ necromass input (Figure 3a), as with the patterns of stocks at the previous section.

Inputs of necromass carbon and seasonal power dissipation index (PDIseasonal) in the high wind exposure (Lanjenchi) and low wind exposure (Nanjenshan) plots. a Total input; b fallen input; c standing input (transect-length-weighted plot average ± SE). Typhoon seasons are indicated in light grey, monsoon seasons in light blue. Red arrows indicate tracks of typhoons with PDI ≥ 0.1 (109 m3 s−2) based on the 20-km distant Hengchun weather station and include Typhoons Tembin (No. 201214), Usagi (No. 201319), Fung-Wong (No. 201416), Soudelor (No. 201513), Goni (No. 201515), Meranti (No. 201614), and Megi (No. 201617).
The number of typhoons varied between years (Figure 3), with high PDI corresponding to high inputs of total, fallen, and standing woody debris (Figure 4). As necromass input was not normally distributed (Shapiro–Wilk normality test, p < 0.001), we log10-transformed the data to have an approximately normal distribution before applying the following tests. Total, fallen and standing necromass input differ significantly between seasons (Appendix 7). The results of the linear mixed-effect model indicated that PDIseasonal was the best climatic valuables for explaining the seasonal variation in the total, fallen and standing necromass input (Table 2). The best models (△AICc = 0) were those included both plot and PDIseasonal (Table 2). Including the interaction variables (plot and the best climatic variable) improved the fallen input and the standing input models (Table 2). Notably, the coefficients of the plot variable of the best models were opposite for the fallen and the standing necromass input models (Table 3).
Necromass Decomposition and Net Fluxes
The decay rate constant of the study area ranged from 0.57 to 1.09 y−1 (Appendix 8). The fast decomposition of IWD and FWD in the low wind exposure plot resulted in the half-life of woody debris smaller than 10 cm being less than one year. We found that the annual necromass input and decomposition fluxes fluctuated annually (Figure 5a). Also, the net flux fluctuated around 0 (Figure 5b). On average, net flux was 0.40 ± 0.43 (Mg C ha−1 y−1) in the high wind exposure plot and 0.08 ± 0.69 (Mg C ha−1 y−1) in the low wind exposure plot during the study period (Figure 5b; Appendix 4).

Necromass input and seasonal power dissipation index (PDIseasonal) in the high wind exposure (Lanjenchi) and the low wind exposure (Nanjenshan) plots. Each point represents transect-length-weighted average of each plot.
Discussion
Necromass Stock and Input at the Global Scale
Our study forests are located on a tropical island that experiences strong disturbances from both typhoon and monsoon winds (Wang 2004; Tu and others 2009). Remarkably, the total quantity of necromass carbon stock in both plots was very low compared to other tropical forests (Figure 6a). Lutz and others (2018) compared many temperate and tropical forests and found that our high wind exposure plot (Lanjenchi) was ranked as one of the smallest biomass forests globally. Chao and others (2009a) proposed that there is a relationship between aboveground biomass and necromass stock in Amazonian forests. Our study supports this relationship by using more data (Figure 6a). It is now clear that necromass tracks tropical forest biomass stocks over a very broad geographical range (Figure 6a).

Fluxes and net values of necromass carbon in the high wind exposure (Lanjenchi) and low wind exposure (Nanjenshan) plots during the study period. Input fluxes include both fallen and standing necromass of carbon (NCI); output fluxes are the decomposition quantity (NCD); the net flux is the difference between input and output fluxes.

Necromass carbon stock in tropical primary forests and its relationships with a aboveground biomass carbon stock and b necromass carbon input. (Data extracted from Baker and others 2007; Chao and others 2008; Palace and others 2008; Chao and others 2009a; Gurdak and others 2014; Sato and others 2016; Gora and others 2019). Values are recalculated assuming 50% of necromass is carbon because the original reports did not measure sample carbon concentrations.
The necromass input measured in our study plots is also low relative to other studies in mature tropical forests (Figure 6b). Notably, the differences in necromass inputs between our study forests and other studies (up to half) (Figure 6b) are less marked than the differences in necromass stocks (up to one-tenth less) (Figure 6a). Lin and others (2020) noted that the major consequences of typhoons on forest ecosystems in Taiwan are typically defoliation, rather than tree death. Our study further shows that at the global scale, the input of branch fall and tree fall in highly wind-disturbed forests is low compared to other tropical forests (Figure 6). The low necromass inputs and stocks are likely to be mediated by their low biomass.
The causal reasons for low biomass in the study forests can be attributed to the dwarfing effect of typhoons (as hypothesised by Lin and others (2020)), of monsoon winds (as hypothesised by Chao and others (2010b)), or both. Our study provides evidence that both wind regimes shape the structure of these forests (see Discussion in ‘Necromass Input at the Landscape Scale: Short Term’). Across the tropics, we expect that due to the long-term interaction of forests with wind disturbances, other localities with frequent typhoon and monsoon winds will also have low necromass stocks and unusual forest structure. These forests are expected to develop a wind-resistant physiognomy that includes many slim and short stems, which generate relatively small quantities of woody necromass (Figure 7a).
Necromass Stock and Input at the Landscape Scale: Long Term
At the landscape scale, despite the significant structural differences in stem density and biomass (Table 1), we detected no significant difference in total necromass stock between the study plots when controlling for the differences of transects (Appendix 6). Moreover, total necromass carbon stock quantities were not readily explained by any of the tested variables (Appendix 6). These results do not support our prediction 1.1 and suggest that at the landscape scale, total necromass stocks are unrelated to the effects of the regional prevailing wind, when accounting for transect variations (Appendix 6). This indicates that the differences are due to specific conditions of some transects being more likely to accumulate higher necromass than others. Thus, at the landscape scale, there is no evidence of direct effects of wind magnitude on necromass stocks.
Necromass input, however, is greater for the high wind exposure plot than the low wind exposure plot, after accounting for the differences at each transect (Table 3). The coefficient of the plot variable for the total necromass input model was positive (Table 3), suggesting that the high wind exposure plot had greater total necromass input during our study period, which supports our prediction 2.1. Apart from disturbance exposure degree, differences in necromass input between sites could also be affected by (1) forest structure, (2) species composition, and (3) frequency and modal size of woody debris. We discuss these factors in turn. (1) Forest structure: forest biomass and height are lower in the high wind exposure plot than the low wind exposure plot (Table 1), but necromass input was no lower, so the difference in necromass input cannot be explained by forest structure at the landscape scale. (2) Species composition: the two forests have distinct floristic composition from one another (Chao and others 2010b), so this might be causally related to the dissimilarity of necromass input at the landscape scale, potentially via differences in monsoon wind regimes favouring different species. (3) Frequency and modal size of woody debris: examining the patterns in woody debris size, we found that the quantities of fine and intermediate standing woody debris were quite high in the high wind exposure plot (Appendix 3b). Thus, a likely explanation is that the accumulation of fine and intermediate standing woody debris in the high wind exposure forest results in a total necromass input comparable to the quantity produced in the high biomass, low wind exposure forest. In other words, the patterns of necromass input are ‘small but many’ in the high wind exposure plot, and ‘large but few’ in the low wind exposure plot.
The fallen necromass input was lower and the standing necromass input was greater in the high wind exposure than the low wind exposure plot (Table 3). This is not what we expected (prediction 2.2). According to the literature, the major causal reasons for trees dying standing are senescence, competition, drought, fire, or large-scale pathogen attacks (for example, Carey and others 1994; Nakagawa and others 2000; Chao and others 2009b). Here there were no records of drought or fire, nor were there large-scale pathogen attacks during the study period. Based on our field observations, we found that the majority of the standing dead woody debris have signs of wind breakages and many had re-sprouting stems. This natural coppicing process is consistent with elsewhere in the tropics (Zimmerman and others 1994) and indicates that the major survival strategy of trees growing in wind-influenced forests is re-sprouting. Thus, we conclude that standing necromass does not necessarily indicate trees or stems dying due to competition or senescence, but can also arise indirectly from the strategy adopted by many trees to survive wind stress by growing multiple stems. In short, at the landscape scale, the relatively high necromass input in the high wind exposure forest, despite its low stature, was likely caused by direct wind effects, indirect wind effects through species composition, and the multiple small dead stems of coppicing trees (Figure 7b).
Necromass Input at the Landscape Scale: Short Term
There were significant seasonal variations in wind magnitude (Figure 1b) and fallen necromass input (Appendix 7b) during our short-term study period, with greater quantities and greater variation typically in typhoon and monsoon seasons. Although we refer to the main typhoon season as mid-July to mid-October, typhoons can occur at any month of the year. For example, the typhoon Noul (No. 201506) was recorded in May but with very low PDI (0.01 109 m3 s−2) (data not shown). However, in general, strong wind mostly occurred before the October and February census (the main typhoon and monsoon seasons) during our study period (Figure 3). There was also among-site variation: at times the low wind exposure plot had greater fallen necromass input (February 2014), whereas at others the high wind exposure plot experienced more (January 2017) (Figure 3b). This demonstrates that there are high variations both in wind patterns and necromass input in the study forests, especially in the monsoon season.
Among the seasonal climatic variables, PDI was the best for predicting necromass input when controlling for differences between transects (Table 2). Compared to PDIseasonal, neither precipitation, nor PDItyphoon, nor the number of typhoons was a good predictor (Table 2). Typhoon metrics alone (PDItyphoon or number of typhoons) cannot reflect the major climatic patterns on necromass input. However, PDIseasonal includes not only the magnitude of typhoons but also monsoon winds, thus reflecting the strength of winds from both processes. Moreover, the best model all includes PDIseasonal, suggesting that despite any other possible inherent differences between plots (for example, Table 1), PDIseasonal was a crucial driving force of necromass input (Table 2). Moreover, including the interaction terms improved the linear mixed-effect models of both the fallen and standing models, demonstrating that there were different responses of plots to PDIseasonal. Table 3 shows that large PDIseasonal may increase the quantities of fallen but not standing woody debris in the high wind exposure plot (negative coefficient for the interaction term of the standing woody debris Model in Table 3; see also Figure 4). This suggests that even for a forest ecosystem frequently influenced by winds, increased wind strength can still increase the likelihood of fallen woody debris, but not standing woody debris.
The effects of typhoons on forest ecosystems have drawn considerable research attention (Lugo 2008; Lin and others 2010), but those of monsoon winds much less so (Yu and others 2014). In our study, the month with the highest quantity of necromass input was not October (right after the peak season of typhoons) but was usually January or February, especially for fallen woody debris (Figure 3b). This shows that monsoon winds can have at least as strong influence on forest carbon dynamics as typhoons. This could be due to two reasons. First, the effects of each typhoon at one location normally last fewer than three days, whereas the period of the northeast monsoon can last for more than three months (from mid-October to mid-February). In other words, typhoons bring winds for a short period, whereas monsoon winds are relatively long-lasting. Thus, even though a single typhoon may have a strong daily PDI, the cumulative seasonal PDI was stronger during monsoon seasons than during typhoon seasons (Figure 3).
Second, the effects of typhoons on necromass input may be delayed. Based on our observations, some trees did not die immediately after being uprooted or broken by typhoons. It was common to find some fallen trunks with new sprouts after the typhoon season and these may last for some time. A similar phenomenon was also observed in another tropical cyclone disturbed forest (Uriarte and others 2019). However, the long-lasting monsoon wind could then have further weakened the vitality of the fallen but still living trees, resulting in peak necromass input in the February census (Figure 3). Also, Figure 3 reveals that in some years with weak typhoon effects on the study region (for example, September 2014 (Typhoon Feng-Wong) and August 2015 (Typhoons Soudelor and Goni)), even though the winter still has strong monsoon winds, the quantity of necromass input is low. Thus, in years where the strength of typhoon winds was insufficient to affect trees (based on our evaluated variables), the long-lasting monsoon winds did not result in substantial necromass inputs. However, in years with strong typhoon magnitude (for example, September 2013 (Usagi) and September 2016 (Meranti and Megi)), the input of necromass was high (Figure 3). An exception was recorded in February 2018 for which the previous typhoon season (October 2017) have a low PDIseasonal, but still had high necromass inputs (Figure 3).
In sum, our results suggest that at a short-term and landscape scale, the main driving factor of necromass carbon balance is typhoons, but that monsoons are a contributing and aggravating factor. Fallen necromass mainly result from the combination of the two climatic events, such that typhoons cause initial stem breakage and monsoon winds subsequently weaken tree vigour (Figure 7c). Therefore, both typhoons and monsoons need to be accounting for when modelling short-term forest dynamics.

Schematic synthesis of the effects of typhoon and monsoon winds on tropical forest ecosystems: a at the global scale; b at the landscape scale over the long term; c at the landscape scale over the short term.
Net Fluxes and Implication of the Future Trend
The decay rate constants of other tropical trees reported in the literature range from 0.015 to 0.67 (y−1) for fresh woody debris ≥ 10 cm (Chambers and others 2000; Baker and others 2007). Thus, the decay rate constants of our high wind exposure and low wind exposure plots are relatively high (Appendix 8). A global-scale study of woody debris decomposition has suggested that subtropical forests may be particularly influenced by the activities of soil macrofauna and with special decomposition pathways (Martin and others 2021), which could be the reason for the high decomposition rate in our study plots.
Although the necromass net fluxes were weakly positive in most years (Figure 5), the standard errors were large relative to the estimate values (Appendix 4), reflecting large year-to-year variation. Besides, the stocks of necromass were relatively low compared to other forests (Figure 6a), so any accumulation of necromass over the past few years was small. This suggests that the ecosystem both before and during our study period has been close to a dynamic equilibrium status.
Changes in climatic patterns could affect necromass dynamics in our forests. There is evidence of a long-term decline in monsoon wind intensity (Xu and others 2006), and there are suggestions that such a trend may continue in the future associated with warming winters (Xu and others 2006). For typhoons, the northwest tropical Pacific has experienced a historical strengthening in intensity but a decrease in frequency and duration (Tu and others 2009; Lin and Chan 2015). Some climate modelling suggests that both these trends will continue as the planet warms (Jiang and Tian 2013; Mei and Xie 2016). Although such changes are likely to affect these forests, it remains challenging to predict precisely how. As the forest structure and composition of our study forests are strongly influenced by monsoon winds (Chao and others 2010b), any decline in the intensity of monsoon wind could result in greater biomass growth after release from its stress (monsoon). In the meantime, since we see that PDI significantly influences the seasonal pattern of fallen and standing inputs (Table 2), we can expect a shift in the seasonality of necromass production, with more intense typhoons generating more woody debris during typhoon seasons but weakening monsoons contributing less woody debris. Consequently, the net balances of biomass and necromass stocks are likely to shift over short time scales at least.
Conclusion
At the global scale, necromass stocks and inputs were exceptionally low in forests influenced by both typhoon and monsoon winds. At the landscape scale, the magnitude of winds helps to explain the seasonal patterns of necromass input, and our analysis points to typhoons as being the primary cause and monsoon winds as aggravating factors in the production of necromass inputs. Therefore, both monsoons and typhoons need to be accounted for when modelling the dynamics of forest carbon balance in tropical and subtropical forests away from the equatorial belt. Our study also demonstrates how tropical trees adapt to windy environments, with reduced stature and the ability to resprout contributing to ecosystem resistance and resilience. Changes in wind intensity and duration need greater attention from climatologists and ecologists as they are likely to drive changes in forest structure, carbon balances and dynamics this century.
Data Availability
References
Baker TR, Honorio Coronado EN, Phillips OL, Martin J, van der Heijden GMF, Garcia M, Silva Espejo J. 2007. Low stocks of coarse woody debris in a southwest Amazonian forest. Oecologia 152:495–504.
Berbeco MR, Melillo JM, Orians CM. 2012. Soil warming accelerates decomposition of fine woody debris. Plant and Soil 356:405–417.
Brienen RJW, Phillips OL, Feldpausch TR, Gloor E, Baker TR, Lloyd J, Lopez-Gonzalez G, Monteagudo-Mendoza A, Malhi Y, Lewis SL, Vásquez Martinez R, Alexiades M, Álvarez Dávila E, Alvarez-Loayza P, Andrade A, Aragão LEOC, Araujo-Murakami A, Arets EJMM, Arroyo L, C. GAA, Bánki OS, Baraloto C, Barroso J, Bonal D, Boot RGA, Camargo JLC, Castilho CV, Chama V, Chao KJ, Chave J, Comiskey JA, Cornejo Valverde F, da Costa L, de Oliveira EA, Di Fiore A, Erwin TL, Fauset S, Forsthofer M, Galbraith DR, Grahame ES, Groot N, Hérault B, Higuchi N, Honorio Coronado EN, Keeling H, Killeen TJ, Laurance WF, Laurance S, Licona J, Magnussen WE, Marimon BS, Marimon-Junior BH, Mendoza C, Neill DA, Nogueira EM, Núñez P, Pallqui Camacho NC, Parada A, Pardo-Molina G, Peacock J, Peña-Claros M, Pickavance GC, Pitman NCA, Poorter L, Prieto A, Quesada CA, Ramírez F, Ramírez-Angulo H, Restrepo Z, Roopsind A, Rudas A, Salomão RP, Schwarz M, Silva N, Silva-Espejo JE, Silveira M, Stropp J, Talbot J, ter Steege H, Teran-Aguilar J, Terborgh J, Thomas-Caesar R, Toledo M, Torello-Raventos M, Umetsu RK, van der Heijden GMF, van der Hout P, Guimarães Vieira IC, Vieira SA, Vilanova E, Vos VA, Zagt RJ. . 2015. Long-term decline of the Amazon carbon sink. Nature 519:344–348.
Burnham KP, Anderson DR. 2002. Model selection and multimodel inference: a practical information-theoretic approach. New York: Springer-Verlag.
Carey EV, Brown S, Gillespie AJR, Lugo AE. 1994. Tree mortality in mature lowland tropical moist and tropical lower montane moist forests of Venezuela. Biotropica 26:255–265.
Central Weather Bureau. 2019. http://rdc28.cwb.gov.tw/data.php.
Chambers JQ, Higuchi N, Schimel JP, Ferreira LV, Melack JM. 2000. Decomposition and carbon cycling of dead trees in tropical forests of the central Amazon. Oecologia 122:380–388.
Chao K-J, Chao W-C, Chen K-M, Hsieh C-F. 2010a. Vegetation dynamics of a lowland rainforest at the northern border of the Paleotropics at Nanjenshan, southern Taiwan. Taiwan J Forest Sci 25:29–40.
Chao K-J, Chen Y-S, Song G-ZM, Chang Y-M, Sheue C-R, Phillips OL, Hsieh C-F. 2017. Carbon concentration declines with decay class in tropical forest woody debris. Forest Ecol Manag 391:75–85.
Chao K-J, Phillips OL, Baker TR. 2008. Wood density and stocks of coarse woody debris in a northwestern Amazonian landscape. Canadian J Forest Res 38:795–825.
Chao K-J, Phillips OL, Baker TR, Peacock J, Lopez-Gonzalez G, Martínez RV, Monteagudo A, Torres-Lezama A. 2009a. After trees die: quantities and determinants of necromass across Amazonia. Biogeosciences 6:1615–1626.
Chao K-J, Phillips OL, Monteagudo A, Torres-Lezama A, Vásquez Martínez R. 2009b. How do trees die? Mode of death in northern Amazonia. J Veg Sci 20:260–268.
Chao W-C, Song G-Z, Chao K-J, Liao C-C, Fan S-W, Wu S-H, Hsieh T-H, Sun I-F, Kuo Y-L, Hsieh C-F. 2010b. Lowland rainforests in southern Taiwan and Lanyu, at the northern border of paleotropics and under the influence of monsoon wind. Plant Ecol 210:1–17.
Chave J, Réjou-Méchain M, Búrquez A, Chidumayo E, Colgan MS, Delitti WBC and other 2014. Improved allometric models to estimate the aboveground biomass of tropical trees. Global Change Biol 20:3177–3190.
Clark DA, Asao S, Fisher R, Reed S, Reich PB, Ryan MG, Wood TE, Yang XJ. 2017. Reviews and syntheses: Field data to benchmark the carbon cycle models for tropical forests. Biogeosciences 14:4663–4690.
Douglas B, Martin M, Bolker B, Walker S. 2015. Fitting linear mixed-effects models using lme4. J Stat Software 67:1–48.
Emanuel K. 2005. Increasing destructiveness of tropical cyclones over the past 30 years. Nature 436:686–688.
Esquivel-Muelbert A, Phillips OL, Brienen RJW, Fauset S, Sullivan MJP, Baker TR and others. Tree mode of death and mortality risk factors across Amazon forests. Nature Commun 11:5515.
Franklin JF, Shugart HH, Harmon ME. 1987. Tree death as an ecological process. Bioscience 37:550–556.
Gora EM, Kneale RC, Larjavaara M, Muller-Landau HC. 2019. Dead wood necromass in a moist tropical forest: stocks, fluxes, and spatiotemporal variability. Ecosystems 22:1189–1205.
Gurdak DJ, Aragao LEOC, Rozas-Davila A, Huasco WH, Cabrera KG and others 2014. Assessing above-ground woody debris dynamics along a gradient of elevation in Amazonian cloud forests in Peru: balancing above-ground inputs and respiration outputs. Plant Ecol Divers 7:143–160.
Harmon ME, Sexton J. 1996. Guidelines for measurements of woody detritus in forest ecosystems. Seattle, Washington: US Long Term Ecological Research Network Office Publication.
IPCC. 2006. Forest lands. 2006 Intergovernmental Panel on Climate Change Guidelines for National Greenhouse Gas Inventories. Vol 4: Agriculture, Forestry, and Other Land Use. Hayama, Japan on behalf of the IPCC: Institute for Global Environmental Strategies (IGES), p83.
Jiang DB, Tian ZP. 2013. East Asian monsoon change for the 21st century: results of CMIP3 and CMIP5 models. Chinese Sci Bull 58:1427–1435.
Keller M, Palace M, Asner GP, Pereira R, Silva JNM. 2004. Coarse woody debris in undisturbed and logged forests in the eastern Brazilian Amazon. Global Change Biol 10:784–795.
Köhl M, Lasco R, Cifuentes M, Jonsson Ö, Korhonen KT, Mundhenk P, Navar JdJ, Stinson G. 2015. Changes in forest production, biomass and carbon: Results from the 2015 UN FAO global forest resource assessment. Forest Ecol Manag 352:21–34.
Lawton RO. 1982. Wind stress and elfin stature in a montane rain forest tree: an adaptive explanation. Am J Bot 69:1224–1230.
Li C-F, Chytrý M, Zelený D, Chen M-Y, Chen T-Y and others 2013. Classification of Taiwan forest vegetation. Appl Veg Sci 16:698–719.
Liao P-S. 2017. Carbon stocks and fluxes of Nanjenshan Tropical Forests, Taiwan. Depart of Life Sciences. Taichung: National Chung Hsing University.
Lin I-I, Chan JCL. 2015. Recent decrease in typhoon destructive potential and global warming implications. Nature Commun 6:7182.
Lin T-C, Hamburg SP, Lin K-C, Wang L-J, Chang CT, Hsia Y-J, Vadeboncoeur MA, McMullen CMM, Liu C-P. 2010. Typhoon disturbance and forest dynamics: lessons from a northwest Pacific subtropical forest. Ecosystems 14:127–143.
Lin T-C, Hogan JA, Chang C-T. 2020. Tropical Cyclone Ecology: A Scale-Link Perspective. Trends in Ecology & Evolution.
Lugo AE. 2008. Visible and invisible effects of hurricanes on forest ecosystems: an international review. Austral Ecol 33:368–398.
Lutz JA, Furniss TJ, Johnson DJ, Davies SJ, Allen D, Alonso A, Anderson-Teixeira KJ and others 2018. Global importance of large-diameter trees. Global Ecology and Biogeography: 1–16.
Martin AR, Domke GM, Doraisami M, Thomas SC. 2021. Carbon fractions in the world’s dead wood. Nat Commun 12:889.
McDowell N, Allen CD, Anderson-Teixeira Kand others 2018. Drivers and mechanisms of tree mortality in moist tropical forests. New Phytol 219:851–869.
Mei W, Xie SP. 2016. Intensification of landfalling typhoons over the northwest Pacific since the late 1970s. Nature Geosci 9:753–757.
Nakagawa M, Tanaka K, Nakashizuka T and others 2000. Impact of severe drought associated with the 1997–1998 EL Niño in a tropical forest in Sarawak. J Trop Ecol 16:355–367.
Olson JS. 1963. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 44:322–331.
Palace M, Keller M, Silva H. 2008. Necromass production: studies in undisturbed and logged Amazon forests. Ecol Appl 18:873–884.
Palace M, Keller M, Hurtt G, Frolking S. 2012. A Review of Above Ground Necromass in Tropical Forests. Sudarchana P editor. Tropical Forests: InTech.
Pan YD, Birdsey RA, Fang JY, Houghton R, Kauppi PE, Kurz WA, Phillips OL, Shvidenko A, Lewis SL, Canadell JG, Ciais P, Jackson RB, Pacala SW, McGuire AD, Piao SL, Rautiainen A, Sitch S, Hayes D. 2011. A large and persistent carbon sink in the world’s forests. Science 333:988–993.
Phillip MS. 1994. Measuring trees and forests. Wallingford, U.K.: CAB International.
Pietsch KA, Eichenberg D, Nadrowski K, Bauhus J, Buscot F, Purahong W, Wipfler B, Wubet T, Yu MJ, Wirth C. 2019. Wood decomposition is more strongly controlled by temperature than by tree species and decomposer diversity in highly species rich subtropical forests. Oikos 128:701–715.
Quéré CL, Andrew RM, Friedlingstein P, Sitch S, Pongratz J, Manning AC. 2018. Global carbon budget 2017. Earth Syst Sci Data Discuss 10:405–448.
R Core Team. 2019. R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing.
Sato T, Yagihashi T, Niiyama K, Abd Rahman K, Azizi R. 2016. Coarse woody debris stocks and inputs in a primary hill dipterocarp forest, peninsular Malaysia. J Trop Forest Sci 28:382–391.
Soethe N, Lehmann J, Engels C. 2006. Root morphology and anchorage of six native tree species from a tropical montane forest and an elfin forest in Ecuador. Plant and Soil 279:173–185.
Sullivan MJP, Lewis SL, Affum-Baffoe K, Castilho C, Costa F, Sanchez AC, Ewango CEN, Hubau W and others 2020. Long-term thermal sensitivity of Earth’s tropical forests. Science 368:869.
Trumbore S. 2006. Carbon respired by terrestrial ecosystems - recent progress and challenges. Global Change Biol 12:141–153.
Tu JY, Chou C, Chu PS. 2009. The abrupt shift of typhoon activity in the vicinity of Taiwan and its association with western North Pacific-East Asian climate change. Journal of Climate 22:3617–3628.
Uriarte M, Thompson J, Zimmerman JK. 2019. Hurricane María tripled stem breaks and doubled tree mortality relative to other major storms. Nature Communications 10.
van der Meer PJ, Bongers F. 1996. Patterns of tree-fall and branch-fall in a tropical rain forest in French Guiana. J Ecol 84:19–29.
van Wagner CE. 1968. The line intersect method in forest fuel sampling. Forest Sci 24:469–483.
Wang C. 2004. Features of monsoon, typhoon and sea waves in the Taiwan Strait. Marine Georesources & Geotechnology 22:133–150.
Whigham DF, Olmsted I, Cano EC, Harmon ME. 1991. The impact of Hurricane Gilbert on trees, litterfall, and woody debris in a dry tropical forest in the Northeastern Yucatan Peninsula. Biotropica 23:434–441.
Xu M, Chang C-P, Fu C, Qi Y, Robock A, Robinson D, Zhang H-m. 2006. Steady decline of east Asian monsoon winds, 1969–2000: Evidence from direct ground measurements of wind speed. Journal of Geophysical Research: Atmospheres 111.
Yu GR, Chen Z, Piao SL, Peng CH, Ciais P, Wang QF, Li XR, Zhu XJ. 2014. High carbon dioxide uptake by subtropical forest ecosystems in the East Asian monsoon region. Proceed Nat Acad Sci United States of Am 111:4910–4915.
Yu J-Y, Chiu P-G. 2012. Constrasting various metrics for measuring tropical cyclone activity. Terr Atmos Ocean Sci 23:303–316.
Zimmerman JK, Everham EM III, Waide RB, Lodge DJ, Taylor CM, Brokaw NVL. 1994. Responses of tree species to hurricane winds in subtropical wet forest in Puerto Rico: implications for tropical tree life histories. J Ecol 82:911–922.
Acknowledgements
We sincerely appreciate the important fieldwork done by Yi-Ju Li, Yen-Chen Chao, Chia-Min Lin, Chien-Hui Liao, Yeh Hsu, Chun-Yao Liu and numerous student volunteers. We thank Chia-Cheng Yang, Wei-Hong Chan, Dr. Wei-Chun Chao, Dr. I-Fang Sun, Dr. Tsung-Hsin Hsieh and Dr. Chang-Fu Hsieh for their pioneer works in the study forests. We are also grateful to Dr. Tsung-I Lin for his statistical consultation and to Kenting National Park for its logistic support. This study was funded by grants to Kuo-Jung Chao from the Ministry of Science and Technology, Taiwan (NSC 101-2313-B-005-024-MY3 and MOST 104-2313-B-005-032-MY3).
Author information
Authors and Affiliations
Corresponding author
Additional information
Author Contributions
KJC and GZMS designed the study. KJC, PSL and YSC carried out the analysis with inputs from GZMS, OLP and HJL. KJC, PSL and YSC wrote the manuscript with inputs from GZMS, OLP and HJL. PSL and YSC coordinated data collection with the help of KJC, GZMS and HJL. All co-authors commented on the manuscript.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Chao, KJ., Liao, PS., Chen, YS. et al. Very Low Stocks and Inputs of Necromass in Wind-affected Tropical Forests. Ecosystems 25, 488–503 (2022). https://doi.org/10.1007/s10021-021-00667-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-021-00667-z