
Abstract
Soil respiration (SR) is a major component of the global carbon cycle and plays a fundamental role in ecosystem feedback to climate change. Empirical modelling is an essential tool for predicting ecosystem responses to environmental change, and also provides important data for calibrating and corroborating process-based models. In this study, we evaluated the performance of three empirical temperature–SR response functions (exponential, Lloyd–Taylor and Gaussian) at seven shrublands located within three climatic regions (Atlantic, Mediterranean and Continental) across Europe. We investigated the performance of SR models by including the interaction between soil moisture and soil temperature. We found that the best fit for the temperature functions depended on the site-specific climatic conditions. Including soil moisture, we identified thresholds in the three different response functions that improved the model fit in all cases. The direct soil moisture effect on SR, however, was weak at the annual time scale. We conclude that the exponential soil temperature function may only be a good predictor for SR in a narrow temperature range, and that extrapolating predictions for future climate based on this function should be treated with caution as modelled outputs may underestimate SR. The addition of soil moisture thresholds improved the model fit at all sites, but had a far greater ecological significance in the wet Atlantic shrubland where a fundamental change in the soil CO2 efflux would likely have an impact on the whole carbon budget.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Soil respiration (SR) is a dominant component of the terrestrial carbon cycle and has a significant influence on global radiative forcing (IPCC 2013). In terrestrial ecosystems, atmospheric CO2 is assimilated during photosynthesis, and then released either via autotrophic respiration or through heterotrophic decomposition of carbon compounds differing in recalcitrance and sensitivity to temperature (Davidson and Janssens 2006). Both soil moisture availability and temperature may alter with a changing climate, and this will affect decomposition processes and root activity, potentially changing rates of CO2 efflux from soils. However, it is poorly understood how altered temperature and soil moisture availability will affect soil CO2 efflux across multiple ecosystems. In fact, because of the interaction of multiple environmental processes often occurring simultaneously it is hard to make predictions beyond empirical data bounds (Vicca and others 2014). SR response functions derived from empirical data collected at different temporal and spatial scales could be useful for improving the predicted impact of future climate on ecosystem processes (Kirschbaum 2004; Vicca and others 2014).
Temperature is often a predominant factor controlling biological metabolic processes and a broad spectrum of relationships between temperature and SR has been tested (Subke and Bahn 2010; Wu and others 2011; Shen and others 2013). Most commonly, the exponential function has been used to model the temperature–respiration relationship (Davidson and Janssens 2006; Beier and others 2009; Vicca and others 2014). In these cases, however, exponential models were usually applied in a relatively narrow temperature range not exceeding 30°C. In situ SR studies covering a wide range of temperature and moisture conditions are rare and the limited availability of such data affects the ability of modellers to fit SR functions to empirical data (Vicca and others 2014). Consequently, to study SR on a wide range of ecosystems and climatic conditions, the Arrhenius, Lloyd–Taylor, Gaussian, and Quadratic functions have been used (Lloyd and Taylor 1994; Tuomi and others 2008; Reichstein and Beer 2008; Lellei-Kovács and others 2011; González-Ubierna and others 2014).
Occasionally, to improve on the fit of a simple exponential model, a wider environmental range has been incorporated by fitting separate functions to subranges of temperature (Murthy and others 2003; Bradford and others 2008) and soil depth (Pavelka and others 2007) or to Mediterranean wet versus dry seasons (de Dato and others 2010). Other studies have used additional parameters to account for factors other than temperature like soil moisture content (Suseela and others 2012; Kopittke and others 2013; Wang and others 2014), soil physical and chemical properties (Wang and others 2003; Balogh and others 2011; Kotroczó and others 2014), different substrate availability (Davidson and others 2006), or different SOM content and quality (Curiel Yuste and others 2010). Other studies have also attempted to provide mechanistic explanations for the temperature dependence of SR (Davidson and others 2006; von Lützow and Kögel-Knabner 2009). In a review, Billings and Ballantyne (2013) examined the mechanisms that are linked to SR, and reported that temperature-induced changes in microbial community structure, microbial metabolic rates and catalytic rate of exo-enzymes may lead to a decline of SR as a response to an increase in the soil temperature.
The relationship between soil moisture and SR has been modelled using many different functions that include linear (Leirós and others 1999), exponential (Rodrigo and others 1997), second-order exponential, that is, Gaussian (Howard and Howard 1993; Mielnick and Dugas 2000; Vicca and others 2014) and reverse exponential (Zhou and others 2007) relationships. Limitation of SR by soil moisture has been observed when substrate diffusion is limited by low soil water availability (Howard and Howard 1993), but also when the diffusion of O2 is restricted by high soil water content (Skopp and others 1990). Mechanistic studies of the relationship between soil moisture and SR conducted by Davidson and others (2006) revealed that not only CO2 efflux is influenced by moisture-induced changes in soil physical properties, but also autotrophic root respiration and heterotrophic microbial decomposition are directly impacted by changes in soil moisture. Evaluation of the impact of soil moisture is more difficult than that of temperature because the efficiency of water uptake is influenced by various soil physical properties and also by physiological processes of the organisms. At any given soil moisture content, water uptake may differ for numerous reasons such as soil texture (sand or clay), plant water use efficiency, stress tolerance and soil microbial composition (for example, fungal to bacterial ratio) (Moyano and others 2013).
The approaches to studies that combined impact of temperature and moisture on SR modelling differ in two fundamental ways: (1) Additive versus interactive (Mielnick and Dugas 2000; Reichstein and others 2002; Qi and others 2002; Xu and others 2004; Zhou and others 2006); (2) Continuous versus threshold (Davidson and others 1998; Reichstein and others 2002; Rey and others 2002; Fernandez and others 2006; Yan and others 2011). Moisture thresholds that alter SR activities significantly may be very important in modelling carbon fluxes, not only in arid and semiarid, but also in mesic ecosystems (Suseela and others 2012).
In a coordinated network of climate change experiments (EU projects CLIMOOR, VULCAN and INCREASE), along a natural temperature and precipitation gradient across European shrublands, whole ecosystem manipulations of warming and summer drought conditions were conducted. The experiments resulted in a trend of increased SR in response to the warming treatments and significant reduction in SR in response to the drought treatments (Emmett and others 2004; Kopittke and others 2014). However, some of the ecosystems also had an individual response to warming and drought that makes general conclusions difficult to draw. In the longer term, repeated summer drought resulted in an increased SR in the hydric ericaceous shrubland in Wales. Sowerby and others (2008) suggested that the year-round reduction in soil moisture content of the organic-rich podzol soil resulted in a year-round stimulation of SR. Lellei-Kovács and others (2008) found that in the semiarid Hungarian shrubland, warming and drought reduced the rate of SR. In the Italian Mediterranean shrubland, de Dato and others (2010) observed a temporary decrease in SR as a short-term response to the warming and drought treatments.
In a previous study, we investigated the mechanisms that control SR in the semiarid Hungarian shrubland with extreme temperature and soil moisture regimes, by empirically modelling SR as a response function of temperature and moisture (Lellei-Kovács and others 2011). Applying the same approaches, here we expand this work by modelling SR using 2- or 3-years of empirical data collected from seven different shrubland ecosystems across Europe with markedly different natural temperature and moisture regimes. We compared the performance of three empirical SR models, the exponential, the Lloyd–Taylor and the Gaussian functions, and integrated moisture into the models using additive and interactive approaches. The aims were to (i) investigate the effect of soil temperature and soil moisture content on SR in the different soils, and (ii) improve model predictions of SR under future climate change scenarios. We hypothesized, that (i) the exponential model performs appropriately only in a relative narrow temperature range, (ii) the Gaussian temperature dependence function would be the best predictive SR model in ecosystems exposed to a relatively large temperature range, and (iii) inclusion of soil moisture thresholds would improve the predictive power of the models at sites in which moisture is an obvious controlling factor (for example, xeric or hydric ecosystems), whilst in mesic ecosystems the inclusion of moisture would have a smaller impact.
Materials and Methods
Characteristics of the Studied Shrubland Ecosystems
The study was conducted along natural temperature and precipitation gradients across Europe (Beier and others 2009), in seven different shrubland ecosystems (see Tables 1, 2), that included four Atlantic heathlands at two sites in Denmark (Mols, DK-M, and Brandbjerg, DK-B), one site in the Netherlands (Oldebroek, NL), and one site in the United Kingdom (Clocaenog, UK) (Sowerby and others 2008), two Mediterranean garrigues, one in Spain (Garraf, ES) (Sardans and others 2008) and one in Italy (Capo Caccia, IT) (de Dato and others 2010), and one shrubland in the Pannonian sandy forest steppe region in Hungary (Kiskunság, HU) (Lellei-Kovács and others 2011). Meteorological data between 2001 and 2012 (except ES between 2002 and 2003 and DK-B between 2006 and 2012) were recorded either directly at the sites, or at standard meteorological stations located nearby (Table 1). Mean annual temperature ranges from 8.0 at the DK-B site to 16.8 at the IT site. Mean annual precipitation varies between 549 mm in IT and 1345 mm at the UK site. The variability of climate among sites could be expressed by the modified Gaussen-index (mean annual precipitation/2× mean annual temperature, Peñuelas and others 2007) with higher aridity at its lower values (Table 1).
Field Experiments and Measurements
Plot-sized climate manipulation experiments were established in the seven shrubland ecosystems (see above). The experimental plots were subjected to either year-round passive night-time warming by insulating reflective curtains, extended drought periods by rain-activated transparent polyethylene roofs or an un-treated control since 1999 (ES, UK, NL, DK-M), since 2001 (HU, IT) or since 2005 (DK-B) (for detailed description of the experimental design and the effects on soil temperature and moisture, see at Beier and others 2004; Lellei-Kovács and others 2008; Mikkelsen and others 2008; de Dato and others 2010). In this study, we used data from different treatments together with data from control plots, that is, a response surface approach, where treatments are seen as a widening of the natural range of environmental variables (see also Lellei-Kovács and others 2011, and Table S1 for data of the treatment effects on soil temperature, soil moisture and SR).
Two or three years of SR measurements were conducted biweekly or monthly in the experimental plots, with exception of periods with snow cover and when the soil surface was frozen. Measurements were done between 2010 and 2012, but in ES between 2002 and 2003. SR data presented are the sum of autotrophic (root respiration) and heterotrophic (microbial respiration) soil processes. SR rates were measured by infrared gas exchange systems equipped with SR chambers: LI-6400XT with LI-6400-09 chamber (LICOR Biosciences, Lincoln, NE USA) in the NL and DK sites; LI-8100 with 8100-102 chamber (LICOR Biosciences, Lincoln, NE USA) in the UK and IT sites; EGM-3 (PP Systems, Hertfordshire, UK) in manual mode to analyse air samples from a closed-type, custom-built PVC chamber in ES; ADC Leaf Chamber Analyzer 4 with PLC and 2250 Soil hood (ADC BioScientific, Hoddesdon, UK) in HU. Three permanent subplots were used within each plot to capture within-plot heterogeneity, and plot means were used in the subsequent analyses. (For further details see: Beier and others 2009; de Dato and others 2010; Lellei-Kovács and others 2011; Kopittke and others 2013.) Micrometeorological variables were recorded in every plot continuously by automated instruments (Table 3): Soil temperature at 5 cm below the soil surface, and volumetric soil moisture content at the defined soil depths (Table 2).
Soil properties including soil texture (mechanical and Pipet Method), soil organic matter content (Tyurin method or dry combustion) and soil pH (by potentiometer with glass electrode) were measured at each site at the given soil depths (Table 2) before starting the treatments. Wilting point and field capacity were determined from the soil moisture retention curve (pF curve) using soil samples from the sites (Table 5) at the defined soil depths (Table 2). An exception was IT, where soil texture data were used to determine wilting point and field capacity (Saxton and Rawls 2006).
Empirical Model of the Temperature and Moisture Sensitivity of SR
For statistical evaluation, we followed the methodology used by Lellei-Kovács and others (2011) and treated the datasets of the seven sites independently. Separate analyses for each site were necessary to account for differences in biota, organic matter content, texture and moisture content (Tables 2, 3).
We first fit three different temperature dependence models (see equations 1–3). Each of the three response functions represents a possible relationship between increasing soil temperature and SR. Specifically: (i) the exponential function assumes that the logarithm of respiration is a linear function of temperature, thus the Q10 temperature coefficient is constant (equation 1); (ii) the Lloyd–Taylor function assumes that the influence of temperature change is higher at lower than at higher temperatures, thus the logarithm of respiration is a saturating function of temperature, and Q10 decreases with increasing temperature and its asymptote is one (that is, at extremely high temperature there is no further change in respiration) (equation 2); and (iii) the Gaussian function presumes that there is an optimal temperature for SR. Above this optimum, an increase in temperature causes a decline in SR. In this case Q10 is also a decreasing function of temperature, but it can fall below one (equation 3).
Equations (1)–(3). The models used to fit soil temperature and SR field data, where SR = soil respiration; T = soil temperature in Kelvin; a, b and c are parameters of the models:
After a log transformation of SR data, the exponential and the Gaussian functions (equations 1, 3) could be fit using linear regression. The Lloyd–Taylor function (equation 2) was fit by non-linear least squares regression also using log-transformed SR as a dependent variable to make the models statistically comparable, as discussed further below. To initialize the parameters of non-linear fit, parameter c was set to zero, whereas starting values of a and b were calculated by linear regression using 1/T as an independent variable.
In some cases, to preserve the expected shape of the fit curve, we had to apply constraints on the parameters of equations (1)–(3). These constraints for the functions were
The potential effect of soil moisture content on SR was analysed comparing three different soil moisture inclusion methods in the temperature dependence models:
-
1.
There is no inclusion of soil moisture content,
-
2.
The effects of soil moisture content and soil temperature are additive (that is, only parameter a depends on soil moisture content),
-
3.
The effects of soil moisture content and soil temperature are interactive.
Combining the three temperature dependence functions and the three soil moisture effects resulted in nine models for each site. We treated the soil moisture effect as a categorical variable as we did not have any a priori knowledge of its functional form. Additive effect means that soil moisture influences only the parameter a, thus, within one model, temperature dependence curves of logSR are parallel at different moisture levels, whereas interaction means that soil moisture influences parameters b and c too resulting in non-parallel temperature dependence curves of logSR.
In many cases, arbitrarily chosen cut-off points are used for transforming continuous variables into categories that introduce subjectivity into the modelling process. To avoid this problem, our categorizations were created by fitting decision tree models using a conditional inference framework that resulted in different soil moisture cut-off points depending on the applied temperature functions. When testing for additive effects, the residuals of the temperature functions were the dependent functions of the conditional inference trees (Hothorn and others 2006) that search for homogeneous groups of residuals (and thus parameter a) according to moisture values. We applied model-based recursive partitioning (Zeileis and others 2005) to search for categories in soil moisture that were homogeneous in the parameters of temperature dependence. Because model-based partitioning can handle linear models only, we assumed that the parameter c of the Lloyd–Taylor function was independent of soil moisture, and equal to the value estimated in the first approach (no soil moisture effect). Based on this assumption, we fit the Lloyd–Taylor function by linear regression using 1/(T − c) as independent variable.
To compare the performance of SR models with different numbers of parameters, we used corrected Akaike Information Criteria (AICc) that combine fit and complexity of models; smaller values indicate a better model (Johnson and Omland 2004). Because log-transformed SR values were used as dependent variables in all models, AICc values calculated for different models were comparable (Burnham and Anderson 2002). For statistical comparison of the models, we calculated the Akaike weights (Johnson and Omland 2004) of the models in two ways: (i) models that considered only soil temperature; (ii) all the nine models of the three temperature functions combined with the three ways of soil moisture inclusions. Akaike weights were calculated for each site and in each of aforementioned methods separately (see in Table 4). As the sum of Akaike weights calculated in one inter-comparison is 1, the model with an Akaike weight above 0.9 was considered unequivocally the best, and all the others were not interpreted. In case of more models having Akaike weights above 0.1, all these models were accepted with approximately a similar level of support in the data (Johnson and Omland 2004).
All statistical analyses were conducted in the R statistical environment (R Development Core Team 2008), tree models were fit using the party package (Hothorn and others 2006).
Calculations of Annual SR Rates by the Empirical Models of SR
Based on the soil temperature and moisture models of SR demonstrated above, we calculated the annual SR using the daily measured soil temperature and soil moisture meteorological data for years 2010, 2011 and 2012 in the control plots at all but the ES site. For the ES site year-round daily soil moisture data were not available for the calculations. We estimated the median and the 90% confidence interval of the estimated annual SR using the Monte Carlo simulation: predicted values were calculated with parameters randomly chosen from a multivariate normal distribution with means and co-variances estimated by fitting 10,000 times. Because of the collinearity of partial derivatives with respect to the parameter b and c in the Lloyd–Taylor model, these parameters were associated with large values in the variance–covariance matrix, leading to extremely wide confidence intervals. Because the wide confidence intervals were an artefact of the non-linear regression, in the case of the Lloyd–Taylor model, we decided to use only the predicted values.
Results
Variability of Environmental Factors and SR During the Study
Soil texture varied among sites, with high sand content at HU, NL, DK-M and DK-B, high silt content at ES and UK, and relatively high clay content at the Mediterranean ES and IT sites. Soil pH was alkaline at HU, ES and IT, whereas it was acidic at the Atlantic UK, NL, DK-M and DK-B sites. Soil organic carbon content, the main substrate for SR, was highly variable among sites (Table 2).
Soil temperature, moisture and SR all differed markedly among the different sites and over the studied period (Table 3). Soil temperature at 5 cm depth showed the largest range in HU between 0.4°C in early spring and 40.5°C in summer, whereas the lowest range was recorded in the UK between 0.6°C in winter and 14.3°C in summer. Volumetric soil moisture content was always higher than the wilting point at the UK, DK-B, DK-M, and the NL sites, but could approach the wilting point at the ES, IT and HU sites (Table 5). The lower soil moisture content in ES and IT than the wilting point is due to the offset caused by the stone fraction (>2 mm) of these soils, which is not included in the determination of the wilting point and field capacity.
SR varied among sites during the measurement periods (Table 3). Overall mean of observed SR rates ranged from 0.84 μmol CO2 m−2 s−1 at the HU site to 3.71 μmol CO2 m−2 s−1 at the DK-M site.
Temperature Control on SR
The best model fit based on Akaike’s Information Criteria (AICc) value, varied among sites (Table 4a; Figure 1). Refer to Table S2 for parameter estimates of the models.

Empirical temperature dependence functions of soil respiration (exponential as solid line, Lloyd–Taylor as dotted line and Gaussian as dashed line) fit to the data of the experimental sites with different climatic conditions. See Table S2 for parameter estimates of the functions.
In four of the seven study sites, the exponential soil temperature–respiration model was not supported by the empirical data (that is, Akaike weights were lower than 0.1). At the Mediterranean ES site, and the Continental HU site, with relatively wide soil temperature ranges (Table 3), we found the Gaussian temperature dependence function to be unequivocally the best model (Table 4b), whereas at the Atlantic heathland of the DK-M site, the Lloyd–Taylor and the Gaussian temperature dependence functions also achieved a low AICc value, that is, high Akaike weight. At the Atlantic heathland of DK-B, the Lloyd–Taylor model showed the lowest AICc value and was accepted with approximately a level of support in the data similar to that of the Gaussian model (see Akaike weights in Table 4b).
In the other three sites, including the Capo Caccia (IT) with Mediterranean climate, and the Atlantic heathlands of Oldebroek (NL) and Clocaenog (UK), the exponential model showed the lowest AICc value, whereas the other two models were also supported by the data (Table 4a, b). However, at the NL and UK sites, the Gaussian model had a c parameter of 0, which corresponds to the exponential model (see Table S2).
Additive and Interactive Soil Temperature and Soil Moisture Control on SR
Inclusion of soil moisture improved model performance in all cases. Table 5 shows the effects of soil moisture characteristics identified by conditional inference trees for the three temperature response functions of SR. We identified separate soil moisture intervals for every study site. Number of intervals ranges from 1 (DK-B, DK-M, NL) to 5 (HU, ES) (Table S2), suggesting the existence of thresholds in the soil moisture effect on SR. Thresholds identified at individual sites were very consistent across the three different models (Table S2). In additive models, functions fit for different soil moisture intervals differed in parameter a, which increased with increasing soil moisture, thus at the same temperature higher moisture resulted in higher SR.
Assuming interactions between soil moisture content and temperature, we found several soil moisture intervals that were homogeneous in the parameters of temperature dependence (Table S2). At most sites, we could not find any trend in the parameter values of temperature dependence functions fit with changes in soil moisture intervals, resulting in crossing curves in the plotted functions (Figure 2), suggesting that optimal soil temperature for SR depended also on moisture.


Empirical temperature dependence models with moisture integration best fit to the site data; every box represents one of the nine models in one site, whereas each curve within a box is an individual soil moisture category of the models (see also Table 5); A Kiskunság, HU; B Garraf, ES; C Capo Caccia, IT; D Mols, DK-M; E Brandbjerg, DK-B; F Oldebroek, NL; G Clocaenog, UK. Lines are black during the temperature intervals within which defined intervals of volumetric soil moisture contents occured in the field. Grey line segments mean extrapolated fittings outside the measured temperature range.
We found that in most cases (except the IT and the DK-B sites), only the models with an interactive soil moisture effect were supported by the empirical data. At the Mediterranean IT site, the exponential temperature model with both additive and interactive moisture models were supported, as well as the Gaussian and the Lloyd–Taylor temperature functions with an interactive moisture effect (Table 4). (The Gaussian model with additive soil moisture model had a c parameter of 0, which corresponds to the exponential model (see Table S2)). The other exception was the Atlantic DK-B site where models with an additive soil moisture effect performed better, and the three temperature models were almost equally supported (Table 4a, c). At the Atlantic DK-M site, the exponential function had unequivocally the best fit, whilst at the Atlantic NL and UK sites, the exponential and the Lloyd–Taylor temperature functions, each with interactive moisture effect, were supported by the data (all these models had an Akaike weight above 0.1). At the Mediterranean ES and the Continental HU sites, the Gaussian temperature model had the highest Akaike weight; at the HU site, this model could be found being unequivocally better than any others, whereas at the ES site also the Lloyd–Taylor temperature model proved to be supported, all with interactive soil moisture integration (Table 4a, c).
Thresholds in Soil Moisture Influencing Soil Temperature Dependence of SR
The applied method revealed significant soil moisture thresholds in the temperature dependence functions showing how the temperature sensitivity was altered at different soil moisture levels. Some thresholds identified by the best model fits were close to the field capacity or wilting points of the studied ecosystems (Table 5), others may reflect characteristic temperature and moisture relations of a given season, see below.
The Continental HU and the Mediterranean ES sites had the most thresholds: these were at the zone of limited water availability approaching the wilting point, and near field capacity (Table 5; Figure 2A, B). At the ES site, the curve that represented the highest soil moisture threshold (at 22.2 vol%, only found with the Lloyd–Taylor model) showed a decrease in SR under the highest soil moisture conditions, indicating lower microbial response to soil moisture during the colder days between November and March, when these higher soil moisture values occurred (Figure 2B). At the other Mediterranean site in IT, one threshold point was also found above the wilting point, and a second threshold (only found with the additive moisture model) between the wet (winter and spring) and the dry (summer and early autumn) periods (Table 5; Figure 2C). Similar to the curve of the Lloyd–Taylor model at the ES site, at the IT site the curve above this second threshold of the additive exponential model represents the wet season (highest soil moisture above 17.7 vol% and lowest soil temperature below 15°C) (Figure 2C). At the mesic DK-M, the only threshold for SR was found above the wilting point, but far below the field capacity value (Table 5; Figure 2D). The similar DK-B site also presented this threshold (Table 5; Figure 2E). At the mesic Atlantic NL site, the first threshold was found between the wilting point and field capacity, whereas the next threshold was found near the field capacity, close to the third threshold. At the NL site, the lowest SR rates were measured at soil moisture contents between 23.7 and 28.2 vol%, coinciding with the winter inactive period between October and March, whereas higher soil moisture occurred often in July and August. At soil moisture contents below 23.7 and above 28.2 vol%, the SR rates increased with increasing soil moisture (Table 5; Figure 2F). At the hydric Atlantic UK site, two thresholds for SR were found near field capacity and also far above field capacity. In this wet ecosystem, SR rates decreased with higher soil moisture content (Table 5; Figure 2G), because of anaerobic soil conditions.
Annual SR
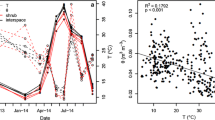
To compare the performances of the SR models, we calculated annual SR using the parameterized models (see model parameters in Table S2) and the daily meteorological data from the sites. The results for the six sites, HU, IT, DK-M, DK-B, NL and UK (Table 6) demonstrate that the annual SR estimated by the significant exponential models are in most cases higher than those estimated by the significant (DK-B) and non-significant (DK-M, NL) Gaussian models, however, the differences are mostly under 3%. Only at NL were the differences 7–25%. Also, for HU, the non-significant exponential model overestimated SR relative to the significant Gaussian model. Only at the IT and UK sites did the exponential models not predict higher annual SR than the other models. At the IT site, the models produced similar estimates. At the UK site, depending on year, the estimates were either not significantly different or the Gaussian model predicted 20% higher annual SR than the exponential and Lloyd–Taylor models. Relative to models without moisture effects, models that included soil moisture resulted in 8, 2 and 14% higher estimates of annual SR for the mesic sites DK-M, DK-B and NL, respectively. For the semiarid HU and the arid IT sites, models without moisture effects underestimated annual SR when it was humid in 2010, but overestimated annual SR in drier years. For the hydric UK site, this tendency was reversed, annual SR was overestimated by the models without moisture effects in the more humid years but underestimated annual SR in 2010 when precipitation was lowest (see Table 6). Soil organic matter content, used as a proxy for soil microbial activity, varied highly among the study sites (Table 2). Apart from the UK site, a significant relationship between annual SR and the soil organic matter content was found (Figure 3; r 2 = 0.961). However, at the UK site with considerably higher soil organic matter content, estimated annual SR was near the mean rate at the other sites (Figure 3), which is likely the result of anaerobic limitation of decomposition and the associated accumulation of organic matter at this site (Table 2).

Relationship between the average annual soil respiration values (g C m−2 y−1) calculated by the significant models and the soil organic carbon contents of six study sites: Kiskunság, Hungary (HU); Oldebroek, the Netherlands (NL); Brandbjerg, Denmark (DK-B); Mols, Denmark (DK-M); Capo Caccia, Italy (IT); Clocaenog, United Kingdom (UK). The ranges between the lowest and the highest annual soil respiration values calculated by all the presented models are shown to demonstrate the low interannual variability and the low variability of the model estimations compared to the high intersite variability. Apart from the UK site, a significant relationship between annual SR and the soil organic matter content is demonstrated.
Discussion
Temperature Control on SR
To accurately predict SR from ecosystems in future climates, it has become necessary to parameterise models with a wider range of temperatures than currently used. In this study, we examined the temperature response functions of SR at seven European shrubland sites of different climatic conditions from Atlantic heathlands through Mediterranean macchias to Continental poplar shrubland, thus extending the temperature and moisture range of our previous SR investigations (Lellei-Kovács and others 2011). In most previous field studies, the temperature–SR function used was fit to a relatively narrow range of soil temperatures, usually below 30°C. Typically, the exponential temperature function fits respiration data well in a relatively narrow temperature range below 30°C, whereas the relationship is weaker at higher temperatures. Thus, our approach increases the predictive power when forecasting the response to a warming future climate, if temperatures are expected to be higher than 30°C (Mielnick and Dugas 2000). When SR is studied under a wider range of temperatures, it is possible that the interaction of additional soil processes, such as substrate and water availability could alter respiration rates, resulting in lower respiration at higher soil temperatures (Ågren and others 1991; Tuomi and others 2008; Reichstein and Beer 2008; Lellei-Kovács and others 2011; González-Ubierna and others 2014).
In the present study, at the Atlantic sites, we could not find a model that unequivocally explained one of the temperature–SR relationships, that is, the exponential function fit was as good as the Gaussian and Lloyd–Taylor functions (Table 4b; Figure 1). This was probably due to the narrow temperature range, always under the optimum temperature, making it impossible to detect differences in the shape of the three models. Despite our efforts to obtain data that spanned a large temperature range by including climate change treatments (Table S1), the measurements taken at the Atlantic sites biased the data to a narrower range than anticipated, with the soil temperature rarely exceeding 20°C. At the Mediterranean IT site, where the exponential SR models performed the best, soil temperature remained within the bounds of 7–29°C. In this case, the relatively high winter soil temperature range was probably due to the strong moderating effect from the Mediterranean Sea that causes mild winter temperatures, in most cases above 10°C. It is therefore likely that at this site we were not able to detect either the lower or the upper temperature limitation on SR (Table 4b; Figure 1).
At the ES and HU sites, the Gaussian function was found to be the best performing temperature–SR function. The Gaussian function assumes that there is an optimal temperature for SR, which can be detected only when field measurements are performed in a sufficiently broad range of temperatures (Ågren and others 1991; Lellei-Kovács and others 2011; González-Ubierna and others 2014). The wide range of soil temperatures at the HU and ES sites (~40°C) may explain why the Gaussian function proved to be the best.
The Influence of Soil Moisture on the Temperature Sensitivity of SR
Our modelling approach integrated both soil moisture and temperature to examine the SR relationship. We revealed clear soil moisture thresholds in the temperature dependence of SR. This indicated that low soil moisture content was an important limiting factor of SR at both the seasonally dry Mediterranean and semiarid Continental sites, and also at the mesic Atlantic sites, whilst high soil moisture content imposing anaerobic conditions proved to limit SR at the hydric Atlantic site in the UK. In some cases, soil moisture thresholds could be connected to the wilting point or the field capacity (Table 5), but other thresholds might be related to more complex physiochemical or biological conditions (Robinson and others 2016), such as the effect of soil moisture content on the availability of various soluble substrates or the effect of specific microbial enzymes with characteristic kinetic properties (Davidson and others 2006). Kopittke and others (2014) reported that integration of soil moisture at the mesic Atlantic NL site did not improve the model fit of the temperature dependence of SR for control treatments, whereas it significantly improved the model fit for drought treatments. The lack of a moisture effect in control plots but appearance of an effect in the drought plots found by Kopittke and others (2014) support our analytical approach of using all treatment data together to cover a wider environmental range within the same model. Under Mediterranean climate at the IT site, de Dato and others (2010) showed a significant difference between temperature sensitivity of the wet vegetative season and the dry non-vegetative season between 2002 and 2004. At this site, we also found that the best fit of the exponential temperature function to the dataset between 2010 and 2011 was separated by soil moisture thresholds (Figure 2C). These two approaches gave similar results in ecosystems where vegetation periods are determined by water availability.
Similar to our results, soil moisture content has been shown to enhance the response of SR to temperature in a continental arid desert (Zhang and others 2010), in a semiarid steppe of Inner Mongolia (Chen and others 2009) and in an old-field climate change experiment (Suseela and others 2012). In the latter study, Suseela and others (2012) observed that both an upper and a lower soil moisture threshold related to SR activity existed, and that changes in soil structural properties during drought resulted in a hysteresis effect. Soil moisture thresholds were also found to change SR responses to temperature in other studies. Rey and others (2002) and Guidolotti and others (2013) found a soil moisture threshold in Mediterranean forests, below which there was no correlation between SR and soil temperature. In a study of temperate forest ecosystems, Wang and others (2006) found that increased temperature sensitivity (Q10) was related to increasing soil moisture content, but that Q10 declined after reaching a soil moisture threshold. Vicca and others (2014) also emphasized the importance of integrating soil moisture in the predictive models of SR, especially considering an altered moisture regime in the future. However, in the modelling approach of Vicca and others (2014) soil temperature is integrated as a simple exponential function, which may weaken the extensibility of the models. For comparison, for the dataset of the ES site, the exponential temperature and Gaussian moisture dependence (model 4 of Vicca and others 2014) achieved an AIC of 271.63. If both temperature and moisture dependence were modelled with the Gaussian function and their effect was additive, then an AIC of 201.75 was achieved. However, for the same dataset, our model with a Gaussian temperature function and interactive moisture thresholds achieved an AIC of 152.99, indicating a better performance of the model.
Our results showed that the SR relationship with soil moisture, the latter depending mostly on precipitation, is non-monotonic, which is congruent with the findings of Vicca and others (2014). In addition, at the plot scale this relationship can also be described as non-linear, with soil moisture thresholds being observed. We expect that the mechanisms that may explain our results are mediated by changes in the belowground community structure that are dependent on temperature and moisture (Ågren and Wetterstedt 2007).
Soil moisture impacts SR directly by changing soil microbial activity and altering soil structure and porosity, and also indirectly by affecting substrate availability (Davidson and others 2006). Under semiarid and arid conditions, there is a strong edaphic water limitation coupled with strong pulse dynamics of resources linked to changes in microclimate (Collins and others 2008; Maestre and others 2013). The close connection between substrate availability and soil processes is also demonstrated by the relationship between annual SR and soil organic carbon content at the studied sites (Figure 3). Similarly, Fernandez and others (2006) demonstrated the impact of soil organic carbon and nitrogen on SR through soil texture and soil moisture availability in a cold desert ecosystem. They found that when soil moisture and temperature are both favourable, soil organic carbon and nitrogen cannot be used to predict SR. A limitation of soil substrate availability for microbes may explain why the Gaussian-type soil temperature–SR model proved to be the best at the HU and ES sites (Table 5), where not only the temperature ranges were the largest (Table 3), but the soil organic matter content was also the lowest (Table 2).
Annual Scale Impacts on SR
The upscaled annual rates of SR showed profound differences among both years and models. As previously demonstrated at the HU site, annual SR rates calculated by the exponential function were systematically higher than those based on the Lloyd–Taylor and Gaussian functions (Lellei-Kovács and others 2011). In this modelling experiment, we also demonstrated that when excluding soil moisture from the models, modelled soil carbon fluxes may be overestimated especially for warm and dry years, which may be more frequent in the future. In the present study, annual SR values were also calculated from modelled data at six study sites (HU, IT, NL, DK-B, DK-M, UK) (see Table 6). We found that the rate of annual SR in NL was very similar to the amount calculated by a different methodology by Kopittke and others (2013, 2014) for the same period, which may validate these methods. Annual SR was also calculated in the work of de Dato and others (2010) for three study years between 2002 and 2004, the values calculated were between 927 and 1145 g C m−2 y−1, which are also similar to the values between 890 and 963 g C m−2 y−1 calculated by the method demonstrated here, for data between 2010 and 2011. At the UK site, annual SR decreased since 2000 because of a natural drought period that triggered an irreversible reduction in soil moisture and erosion of organic matter (Robinson and others 2016). In the period between 2010 and 2012, annual SR was around 400 g C m−2 y−1 (Domínguez and others 2015), which is also consistent with our results suggesting annual SR between 323 and 345 g C m−2 y−1.
At every site, the models that included soil moisture, always improved the model fit compared to those that excluded soil moisture. Furthermore, at three of the four Atlantic sites, including soil moisture resulted in higher estimated annual SR, independently of the applied temperature dependence function. At the IT and the HU sites, the direction of the alteration was dependent on the year: including soil moisture effect decreased the calculated annual SR in a drier year, and increased in a more humid year. The results are congruent with our previously published work, where excluding soil moisture resulted in an overestimation of rates of annual SR during a dry and hot year, but an underestimation of annual SR in a wet and cold year (Lellei-Kovács and others 2011). In the present study, which considered the period 2010 to 2012, variation in soil moisture resulted in a difference of 1 to 25% in the outputs from the nine different models we considered. This variation in output warrants further investigation into the uncertainty of model estimations and highlights the importance of appropriate model choice in the prediction of the future impacts of climate change on SR of different ecosystems.
Conclusions
In this study of European shrubland ecosystems under Atlantic, Mediterranean or Continental climate, we demonstrated that the temperature dependence function that best explains SR depended strongly on the temperature range where the study was conducted. We also showed that in these ecosystems when soil temperature range was above 30°C, the Gaussian function with optimum temperature provided a better fit to the data, than the exponential temperature function. Furthermore, we found that soil moisture strongly affected SR, not only in arid and semiarid, but also in mesic and hydric ecosystems, and the parameters of the temperature dependence functions changed significantly at distinctive soil moisture thresholds. These moisture thresholds may be connected to soil and ecosystem-specific variables, such as wilting point of the plants or field capacity of the soil. In years with high precipitation and in mesic and hydric ecosystems, the models that integrate moisture may estimate a higher level of annually respired carbon. These results highlight the importance of the choice from among the temperature dependence functions and the inclusion of soil moisture data when modelling SR, especially when predicting SR responses in a wide range of climatic conditions or in a changing climate.
References
Ågren GI, Wetterstedt JÅM. 2007. What determines the temperature response of soil organic matter decomposition? Soil Biol Biochem 39:1794–8.
Ågren GI, McMurtrie RE, Parton WJ, Pastor J, Shugart HH. 1991. State-of-the-art of models of production-decomposition linkages in conifer and grassland ecosystems. Ecol Appl 1:118–38.
Balogh J, Pintér K, Sz Fóti, Papp M, Cserhalmi D, Nagy Z. 2011. Dependence of soil respiration on soil moisture, clay content, soil organic matter, and CO2 uptake in dry grasslands. Soil Biol Biochem 43:1006–13.
Beier C, Emmett B, Gundersen P, Tietema A, Peňuelas J, Estiarte M, Gordon C, Gorissen A, Llorens L, Roda F, Williams D. 2004. Novel approaches to study climate change effects on terrestrial ecosystems in the field: drought and passive nighttime warming. Ecosystems 7:583–97.
Beier C, Emmett B, Tietema A, Schmidt IK, Peñuelas J, Kovács-Láng E, Duce P, de Angelis P, Gorissen A, Estiarte M, de Dato G, Sowerby A, Kröel-Dulay G, Lellei-Kovács E, Kull O, Mand P, Petersen H, Gjelstrup P, Spano D. 2009. Carbon and nitrogen balances for six shrublands across Europe. Glob Biogeochem Cycles 23:GB4008.
Billings SA, Ballantyne FIV. 2013. How interactions between microbial resource demands, soil organic matter stoichiometry, and substrate reactivity determine the direction and magnitude of soil respiratory responses to warming. Glob Change Biol 19:90–102.
Bradford MA, Davies CA, Frey SD, Maddox TR, Melillo JM, Mohan JE, Reynolds JF, Treseder KK, Wallenstein MD. 2008. Thermal adaptation of soil microbial respiration to elevated temperature. Ecol Lett 11:1316–27.
Burnham KP, Anderson DR. 2002. Model selection and multimodel inference: a practical information-theoretic approach. 2nd edn. New York: Springer.
Chen S, Lin G, Huang J, Jenerette GD. 2009. Dependence of carbon sequestration on the differential responses of ecosystem photosynthesis and respiration to rain pulses in a semiarid steppe. Glob Change Biol 15:2450–61.
Collins SL, Sinsabaugh RL, Crenshaw C, Green L, Porras-Alfaro A, Stursova M, Zeglin LH. 2008. Pulse dynamics and microbial processes in aridland ecosystems. J Ecol 96:413–20.
Curiel Yuste J, Ma S, Baldocchi DD. 2010. Plant-soil interactions and acclimation to temperature of microbial-mediated soil respiration may affect predictions of soil CO2 efflux. Biogeochemistry 98:127–38.
Davidson EA, Janssens IA. 2006. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 440:165–73.
Davidson EA, Belk E, Boone RD. 1998. Soil water content and temperature as independent or confounded factors controlling soil respiration in temperate mixed hardwood forest. Glob Change Biol 4:217–27.
Davidson EA, Janssens IA, Luo Y. 2006. On the variability of respiration in terrestrial ecosystems: moving beyond Q10. Glob Change Biol 12:154–64.
de Dato GD, de Angelis P, Sirca C, Beier C. 2010. Impact of drought and increasing temperatures on soil CO2 emissions in a Mediterranean shrubland (gariga). Plant Soil 327:153–66.
Domínguez MT, Sowerby A, Smith AR, Robinson DA, Van Baarsel S, Mills RTE, Marshall MR, Koller E, Lebron I, Hall J, Emmett BA. 2015. Sustained impact of drought on wet shrublands mediated by soil physical changes. Biogeochemistry 122:151–63.
Emmett B, Beier C, Estiarte M, Tietema A, Kristensen HL, Williams D, Peñuelas J, Schmidt I, Sowerby A. 2004. The response of soil processes to climate change: results from manipulation studies of shrublands across an environmental gradient. Ecosystems 7:625–37.
Fernandez DP, Neff JC, Belnap J, Reynolds RL. 2006. Soil respiration in the cold desert environment of the Colorado Plateau (USA): abiotic regulators and thresholds. Biogeochemistry 78:247–65.
González-Ubierna S, de la Cruz MT, Casermeiro MÁ. 2014. Climate factors mediate soil respiration dynamics in Mediterranean agricultural environments: an empirical approach. Soil Res 52:543–53.
Guidolotti G, Rey A, D’Andrea E, Matteucci G, De Angelis P. 2013. Effect of environmental variables and stand structure on ecosystem respiration components in a Mediterranean beech forest. Tree Physiol 33:960–72.
Hothorn T, Hornik K, Zeileis A. 2006. Unbiased recursive partitioning: a conditional inference framework. J Comput Graph Stat 15:651–74.
Howard DM, Howard PJA. 1993. Relationships between CO2 evolution, moisture content and temperature for a range of soil types. Soil Biol Biochem 25:1537–46.
IPCC 2013. The physical science basis. In: Stocker TF, Qin D, Plattner G-K, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM, Eds. Climate change 2013: contribution of working group I to the fifth assessment report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press.
Johnson JB, Omland KS. 2004. Model selection in ecology and evolution. Trends Ecol Evol 19:101–8.
Kirschbaum MUF. 2004. Soil respiration under prolonged soil warming: are rate reductions caused by acclimation or substrate loss? Glob Change Biol 10:1870–7.
Kopittke GR, van Loon EE, Tietema A, Asscheman D. 2013. Soil respiration on an aging managed heathland: identifying an appropriate empirical model for predictive purposes. Biogeosciences 10:3007–38.
Kopittke GR, Tietema A, van Loon EE, Asscheman D. 2014. Fourteen annually repeated droughts suppressed autotrophic soil respiration and resulted in an ecosystem change. Ecosystems 17:242–57.
Kotroczó Zs, Veres Zs, Fekete I, Krakomperger Zs, Tóth JA, Lajtha K, Tóthmérész B. 2014. Soil enzyme activity in response to long-term organic matter manipulation. Soil Biol Biochem 70:237–43.
Leirós MC, Trasar-Cepeda C, Seoane S, Gil-Sotres F. 1999. Dependence of mineralization of soil organic matter on temperature and moisture. Soil Biol Biochem 31:327–35.
Lellei-Kovács E, Kovács-Láng E, Kalapos T, Botta-Dukát Z, Barabás S, Beier C. 2008. Experimental warming does not enhance soil respiration in a semiarid temperate forest-steppe ecosystem. Commun Ecol 9:29–37.
Lellei-Kovács E, Kovács-Láng E, Botta-Dukát Z, Kalapos T, Emmett B, Beier C. 2011. Thresholds and interactive effects of soil moisture on the temperature response of soil respiration. Eur J Soil Biol 47:247–55.
Lloyd J, Taylor JA. 1994. On the temperature dependence of soil respiration. Funct Ecol 8:315–23.
Maestre FT, Escolar C, de Guevara ML, Quero JL, Lázaro R, Delgado-Baquerizo M, Ochoa V, Berdugo M, Gozalo B, Gallardo A. 2013. Changes in biocrust cover drive carbon cycle responses to climate change in drylands. Glob Change Biol 19:3835–47.
Mielnick PC, Dugas WA. 2000. Soil CO2 flux in a tallgrass prairie. Soil Biol Biochem 32:221–8.
Mikkelsen TN, Beier C, Jonasson S, Holmstrup M, Schmidt IK, Ambus P, Pilegaard K, Michelsen A, Albert K, Andresen LC, Arndal MF, Bruun N, Christensen S, Danbæk S, Gundersen P, Jørgensen P, Linden LG, Kongstad J, Maraldo K, Priemé A, Riis-Nielsen T, Ro-Poulsen H, Stevnbak K, Selsted MB, Sørensen P, Larsen KS, Carter MS, Ibrom A, Martinussen T, Miglietta F, Sverdrup H. 2008. Experimental design of multifactor climate change experiments with elevated CO2, warming and drought: the CLIMAITE project. Funct Ecol 22:185–95.
Moyano FE, Manzoni S, Chenu C. 2013. Responses of soil heterotrophic respiration to moisture availability: an exploration of processes and models. Soil Biol Biochem 59:72–85.
Murthy R, Griffin KL, Zarnoch SJ, Dougherty PM, Watson B, Haren JV, Patterson RL, Mahato T. 2003. Carbon dioxide efflux from a 550 m2 soil across a range of soil temperatures. For Ecol Manag 178:311–27.
Pavelka M, Acosta M, Marek MV, Kutsch W, Janous D. 2007. Dependence of the Q10 values on the depth of the soil temperature measuring point. Plant Soil 292:171–9.
Peñuelas J, Prieto P, Beier C, Cesaraccio C, de Angelis P, de Dato G, Emmett BA, Estiarte M, Garadnai J, Gorissen A, Kovács-Láng E, Kröel-Dulay G, Llorens L, Pellizzaro G, Riis-Nielsen T, Schmidt IK, Sirca C, Sowerby A, Spano D, Tietema A. 2007. Response of plant species richness and primary productivity in shrublands along a north-south gradient in Europe to seven years of experimental warming and drought: reductions in primary productivity in the heat and drought year of 2003. Glob Change Biol 13:2563–81.
Qi Y, Xu M, Wu J. 2002. Temperature sensitivity of soil respiration and its effects on ecosystem carbon budget: nonlinearity begets surprises. Ecol Model 153:131–42.
R Development Core Team. 2008. R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. ISBN 3-900051-07-0. http://www.R-project.org.
Reichstein M, Beer C. 2008. Soil respiration across scales: the importance of a model-data integration framework for data interpretation. J Plant Nutr Soil Sci 171:344–54.
Reichstein M, Tenhunen JD, Roupsard O, Ourcival JM, Rambal S, Miglietta F, Peressotti A, Pecchiari M, Tirone G, Valentini R. 2002. Severe drought effects on ecosystem CO2 and H2O fluxes at three Mediterranean evergreen sites: revision of current hypotheses? Glob Change Biol 8:999–1017.
Rey A, Pegoraro E, Tedeschi V, De Parri I, Jarvis PG, Valentini R. 2002. Annual variation in soil respiration and its components in a coppice oak forest in Central Italy. Glob Change Biol 8:851–66.
Robinson DA, Jones SB, Lebron I, Reinsch S, Domínguez MT, Smith AR, Jones DL, Marshall MR, Emmett BA. 2016. Experimental evidence for drought induced alternative stable states of soil moisture. Sci Rep 6:20018.
Rodrigo A, Recous S, Neel C, Mary B. 1997. Modelling temperature and moisture effects on C-N transformations in soils: comparison of nine models. Ecol Model 102:325–39.
Sardans J, Peñuelas J, Estiarte M, Prieto P. 2008. Warming and drought alter C and N concentration, allocation and accumulation in a Mediterranean shrubland. Glob Change Biol 14:2304–16.
Saxton KE, Rawls WJ. 2006. Soil water characteristic estimates by texture and organic matter for hydrologic solutions. Soil Sci Soc Am J 70:1569–78.
Shen Z, Shi B, Wang B, Jiang H-J. 2013. The temperature dependence of soil organic matter decomposition and CO2 efflux: a review. Acta Ecol Sin 33:3011–19 (in Chinese).
Skopp J, Jawson MD, Doran JW. 1990. Steady-state aerobic microbial activity as a function of soil water content. Soil Sci Soc Am J 54:1619–25.
Sowerby A, Emmett B, Tietema A, Beier C. 2008. Contrasting effects of repeated summer drought on soil carbon efflux in hydric and mesic heathland soils. Glob Change Biol 14:2388–404.
Subke J-A, Bahn M. 2010. On the temperature sensitivity of soil respiration: can we use the immeasurable to predict the unknown? Soil Biol Biochem 42:1653–6.
Suseela V, Conant RT, Wallenstein MD, Dukes JS. 2012. Effects of soil moisture on the temperature sensitivity of heterotrophic respiration vary seasonally in an old-field climate change experiment. Glob Change Biol 18:336–48.
Tuomi M, Vanhala P, Karhu K, Fritze H, Liski J. 2008. Heterotrophic soil respiration—comparison of different models describing its temperature dependence. Ecol Model 211:182–90.
Vicca S, Bahn M, Estiarte M, van Loon EE, Vargas R, Alberti G, Ambus P, Arain MA, Beier C, Bentley LP, Borken W, Buchmann N, Collins SL, de Dato G, Dukes JS, Escolar C, Fay P, Guidolotti G, Hanson PJ, Kahmen A, Kröel-Dulay G, Ladreiter-Knauss T, Larsen KS, Lellei-Kovács E, Lebrija-Trejos E, Maestre FT, Marhan S, Marshall M, Meir P, Miao Y, Muhr J, Niklaus PA, Ogaya R, Peñuelas J, Poll C, Rustad LE, Savage K, Schindlbacher A, Schmidt IK, Smith AR, Sotta ED, Suseela V, Tietema A, van Gestel N, van Straaten O, Wan S, Weber U, Janssens IA. 2014. Can current moisture responses predict soil CO2 efflux under altered precipitation regimes? A synthesis of manipulation experiments. Biogeosciences 11:2991–3013.
von Lützow M, Kögel-Knabner I. 2009. Temperature sensitivity of soil organic matter decomposition—what do we know? Biol Fertil Soils 46:1–15.
Wang WJ, Dalal RC, Moody PW, Smith CJ. 2003. Relationships of soil respiration to microbial biomass, substrate availibility and clay content. Soil Biol Biochem 35:273–84.
Wang C, Yang J, Zhang Q. 2006. Soil respiration in six temperate forests in China. Glob Change Biol 12:2103–14.
Wang B, Zha TS, Jia X, Wu B, Zhang YQ, Qin SG. 2014. Soil moisture modifies the response of soil respiration to temperature in a desert shrub ecosystem. Biogeosciences 11:259–68.
Wu Z, Dijkstra P, Koch GW, Peñuelas J, Hungate BA. 2011. Responses of terrestrial ecosystems to temperature and precipitation change: a meta-analysis of experimental manipulation. Glob Change Biol 17:927–42.
Xu L, Baldocchi DD, Tang J. 2004. How soil moisture, rain pulses, and growth alter the response of ecosystem respiration to temperature. Glob Biogeochem Cycles 18:GB4002.
Yan L, Chen S, Huang J, Lin G. 2011. Water regulated effects of photosynthetic substrate supply on soil respiration in a semiarid steppe. Glob Change Biol 17:1990–2001.
Zeileis A, Hothorn T, Hornik K. 2005. Model-based recursive partitioning. J Comput Graph Stat 17:492–514.
Zhang LH, Chen YN, Zhao RF, Li WH. 2010. Significance of temperature and soil water content on soil respiration in three desert ecosystems in Northwest China. J Arid Environ 74:1200–11.
Zhou X, Sherry RA, An Y, Wallace LL, Luo Y. 2006. Main and interactive effects of warming, clipping, and doubled precipitation on soil CO2 efflux in a grassland ecosystem. Glob Biogeochem Cycles 20:GB1003.
Zhou X, Wan SQ, Luo YQ. 2007. Source components and interannual variability of soil CO2 efflux under experimental warming and clipping in a grassland ecosystem. Glob Change Biol 13:761–75.
Acknowledgments
We gratefully acknowledge the support of the INCREASE Project (http://increase.ku.dk) funded by the EC FP7-Infrastructure-2008-1 Grant Agreement 227628, and the Hungarian Scientific Research Fund (OTKA K112576 and PD115637). ME and JP research was supported by the European Research Council Synergy Grant ERC-2013-SyG-610028 IMBALANCE-P, the Spanish Government Grant CGL2013-48074-P and the Catalan Government Grant SGR 2014-274. We thank the two anonymous reviewers for helpful comments and suggestions.
Author information
Authors and Affiliations
Corresponding author
Additional information
Author contributions
EKL and ELK conceived the study; ELK, GDD, ME, GG, GRK, GKD, KSL, JP, ARS, AS, AT and IKS performed the research; BDZ and ELK analyzed the data; and ELK wrote the paper.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Lellei-Kovács, E., Botta-Dukát, Z., de Dato, G. et al. Temperature Dependence of Soil Respiration Modulated by Thresholds in Soil Water Availability Across European Shrubland Ecosystems. Ecosystems 19, 1460–1477 (2016). https://doi.org/10.1007/s10021-016-0016-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-016-0016-9