
Abstract
Glutathione-S-transferase(s) (GST) enzyme from Brugia malayi has been exploited as a target in lymphatic filariasis therapeutics. An active GST is a homodimer of a 208 residue long monomer consisting of two domains, a smaller α/β domain and a larger α domain. The components of the glutathione (GSH) system, mainly GST enzymes, are critical antioxidant and detoxification system responsible for the long-term existence of filarial worms in mammalian host; hence they are major chemotherapeutic targets in filarial species. In the present study, 58 phytochemicals from 10 plants, predicted and reported to have potential nematicidal activity and ADMET satisfaction, have been docked to GST enzyme of B. malayi to assess their binding affinity and consequently their inhibitory activity. A comparative study has been made with commonly employed chemotherapeutic GST inhibitors such as cibacron-blue, butylated hydroxyanisole, hexyl glutathione and ethacrynic acid. In vitro effects of potential drug like compound from in silico results have been done for validation of docking studies. In vitro assay revealed efficacy in GST inhibition in the following compounds: linalool (97.50%), alpha-pinene (90.00%), strychnine (87.49%), vanillin (84.99%), piperine (79.99%), isoeugenol (62.49%), curcumin (57.49%), beta-caryophyllene (39.50%), cinnamic acid (27.49%), capsaicin (19.99%), citronellol (19.99%) and geraniol (17.49%). An online database (www.spicebioinfo.res.in/gstleadbase) has been developed, which will serve as a useful repository of information on GST inhibitors for future development of drugs against filarial nematodes. These findings thus suggest that the above phytochemicals could be potentially developed as lead molecules for targeting GST of lymphatic filarial parasites.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Lymphatic filariasis (LF) is a mosquito-borne tropical disease caused by the nematode parasites Wuchereria bancrofti, Brugia malayi and B. timori [1]. It is the major cause of acute and chronic morbidity in 81 countries in Asia-Pacific, Africa and the Americas. Approximately 1.3 billion people living in these regions are at risk of infection [2]. The adult parasites live 5 to 10 years, of which the fecund life span is 4 to 6 years. Several hundreds to thousands of infective mosquito bites are necessary to establish infection. Of these, three parasites W. bancrofti accounts for nearly 90% of LF infections worldwide. B. malayi is prevalent only in some parts of South and Southeast Asia, and B. timori is found only in Indonesia. The drugs used for treating LF include annual doses of diethylcarbamazine (DEC), DEC plus albendazole, or ivermectin plus albendazole; none of these is effective in killing adult worms, and treatments are therefore aimed at reducing transmission and pathology [3, 4]. Since an effective treatment for filarial adult worms is currently unavailable, new chemical classes of compounds with macrofilaricidal activities are now required [5]. Recently Srinivasan et al. [6] reported ethacrynic acid, plumbagin and curcumin as inhibitory compounds against GSTs of bovine filarial worms Setaria digitata. Only a few studies have reported the use of phytochemicals as GST inhibitors.
The components of the glutathione (GSH) system GSTs (glutathione-S-transferase(s)) and GSHPx (glutathione peroxidases) are the major defense systems present in filarial nematodes. The role of this secreted enzyme is the inhibition of the oxidative burst of leukocytes and neutralization of secondary products of lipid peroxidation, thus providing an explanation for the resistance of these parasites to immune effector mechanisms and their persistence in the mammalian host [7]. The mechanism of action of GST(s) (E.C.2.5.1.18), a large family of multifunctional dimeric enzymes includes defense against oxidative attack via conjugation of electrophiles to glutathione and reduction of lipid hydroperoxides [8]. Due to their primary role in drug metabolism, GSTs have been the recent focus of research as a potential drug target for anti-schistosomal [9], antimalarial [10, 11], and antifilarial [12–14] drug development. In addition to their isomerization and GSH conjugation activities, in mammals these enzymes contribute to defense against oxidative stress, by virtue of both their selenium-independent GSH peroxidase activities [15]. Inhibition of parasitic GST affects the survival of the parasites or helps in the enhancement of activity of presently available antifilarial drugs [16]. GST from human filarial parasites is significantly different from human GST in sequence and structure [17]. Hence B. malayi GST was exploited to design new target based chemotherapeutic agents.
An active GST is a homodimer of a 208 residue long monomer consisting of two domains (smaller α/β domain and larger α domain) (Fig. 1). The N-terminal small domain (residues 1 to 74) is an α/β structure with the folding topology βαβαββα arranged in the order β2, β1, β3 and β4 with β3 anti-parallel to the others, forming a regular β-sheet with a right-handed twist surrounded by three α-helices. The C terminal, large domain 2 (82–208 residues) is α-helical. The residues that interface the two βαβ and ββα motifs are Trp38, Phe8, Val33, Cys47, Leu52 and Leu43 in human π GST. In Bm-GST the residues Val33, Cys47 and Leu43 are replaced by Ile 38, Phe47 and Met43 [17]. The secondary structure of B. malayi GST has been generated by GenTHREADER – Protein fold recognition software (http://bioinf.cs.ucl.ac.uk/threader/) [18].

Secondary structure of Brugia malayi GST: Generated by GenTHREADER – Protein fold recognition software (http://bioinf.cs.ucl.ac.uk/threader/) [18]
A wide range of chemical compounds including alkaloids, coumarins, flavonoids, benzofurans, terpenoids and steroids have been isolated from various plant extracts and these have been found to possess various pharmacological, nematicidal and insecticidal activities. A comprehensive review of chemical constituents and pharmacological profiles of 10 selected medicinal plants, including spices, have led to the identification of potential nematicides. These new nematicides of natural origin may lead to higher safety and efficiency in nematode control and nematicidal drug development.
Historically, herbs, shrubs and spices have enjoyed a rich tradition of use for their flavor enhancement characteristics and medicinal properties [19, 20]. Spices hold the promise of providing both significant clinical benefits and key insights into the pathophysiology of cancer, arthritis, inflammation, respiratory disorders, gastrointestinal disturbances, allergy and microbial infections. Numerous demonstrations of preclinical efficacy of turmeric and various spices in animal models for preventing cancer and cardiovascular disorders have been reported [21]. This study was conducted with the objective of exploring the nematicidal activity of herbs and spices, with special reference to its potential to inhibit GST activity.
Materials and methods
Database screening and activity prediction
Initially, a review of plants with nematicidal property was made, which helped us identify 10 spices and medicinal plants: coriander, cassia, turmeric, allspice, cinnamon, strychnous, lemongrass, garlic, litsea and vanilla. The chemical compounds from these plants were collected through literature search and from Dr. Duke’s phytochemical and ethno-botanical databases (http://ars-grin.gov/duke/). The screening results revealed the presence of 128 nematicidal phytochemicals in these plants. The PASS server [22] was used to predict nematicidal activity and GST substrate activity of the phytochemicals (http://195.178.207.233/PASS/AP.html); PreADMET server (http://preadmet.bmcrd.org/) was used to predict the drug-likeness and ADME-Tox (Absorption, Distribution, Metabolism Excretion and Toxicity) properties [23]. The ADME-Tox properties of a compound together with its pharmacological properties such as drug likeness are conventionally a part of drug development. The compounds obeying the ADMET rules and drug likeness rules were short listed for docking studies.
Ligand structure
The canonical smiles notations of phytochemicals were collected from PubChem (http://pubchem.ncbi.nlm.nih.gov/), ChemSpider (http://chemspider.com) and DrugBank (http://www.drugbank.ca/). The 3D structures of compounds were developed by 3D Structure Generator CORINA [24, 25] using canonical smiles of the compound. Energy minimization and molecular optimization of all compounds were done using Arguslab 4.0.1 [26]. Geometry optimization was carried out using AM1 (Austin Model 1), semi-empirical quantum mechanics force field in Arguslab4.0.1. The best conformer thus obtained was based on energy minimization and geometry optimization. The final structures exhibiting lowest energy were saved in *.pdb format for input in to MVD environment.
Target protein structure
Theoretically solved structure of Brugia malayi glutathione-S-transferase was selected as the target for docking study, since to date there are no experimentally solved structures for Wuchereria bancrofti, Brugia malayi or B. timori. The GST structure was downloaded from Protein Data Bank (PDB id - 1SJO) and the structure optimized using Swiss PDB viewer software. Three active sites were detected in the GST enzyme of B. malayi using Molegro Virtual Docker. Active site residues of the GST enzyme were predicted using WHATIF server (http://swift.cmbi.ru.nl/servers/html/index.html) [27]. Active site residues predicted by WHATIF were Tyr7, Tyr101, Tyr106, Phe8, Phe45, Phe47, Phe155, Pro9, Pro51, Pro201, Ile10, Ile33, Ile105, Ile200, Arg11, Arg32, Arg95, Gly12, Gly48, Gly64, Leu13, Leu50, Asn34, Asn203, Ala35, Try38, Lys42, Lys103, Gln49, Gln62, Ser63, His98, Thr99, Thr102, Asp159, Val61, and Val202. Docking was carried out using single active-site having large volume (79.608 Å3) among the three cavities. This cavity was chosen since it binds glutathione with the highest specificity compared to the other sites, when docked with the whole protein using MVD. Amino acid residues present in the active site selected for docking and grid generation were Gln49, Gln12, Ile10, Ile105, Gly12, Ser63, His98, Tyr7, Tyr101, Tyr106, Pro9, Pro51, Pro201, Arg11, Arg32, Arg95, Thr99, Thr102, Val61, Val202, Lys42 and Lys103.
Molecular docking
Molecular docking study was carried out by using Molegro Virtual Docker [28]. The entire protein structure was loaded on to MVD platform for docking process. MVD performs flexible ligand docking, so the optimal geometry of the ligand is determined during the docking. MVD includes MolDock Score [28] and PLANTS Score [29] for evaluating docking solutions. MVD returns multiple poses representing different potential binding modes. This can be useful when the best-scoring (i.e., lowest-energy) pose does not represent the native binding mode or when multiple binding modes exist. Here clustering has been used to reduce the number of poses found during the docking run and only the most promising ones are reported. Compounds with the lowest dock score and high interaction with active-site was taken for in vitro studies based on the availability of the compound.
In vitro GST assay
The phytochemicals β-caryophyllene, capsaicin, cinnamic acid, citronellol, curcumin, eugenol, geraniol, isoeugenol, linalool, myristicin, neral, α-pinene, piperine, terpineol, vanillin and strychnine were purchased in the pure form from Sigma Chemicals, USA; glutathione (GSH) and 1-chloro-2, 4-dinitrobenzene (CDNB) were purchased from Sisco Research Laboratories Pvt. Ltd., (Mumbai, India). Dirofilaria immitis microfilaria, the canine filarial nematode used for in vitro study, was obtained from the District Veterinary Centre Campus, Calicut, Kerala.
GST crude enzyme was obtained by centrifuging the serum containing ∼2000 filarial nematodes at 1000 rpm for 2 min, and washing twice with phosphate buffered saline (PBS) at pH 7.4. The nematodes were ground with micro pestle and glass powder. The solution was centrifuged at 10000 rpm at 4 °C for 30 min. Supernatant was dialyzed against PBS overnight and made up to 2 ml. The following phytochemicals were used to study their GST inhibitory activity, at a concentration of 0.001 mg ml−1 in ethanol: β-caryophyllene, capsaicin, cinnamic acid, citronellol, curcumin, eugenol, geraniol, isoeugenol, linalool, myristicin, neral, α-pinene, piperine, terpineol, vanillin and strychnine (dissolved in water).
The dialyzed enzyme fraction (0.1 ml) was incubated in the presence of 1 ml of 0.001 mg ml−1 concentration of the phytochemicals listed above, in the presence of 1 mM glutathione reduced (GSH), and 0.1 M phosphate buffer, pH 6.5, for 1 hour at room temperature. A control containing ethanol was also maintained. GST activity was measured using the method of Habig et al. [30], by initiating the reaction with the addition of 1 mM 1-chloro-2,4-dinitrobenzene (CDNB) and following the change in absorbance at 340 nm, in a Shimadzu 1601 UV-Visible spectrophotometer. The GST activity was expressed as change in absorbance at 340 nm per minute per ml crude enzyme extract. Two replicates of each treatment were maintained.
Results and discussion
Biological activity prediction yielded 58 of the 128 phytochemicals with nematicidal, anti-helmintic and GST substrate activities. These phytochemicals also satisfied both ADME-Tox and drug likeness rules and were selected for docking studies. Docking results showed that all 58 compounds docked satisfactorily to the GST enzyme active site with good docking scores of less than −51.532 kcal mol−1. Hence these phytochemicals of comparatively less docking energy and greater number of hydrogen bond interactions were selected as promising lead compounds after docking studies (Table 1).
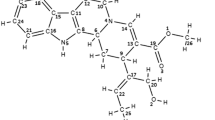
An in silico study was performed to compare the binding affinity of commonly employed chemotherapeutic GST inhibitor substances such as cibacron-blue, butylated hydroxyanisole, hexyl glutathione and ethacrynic acid, with the phytochemicals used for this study. Several potent phytochemicals that possess docking scores very similar to the current GST inhibitor drugs were identified (Table 2). Among the GST inhibitors butylated hydroxyanisole (BHA) markedly reduces worm viability [31]. BHA had a docking score of −68.431 kcal mol−1; phytochemicals with lower docking score than BHA are potential GST inhibitors, and represent promising starting points as lead compounds to treat LF. Thus phytochemicals which exhibit low dock scores and strong hydrogen bond interaction energy and greater number of hydrogen bonds in docking studies such as curcumin (PubChem CID: 969516; MW: 368.380 g mol−1), vanillin (PubChem CID: 1183; MW: 152.147 g mol−1), strychnine (PubChem CID: 5979; MW: 334.412 g mol−1), genostrychnine (PubChem CID: 73393; MW: 350.411 g mol−1), brucine (PubChem CID: 442021; MW: 394.464 g mol−1), brucine-n-oxide (PubChem CID: 161215; MW: 410.463 g mol−1), beta-colubrine (PubChem CID: 10512; MW: 364.438 g mol−1) and linalool (PubChem CID: 6549; MW: 154.249 g mol−1) are promising hits as GST inhibitors of natural origin. The hydrogen bond interaction of these lead compounds with the target residues is shown in Fig. 2. The analysis showed that curcumin has greater number of H-bond interactions and strychnine the least. Table 3 shows interacting properties of the eight highly docked phytochemicals to the target protein.


Docking view showing Hydrogen bond interaction of ligands with residues in active site of GST enzyme. (a) curcumin, (b) brucine-n-oxide,(c) beta-colubrine, (d) brucine, (e) genostrychnine, (f) strychnine, (g) vanillin and (h) linalool
GST is extensively investigated as a major target against several parasitic infections [31–35]. GST of filarial nematodes has very similar function and multiple sequence analysis revealed its similarity in sequence level (Fig. 3). There is no sequence in public domain of GST of B. timori to compare its relatedness to other LF GSTs. GST protein of B. malayi (BmGST) and W. bancrofti (WbGST) shares 98% similarity and GST of the canine filariasis nematode, Dirofilaria immitis (DiGST) shares only 74% similarity to BmGST and 75% to WbGST. We used D. immitis GST for in vitro studies, since the other filarial nematodes were not available.

Multiple sequence alignment (ClustalW) of GST sequences of B. malayi, D. immitis and W. bancrofti, which show that these sequences are identical. BmGST (Y12788) and WbGST (AY195867) show 98% similarity, BmGST and DiGST (P46426) share 74% similarity and WbGST and DiGST 75% similarity. The (*) denotes identical bases, (:) denotes strongly similar amino acids and (.) denotes weakly similar amino acids
In vitro studies indicated that linalool (97.50%), alpha-pinene (90.00%), strychnine (87.49%), vanillin (84.99%), piperine (79.99%), isoeugenol (62.49%), curcumin (57.49%), beta-caryophyllene (39.50%), cinnamic acid (27.49%), capsaicin (19.99%), citronellol (19.99%) and geraniol (17.49%) have good potential as nematicidal compounds against filarial GST (Table 4). Molecular structures of these compounds are given in Fig. 4. Terpineol, neral and myristicin had no detectable inhibitory effect. These in vitro studies help validate the results obtained from in silico docking studies. The reason why in vitro activities do not correlate closely with in silico docking could be because we have used B. malayi GST for in silico studies, while in vitro studies were carried out with D. immitis, due to the difficulty in obtaining samples of B. malayi. Multiple sequence alignment of GST sequences revealed that BmGST and DiGST (P46426) share only 74% similarity (Fig. 3). The difference in the correlation between the in silico and in vitro results could be attributed to the structural differences among the BmGST and DiGST.



Molecular structures of assayed compounds - Structures drawn using ChemSketch software
Curcumin is being used for treatment of cancer, wounds and as a cosmetic [36] among other medicinal uses. GST inhibiting activity of curcumin has been identified in various organisms [37, 38] and its worm motility inhibition was found to be effective at 54.29 μM [6]; the dried seed of Strychnos nux-vomica L., has been effectively used in Chinese folk medicine for the treatment of liver cancer and associated pathological abnormalities for ages [39]. Vanilla is a valued spice for its aroma and flavor. The anti-inflammatory activity of linalool has been reported earlier [40]. Piperine, a bioavailability enhancer from black pepper (Piper spp.), has already been reported to inhibit glucuronidation activity in rats and guinea pigs [41]. Singh et al. [42] reported that piperine inhibited rat hepatocyte-mediated glucuronidation of 3-hydroxybenzo[a]pyrene with an IC50 of 50 μmol L−1. Co-administration of piperine and curcumin to humans and rats enhanced the bioavailability of curcumin by 2000% and 154%, respectively [43]. Isoeugenol is a well-known antioxidant and its other biological activities include anti-inflammatory, antibiotic, antioxidant, anti-carcinogenic and local anaesthetic activities.
Since the above studied compounds are of natural origin, which satisfies both ADMET and drug likeness properties, these compounds can be used as potent lead compounds against filarial parasites. An online database (www.spicebioinfo.res.in/gstleadbase) has been developed, which it is hoped will serve as a useful repository of information on GST inhibitors for future development of drugs against filarial nematodes.
Conclusions
To summarize, we have employed virtual screening protocol, molecular docking to identify potential drug-like inhibitors of the detoxifying enzyme - GST - of Brugia malayi. Several potential drug-like inhibitors have been screened and found to interact with GST satisfactorily. Phytochemicals like curcumin, brucine-n-oxide, beta-colubrine, brucine, genostrychnine, strychnine, vanillin and linalool revealed strong binding with less docking scores and more number of hydrogen bond interactions to GST of B. malayi. This in vitro and in silico docking study validates GST inhibitory activity of compounds such as linalool, alpha-pinene, strychnine, vanillin, piperine, isoeugenol, curcumin, beta-caryophyllene, cinnamic acid, capsaicin, citronellol and geraniol, hence these compounds are novel, alternative drug therapy, of natural origin, for treatment of filariasis through inhibition of GST. Further studies are required to mark them as lead compounds for the development of novel drugs against lymphatic filariasis.
References
WHO (2002) Lymphatic Filariasis: The Disease and Its Control. Technical Report 71. Geneva
WHO (2008) Global programme to eliminate lymphatic filariasis. Wkly Epidemiol Rec 83:333–341
Liu LX, Weller PF (1996) Antiparasitic drugs. N Engl J Med 334:1178–1184
Oliveira-Menezes A, Lins R, Norões J, Dreyer G, Lanfredi RM (2007) Comparative analysis of a chemotherapy effect on the cuticular surface of Wuchereria bancrofti adult worms in vivo. Parasitol Res 101:1311–1317
Bajpai P, Verma SK, Katiyar D, Tewari N, Tripathi RP, Bansal I, Saxena JK, Misra-Bhattacharya S (2005) Search for new prototypes for the chemotherapy of filariasis: a chemotherapeutic and biochemical approach. Parasitol Res 95:383–390
Srinivasan L, Mathew N, Muthuswamy K (2009) In vitro antifilarial activity of glutathione-S-transferase inhibitors. Parasitol Res 105:1179–1182
Cookson E, Blaxter ML, Selkirk ME (1992) Identification of the major soluble cuticular glycoprotein of lymphatic filarial nematode parasites (gp29) as a secretory homolog of glutathione peroxidase (filariasis/Brugia/antioxidant/surface protein). Proc Natl Acad Sci USA 89:5837–5841
Ahmad R, Srivastava AK (2008) Inhibition of glutathione-S-transferase from Plasmodium yoelii by protoporphyrin IX, cibacron blue and menadione: implications and therapeutic benefits. Parasitol Res 102:805–807
Michele AM, DeWight RW, John AT (1995) Crystal structures of aschistosomal drug and vaccine target: glutathione S-transferase from Schistosoma japonica and its complex with the leading antischistomal drug Praziquantel. J Mol Biol 246:21–27
Harwaldt P, Rahlfs S, Becker K (2002) Glutathione S-transferase of the malarial parasite Plasmodium falciparum: characterization of a potential drug target. Biol Chem 383:821–830
Ahmad R, Srivastava AK (2007) Purification and biochemical characterization of cytosolic glutathione-S-transferase from malarial parasites Plasmodium yoelii. Parasitol Res 100:581–588
Brophy PM, Campbell AM, van Eldik AJ, Teesdale-Spittle PH, Liebau E, Wang MF (2000) Beta-carbonyl substituted glutathione conjugates as inhibitors of O. volvulus GST2. Bioorg Med Chem Lett 10:979–981
Liebau E, Wildenburg G, Brophy PM, Walter RD, Henkle-Duhrsen K (1996) Biochemical analysis, gene structure and localization of the 24 kDa glutathione S-transferase from Onchocerca volvulus. Mol Biochem Parasitol 80:27–39
Rao UR, Salinas G, Mehta K, Klei TR (2000) Identification and localization of glutathione S-transferase as a potential target enzyme in Brugia species. Parasitol Res 86:908–915
Zhao TJ, Singhal SS, Piper TJ, Cheng JZ, Pandya U, Clark-wronski J, Awasthi S, Awasthi YC (1999) The role of human glutathione-S-transferases hGSTA1-1 and hGSTA2-2 in protection against oxidative stress. Arch Biochem Biophys 367:216–224
Lüersen K, Walter RD, Müller S (1998) The putative γ - glutamylcysteine synthetase from Plasmodium falciparum contains large insertions and a variable tandem repeat. Mol Biochem Parasitol 98:131–142
Bhargavi R, Vishwakarma S, Murty US (2005) Modeling analysis of GST (glutathione-S-transferases) from Wuchereria bancrofti and Brugia malayi. Bioinform 1:25–27
Jones DT (1999) GenTHREADER: an efficient and reliable protein folds recognition method for genomic sequences. J Mol Biol 287:797–815
Okunade AL (2002) Ageratum conyzoides L. (Asteraceae). Fitoterapia 73:1–16
Kaefer CM, Milner JA (2008) The role of herbs and spices in cancer prevention. J Nutr Biochem 19:347–361
Luthra PM, Singh R, Chandra R (2001) Therapeutic uses of Curcuma longa (Turmeric). Indian J Clin Biochem 16:153–160
Filimonov DA, Poroikov VV (1996) PASS: Computerized prediction of biological activity spectra for chemical substances. Bioactive Compound Design. Possibilities for Industrial Use. BIOS Scientific, Oxford, pp 47–56
Lee Sung Kwang (2005) In silico high-throughput screening for ADME/Tox properties: PreADMET program. Abstr Conf Comb Chem Jpn 21:22–28
Schönberger H, Schwab CH, Hirsch AJ, Gasteiger J (2000) Molecular modelling of fullerene dendrimers. J Mol Model 6:379–395
Sadowski J, Gasteiger J, Klebe G (1994) Comparison of automatic three-dimensional model builders using 639 X-ray structures. J Chem Inf Comput Sci 34:1000–1008
Peng C, Ayali PY, Schlegel HB, Frisch MJ (1995) Using redundant internal coordinates to optimize equilibrium geometries and transition states. J Comput Chem 16:49–51
Hendlich M, Rippmann F, Barnickel G (1997) Mapping of protein surface cavities and prediction of enzyme class by a self-organizing neural network. J Mol Graph 15(359–363):389
Thomsen R, Christensen MH (2006) MolDock: a new technique for high-accuracy molecular docking. J Med Chem 49:3315–3321
Korb O, Stutzle T, Exner TE (2009) Empirical scoring functions for advanced protein-ligand docking with PLANTS. J Chem Inf Model 49:84–96
Habig WH, Pabst MJ, Jakoby WB (1974) Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J Biol Chem 249:7130–7139
Gupta S, Bhandari YP, Reddy MV, Harinath BC, Rathaur S (2005) Setaria cervi: immunoprophylactic potential of glutathione-S-transferase against filarial parasite Brugia malayi. Exp Parasitol 109:252–255
Morrison CA, Colin T, Sexton JL, Bowen F, Wicker J, Friedel T, Spithill TW (1996) Protection of cattle against Fasciola hepatica infection by vaccination with glutathione S-transferase. Vaccine 14:1603–1612
Grezel D, Capron M, Grzych JM, Fontaine J, Lecocq JP, Capron A (1993) Protective immunity induced in rat schistosomiasis by a single dose of the Sm28 GST recombinant antigen: effector mechanisms involving IgE and IgA antibodies. Eur J Immunol 23:454–460
Sexton JL, Milner AR, Panaccio M, Waddington J, Wijffels G, Chandler D, Thompson C, Wilson L, Spithill TW, Mitchell GF, Campbell NJ (1990) Glutathione S-transferase: novel vaccine against Fasciola hepatica infection in sheep. J Immunol 145:3905–3910
Veerapathran A, Dakshinamoorthy G, Gnanasekar M, Reddy MVR, Kalyanasundaram R (2009) Evaluation of Wuchereria bancrofti GST as a vaccine candidate for lymphatic filariasis. PLoS Negl Trop Dis 3:e457
Kawamori T, Lubet R, Steele VE, Kelloff GJ, Kaskey RB, Rao CV, Reddy BS (1999) Chemopreventive effect of curcumin, a naturally occurring anti-inflammatory agent, during the promotion/progression stages of colon cancer. Cancer Res 59:597–601
Oetari S, Sudibyo M, Commandeur JNM, Samhoedi R, Vermeulen NPE (1996) Effects of curcumin on cytochrome P450 and glutathione S-transferase activities in rat liver. Biochem Pharmacol 51:39–45
Awasthi S, Pandya U, Singhal SS, Lin JT, Thiviyanathan V, Seifert WE Jr, Awasthi YC, Ansari GAS (2000) Curcumin–glutathione interactions and the role of human glutathione S-transferase P1-1. Chem Biol Interact 128:19–38
Deng XK, Yin W, Li WD, Yin FZ, Lu XY, Zhang XC, Hua ZC, Cai BC (2006) The anti-tumor effects of alkaloids from the seeds of Strychnos nux-vomica on HepG2 cells and its possible mechanism. J Ethnopharmacol 106:179–186
Peana AT, D'aquila PS, Panin F, Serra G, Pippia P, Moretti MDL (2002) Antiinflammatory activity of linalool and linalyl acetate constituents of essential oils. Phytomedicine 9:721–726
Reen RK, Jamwal DS, Taneja SC, Koul JL, Dubey RK, Wiebel FJ, Singh J (1993) Impairment of UDP-glucose dehydrogenase and glucuronidation activities in liver and small intestine of rat and guinea pig in vitro by piperine. Biochem Pharmacol 46:229–238
Singh J, Dubey RK, Atal CK (1986) Piperine-mediated inhibition of glucuronidation activity in isolated epithelial cells of the guinea-pig small intestine: evidence that piperine lowers the endogeneous UDP-glucuronic acid content. J Pharmacol Exp Ther 236:488–489
Shoba G, Joy D, Joseph T, Majeed M, Rajendran R, Srinivas PS (1998) Influence of piperine on the pharmacokinetics of curcumin in animals and human volunteers. Planta Med 64:353–356
Acknowledgments
We acknowledge with thanks the facilities provided by Director, Indian Institute of Spices Research (IISR), Calicut; Dr. Santhosh J. Eapen, Co-ordinator, Distributed Information Sub-Centre (DBT, New Delhi), of IISR (Indian Institute of Spices Research), Dr. O.K. Sindu, District Veterinary Centre Campus, Calicut, who supplied the filarial nematodes Dirofilaria immitis and the assistance rendered by Ms. M. Dinsha, Senior Research Fellow, IISR.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Azeez, S., Babu, R.O., Aykkal, R. et al. Virtual screening and in vitro assay of potential drug like inhibitors from spices against glutathione-S-transferase of filarial nematodes. J Mol Model 18, 151–163 (2012). https://doi.org/10.1007/s00894-011-1035-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00894-011-1035-2