
Abstract
The complete nucleotide sequence of Alternaria longipes dsRNA virus 1 (AlRV1), a novel double-stranded RNA (dsRNA) mycovirus, was determined and analyzed. AlRV1-HN28 contains a single dsRNA genome segment 3415 base pairs in length (excluding the 3′ poly(A) tail) and was predicted to contain two discontiguous open reading frames (ORFs, ORF A and ORF B). The 5′-proximal ORF A (1182 nt) potentially encodes a protein of 394 amino acids (aa) with a predicted molecular mass of 43 kDa; this protein showed no significant similarities to any other sequences in any of the NCBI protein databases. The 3′-proximal ORF B (1737 nt) encodes a protein of 579 aa with a predicted molecular mass of 65 kDa; this protein sequence shares similarities with the conserved domains of RNA-dependent RNA polymerases of other mycoviruses. Phylogenetic analysis indicated that AlRV1-HN28 was closely related to four other unclassified viruses, which suggests that the AlRV1-HN28 isolated from Alternaria longipes may belong to a new family of dsRNA mycoviruses. This is the first report of the full-length nucleotide sequence of a mycovirus that infects Alternaria longipes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Fungal viruses (mycoviruses) are viruses that selectively infect fungi and can target all major fungal groups. Many mycoviruses contain double-stranded (ds) RNA genomes, whereas some have plus-sense single-stranded (ss) RNA genomes. Viruses with dsRNA genomes are grouped into the following five families: Reoviridae, Totiviridae, Partitiviridae, Chrysoviridae and Endornaviridae. Of these, only members of the family Totiviridae contain a single dsRNA genome segment 4.6-7.0 kb in length. Mycoviruses with ssRNA genomes are grouped into the following five families: Hypoviridae, Narnaviridae, Barnaviridae, Alphaflexiviridae (genera Botrexvirus and Sclerodarnavirus) and Gammaflexiviridae [3]. So far, the Sclerotinia sclerotiorum hypovirulence-associated DNA virus 1 (SsHADV-1) is the only mycovirus with a DNA genome [15]. In recent years, some novel mycoviruses have been identified, some of which are able to associate with plant viruses. The Fusarium graminearum virus DK21 and Botrytis virus X resemble positive-strand RNA plant viruses [2], whereas the S. sclerotiorum RNA virus L is related to animal pathogenic viruses [4]. The majority of the reported mycoviruses have no pathogenic effect on the host, including the Fusarium graminearum-associated dsRNA virus FgHV1 [12], but several viruses can cause severe symptoms. Cryphonectria hypovirus 1 (CHV1), which belongs to the family Hypoviridae, is a classic example of a virus that can be used successfully for the biological control of chestnut blight in Europe [6].
Alternaria is a genus of ubiquitous fungi that includes saprobic, endophytic and pathogenic species associated with a wide variety of substrates, and members of this genus are known to be serious plant pathogens, causing major losses on a wide range of crops [14]. Alternaria longipes infects tobacco plants, causing brown spot disease on the leaves. A. longipes is a cosmopolitan species and has been reported in many countries, resulting in significant crop losses [5]. Virus-like particles associated with dsRNAs have been detected in A. alternata isolated from cotton seeds and Japanese pear trees [1, 7]. In 2009, a dsRNA identified as A. alternata virus-1 (AaV-1), which was detected in the EGS 35-193 strain of Alternaria alternata, was described. This virus has four dsRNA genome segments and clusters with Aspergillus mycovirus 341 (AsV341). It was the first mycovirus to be reported on Alternaria with a potential biological effect [1].
In the present study, dsRNA from the novel mycovirus Alternaria longipes dsRNA virus 1 (AlRV1-HN28) was isolated from the Alternaria longipes strain HN28. The virus has a single dsRNA segment with a distinct 2-ORF genome structure. Phylogenetic analysis of the putative RNA-dependent RNA polymerase (RdRp) indicated that AlRV1-HN28 was closely related to unclassified mycoviruses, whereas differences in its genome organization suggest that it may belong to a new family of mycoviruses.
Provenance of the virus material
The dsRNA virus was purified from Alternaria longipes strain HN28, which was isolated from diseased seeds and glumes from Henan Province in China. The HN28 strain was cultured on potato dextrose agar (PDA) plates overlaid with cellophane membranes for one week at 25 °C in the dark. The mycelium mass was used for dsRNA extraction using the CF-11 cellulose chromatography method [10]. The dsRNAs were treated with S1 nuclease and DNase I to eliminate contaminating single-stranded RNA (ssRNA) and DNA. The dsRNAs were electrophoresed on a 1 % (w/v) agarose gel. The purified dsRNA and random primers (5′-GACGTCCAGATCGCGAATTCNNNNNN-3′) were used to synthesize cDNAs [12]. The resulting cDNAs were amplified using a single specific primer (5′-GACGTCCAGATCGCGAATTC-3′) and PrimeSTAR® HS DNA Polymerase (TaKaRa) on a thermal cycler (Bio-Rad, Hercules, CA, USA). The RT-PCR products were purified using a gel extraction kit, ligated to the PMD18-T vector and introduced by transformation into Escherichia coli strain DH5α (TaKaRa) for sequencing. Based on the sequences obtained, dsRNA-specific primers were designed and used for RT-PCR. To clone the termini of the dsRNAs, a classic 3′-RACE protocol using an adaptor-linked oligo-dT primer was performed using the 3′-Full RACE Core Set Ver.2.0 (TaKaRa) according to the manufacturer’s instructions. The 3′ RNA-ligase-mediated RACE (RLM-RACE) protocol was performed to determine the 5′-terminal sequences of the coding strand of the dsRNA element [8]. At least four independent overlapping clones were obtained to sequence every base. The resulting sequences were assembled and analyzed using DNAMAN software (Lynnon Biosoft, Quebec, Canada), and the complete nucleotide sequence of the dsRNA was deposited in the GenBank database under the accession number KJ817371. Homology searches were performed using the National Center for Biotechnology Information (NCBI) BLAST program. A phylogenetic tree was constructed using the neighbor-joining method from the MEGA 4.0 program [9].
Sequence properties
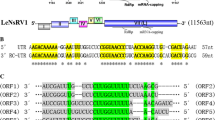
The novel unclassified dsRNA virus Alternaria longipes dsRNA virus 1 (AlRV1), which was isolated from Alternaria longipes strain HN28, has a single genome segment with a nucleotide sequence 3415 bp in length (excluding the 3′ poly(A) tail) (Fig. 1a). Sequence analysis showed that the coding strand of the dsRNA contained two open reading frames (ORFs; ORF A and ORF B) separated by 71 nt. The 5′ untranslated region (UTR) and 3′-UTR of the AlRV1-HN28 were 318 bp and 107 bp long, respectively (Fig. 1b). We also identified two smaller ORFs (300-500 nt in length) in the plus and minus genomic strands. The potentially encoded proteins of these small ORFs did not have any detectable similarity to any proteins in the NCBI database. Thus, these smaller ORF candidates were not examined further.

(a) dsRNA purified from Alternaria longipes was isolated and separated on a 1 % agarose gel. Lane M, DNA marker (5-kb ladder, TaKaRa); lane 1, dsRNA sample after treatment with both RNase-free DNase I and S1 nuclease. (b) Schematic representation of the genomic organization of AlRV1-HN28. The AlRV1-HN28 genome is 3415 bp in length and contains two ORFs (ORF A and ORF B). ORF B encodes the RNA-dependent RNA polymerase (RdRp); the black box in ORF B represents the conserved domain within the RDRP. Thick black lines indicate 5′-, 3′-, and internal UTRs. The poly(A)-tail at the 3′ end of the coding strand is represented as A(n)
Analysis of the genomic organization indicated that ORF A (nt positions 319-1503) was predicted to encode a 394-amino-acid (aa) protein with a molecular mass of 43 kDa. No putative conserved domains were detected in the ORF A protein using the conserved domain search program on the NCBI website, and a BLASTp search using the predicted aa sequence of ORF A showed no significant similarities to any of the sequences present in the NCBI protein databases. ORF B (nt positions 1572-3308) encoded a protein of 579 aa with a predicted molecular mass of 65 kDa. A homology search with the aa sequence encoded by ORF B revealed a low percentage of sequence similarity to the (putative) RdRps of four unclassified dsRNA viruses (Rhizoctonia solani dsRNA virus 1, RsRV1 [17], Fusarium graminearum dsRNA mycovirus-4, FgV-4 [16], Heterobasidium RNA virus 6, HRV6 [11], Cryphonectria parasitica bipartite mycovirus 1, CpRV1), 37 members of the family Partitiviridae (genera Alphapartitivirus, Betapartitivirus, Gammapartitivirus and Deltapartitivirus) [13], two members of the family Astroviridae, and six positive-strand ssRNA viruses of the genus Potyvirus. Furthermore, a search of the conserved domain database (CDD) and multiple protein alignments confirmed that the predicted ORF B protein contained a conserved viral RdRp domain (cd01699, RNA_dep_RNAP) with six conserved motifs that are characteristic of the RdRps of dsRNA mycoviruses.
A phylogenetic tree based on the RdRp sequences of AlRV1-HN28 and 33 selected RNA viruses was generated using the NJ method (Fig. 2). Together with 20 members of the family Partitiviridae and the unclassified dsRNA viruses identified through BLAST searching, an additional 12 dsRNA viruses from the families Totiviridae and Chrysoviridae were subjected to phylogenetic analysis. It was revealed that AlRV1-HN28 formed a clear, well-supported taxonomic cluster together with RsRV1, FgV-4, HRV6 and CpRV1 in the unclassified family, which was grouped separately from the members of the family Partitiviridae. However, the genomic composition of the unclassified family differed according to the species: RsRV1 (2 segments: 1811, 2379 bp), FgV-4 (2 segments: 1739, 2383 bp), CpRV1 (2 segments: 1762, 2026 bp) and HRV6 (non-segmented: 2050 bp). Notably, the genomic composition of AlRV1-HN28 was similar to that of HRV6, which is composed of a single dsRNA 3415 bp in length. Therefore, AlRV1-HN28, RsRV1, FgV-4, HRV6 and CpRV1 might be considered members of a new virus family that can potentially be divided further into two genera.

Unrooted phylogenetic tree constructed by the neighbor-joining method (1,000 bootstrap replicates) using the deduced amino acid sequences of the putative RdRp regions. The scale bar corresponds to a genetic distance of 0.1 amino acid substitutions per site. The abbreviation of virus names and GenBank accession numbers are as follows: GaRV-MS1, Gremmeniella abietina RNA virus MS1(NP_659027.1); PsV-S, Penicillium stoloniferum virus S(YP_052856.2); OPV1, Ophiostoma partitivirus 1(CAJ31886.1); DdV1, Discula destructiva virus 1(NP_116716.1); PsV-F, Penicillium stoloniferum virus F (YP_271922.1); FiCV, fig cryptic virus (CBZ05552.1); PepCV1, pepper cryptic virus 1 (AEJ07890.1); BCV3, beet cryptic virus 3 (AAB27624.1); FpV1, Fusarium poae virus 1 (AAC98734.1); RhsV717, Rhizoctonia solani virus 717 (AAF22160.1); WCCV2, White clover cryptic virus 2 (AGJ83763.1); HetPV1, Heterobasidion partitivirus 1 (ADV15448.1); VCV, Vicia cryptic virus (ABN71234.1); WCCV-1, white clover cryptic virus 1(YP_086754.1); BCV-1, beet cryptic virus 1(YP_002308574.1); CarCV, carrot cryptic virus (ACL93278.1); RsRV1, Rhizoctonia solani dsRNA virus 1(AFZ85210.1); FgV-4, Fusarium graminearum dsRNA mycovirus-4 (YP_003288790.1); HRV6, Heterobasidium RNA virus 6 (AHA82556.1); CpRV1, Cryphonectria parasitica bipartite mycovirus 1(YP_007985675.1); ScVL-A, Saccharomyces cerevisiae virus L-A (AAA50508.1); TaV1, Tuber aestivum virus 1 (ADQ54106.1); ScVL-BC, Saccharomyces cerevisiae virus L-BC (AAB02146.1); SsRV2, Sphaeropsis sapinea RNA virus 2 (NP_047560.1); LRV2-1, Leishmania RNA virus 2-1 (AAB50031.1); LRV1-1, Leishmania RNA virus 1-1 (NP_041191.1); PcV, Penicillium chrysogenum virus (YP_392482.1); AbV1, Agaricus bisporus virus 1(CAA64144.1); FgV-China9, Fusarium graminearum mycovirus-China 9 (ADU54123.1); AMV1816, Aspergillus mycovirus 1816 (EU289896.1); MoCV-1, Magnaporthe oryzae chrysovirus 1 (YP_003858286.1); TcV2, Tolypocladium cylindrosporum virus 2 (CBY84993.1)
References
Aoki N, Moriyama H, Kodama M, Arie T, Teraoka T, Fukuhara T (2009) A novel mycovirus associated with four double-stranded RNAs affects host fungal growth in Alternaria alternata. Virus Res 140:179–187
Howitt RLJ, Beever RE, Pearson MN, Forster RLS (2006) Genome characterization of a flexuous rod-shaped mycovirus, Botrytis virus X, reveals high amino acid identity to genes from plant ‘potex-like’viruses. Arch Virol 151:563–579
King AMQ, Adams MJ, Carstens EB, Lefkowitz EJ (eds) (2011) Virus taxonomy: ninth report of the International Committee on Taxonomy of Viruses. Elsevier Academic Press, London, p 1248
Liu HQ, Fu YP, Jiang DH, Li GQ, Xie J, Peng YL, XH Y, Ghabrial SA (2009) A novel mycovirus that is related to the human pathogen hepatitis E virus and rubi-like viruses. J Virol 83:1981–1991
Luo YY, Yang JK, Zhu ML, Liu CJ, Li HY, Lu ZB, Pan WZ, Zhang ZH, Bi W, Zhang KQ (2012) The group III two-component histidine kinase AlHK1 is involved in fungicides resistance, osmosensitivity, spore production and impacts negatively pathogenicity in Alternaria longipes. Curr Microbiol 64:449–456
Nuss DL (2005) Hypovirulence: mycoviruses at the fungal-plant interface. Nat Rev Microbiol 3:632–642
Shepherd HS (1988) Virus-like particles in tentoxin-production strains of Alternaria alternata. J Virol 62:3888–3891
Suzuki NS, Supyani K, Maruyama, Hillman BI (2004) Complete genome sequence of Mycoreovirus 1/Cp9B21, a member of a new genus within the family Reoviridae, from the chestnut blight fungus Cryphonectria parasitica. J Gen Virol 85:3437–3448
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28:2731–2739
Valverde RA, Nameth ST, JORDAN RL (1990) Analysis of double-stranded RNA for plant virus diagnosis. Plant Dis 74:255–258
Vainio EJ, Hyder R, Aday G, Hansen E, Piri T, Doğmuş-Lehtijärvi T, Lehtijärvi A, Korhonen K, Hantula J (2012) Population structure of a novel putative mycovirus infecting the conifer root-rot fungus Heterobasidion annosum sensu lato. Virology 422:366–376
Wang S, Kondo H, Liu L, Guo L, Qiu D (2013) A novel virus in the family Hypoviridae from the plant pathogenic fungus Fusarium graminearum. Virus Res 174:69–77
Woudenberg JHC, Groenewald JZ, Binder M, Crous PW (2013) Alternaria redefined. Stud Mycol 75:171–212
Nibert ML, Ghabrial SA, Maiss E, Lesker T (eds) (2013) A taxonomic proposal of the International Committee on Taxonomy of Viruses: Reorganization of family Partitiviridae to contain 4 new genera of plant and/or fungal viruses, with elimination of 3 current genera and creation of 16 new species
Yu X, Li B, Fu Y, Jiang D, Ghabrial SA, Li G, Peng Y, Xie J, Cheng J, Huang J, Yi X (2010) A geminivirus-related DNA mycovirus that confers hypovirulence to a plant pathogenic fungus. Proc Natl Acad Sci 107:8387–8392
Yu J, Kwon SJ, Lee KM, Son M, Kim KH (2009) Complete nucleotide sequence of double-stranded RNA viruses from Fusarium graminearum strain DK3. Arch Virol 154:1855–1858
Zheng L, Liu H, Zhang M, Cao X, Zhou E (2013) The complete genomic sequence of a novel mycovirus from Rhizoctonia solani AG-1 IA strain B275. Arch Virol 158:1609–1612
Acknowledgments
This work was supported by the National Natural Science Foundation of China (31171818). We greatly thank Dr. Meng Zhang at Henan Agricultural University, Zhengzhou, Henan, China, for assistance with fungal identification.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article

Cite this article
Lin, Y., Zhang, H., Zhao, C. et al. The complete genome sequence of a novel mycovirus from Alternaria longipes strain HN28. Arch Virol 160, 577–580 (2015). https://doi.org/10.1007/s00705-014-2218-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00705-014-2218-9