
Abstract
Introduction
This experimental study in pigs was aimed at evaluating spinal growth disorders after partial arrest of the vertebral epiphyseal plates (EP) and neurocentral cartilages (NCC). Unilateral and multisegmental single or combined lesions of the physeal structures were performed by electrocogulation throughout a video-assisted thoracoscopical approach.
Materials and methods
Thirty 4-week-old domestic pigs (mean weight 16 kg) were included in the experiments. The superior and inferior epiphyseal plates of T5 to T9 vertebra were damaged in ten animals by hemicircumferential electrocoagulation (group I). In other ten pigs (group II), right NCC at the same T5–T9 levels were damaged. Ten other animals underwent combined lesions of the ipsilateral hemiepiphyseal plates and NCC at the T5–T9 levels. A total of 26 animals could be evaluated after 12 weeks of follow-up using conventional X-rays, CT scans and histology.
Results
The pigs with hemicircumferential EP damage developed very slight concave non-structured scoliotic deformities without vertebral rotation.(mean 12° Cobb; range10–16°). Some of the damaged vertebra showed a marked wedgening with unilateral development alteration of the vertebral body, including the adjacent discs The animals with damage of the NCC developed mild scoliotic curves (mean 19° Cobb; range 16–24°) with convexity opposite to the damaged side and loss of physiological kyphosis. The injured segments showed an asymmetric growth with hypoplasia of the pedicle and costovertebral joints at the damaged side. The pigs undergoing combined EP and NCC lesions developed minimal non-structured curves, ranging from 10 to 12° Cobb. In these animals there was a lack of growth of a vertebral hemibody and disc hypoplasia at the damaged segments. Both damage of the NCC and the EP affect the height of the vertebral body. No spinal stenosis was found in any case. In most cases, the adjacent superior and inferior vertebral EP to damaged segments had a compensatory growth that maintained the straight spinal shape.
Conclusions
In summary, unilateral direct lesion of the EP by hemicircumferential thoracoscopic electrocoagulation modifies vertebral growth, but is not able to induce true scoliostic curves in pigs. Only animals with damaged NCC developed mild scoliotic curves of lordotic type. This work rediscovers and emphasizes the decisive role of the neurocentral cartilage in the ethiopatogeny of idiopathic scoliosis.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Spinal growth modulation has gained credibility as a nonfusion alternative for treatment of idiopathic scoliosis. Morphologic studies of scoliotic vertebrae have demonstrated asymmetric development of the vertebral growth cartilages, that is, the upper and lower epiphyseal plates (EP) and the neurocentral cartilage (NCC). The reason for such asymmetric physeal growth, which is in turn responsible for the vertebral wedging and rotation, has been found to be multifactorial.
In order to guide vertebral growth and to compensate for the tendency toward progression of the scoliotic deformity, several techniques have been used to perform physiodesis of the vertebral growth cartilage. Spinal deformities have been experimentally induced by physeal stapling [1], theatering a flexible cable to the vertebral bodies [2] and by posterior transpedicular screws [3–5]. Most of these techniques involve the destruction of spinal structures adjacent to growth plates, which also might influence the development of the scoliotic curvature.
The effect of isolated or combined unilateral multisegmentary damage of the epiphyseal plate (EP) and the neurocentral cartilage (NCC) on the vertebral growth was experimentally analysed in a pig model. The purpose of this study was to assess the development of spinal deformity after direct injury to the growth vertebral cartilages. Using a minimally invasive approach in combination with video-assisted thoracoscopy, a hemicircumferential veretebral physiodesis in EPs and/or NCC was performed on a pig animal model. Animals were skeletally immature with their vertebral growth potential still active.
Materials and methods
The local ethics committee for experimental animal research approved this study. Thirty 6- to 8-week-old domestic pigs with a mean weight of 16 kg were included in the experiment. Prior to the start of the surgical procedures, one animal not included in the series was used for a close anatomic study on the morphology and architecture of the epiphyseal and neurocentral vertebral cartilages of the pig. The objective of this preliminary study was to set up the surgical techniques trying to avoid injury to neural structures and provide a reference for the identification of cartilages through video-assisted thoracic endoscopy. Anatomical specimens from T5 to T9 vertrebrae were extracted and some measurements were taken: the height of the vertebral bodies and the distance from the epiphyseal and neurocentral cartilage to the spinal canal.
Once the vertebral anatomy was studied, the animals were divided in three groups according to the cartilages to be damaged. Physeal blocking was performed by direct injury of the EP and/or the NCC by electrocautery using video-assisted right thoracoscopy. In the first group of ten animals, the superior and inferior epiphyseal plates of T5 to T9 vertebra were damaged by right hemicircumferential electrocoagulation. In other ten pigs, right NCC was damaged by electrocoagulation at the same T5–T9 levels. The other ten animals of the third group underwent combined lesions of the ipsilateral hemiepiphyseal plates and NCC.
Surgical technique and post-operative care
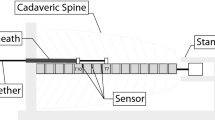
We used an appropriate anaesthetic protocol to reduce morbidity, prevent interactions with the experimental procedure, reduce the degree of suffering, optimise performance and ensure the quality of the research. The animals were positioned such that the left lateral chest was super-elevated, so that thoracoscopy could proceed through the right hemithorax.
Six trocars were inserted through portals: three in the posterior axillary line on ribs 6, 8 and 10 (supradiaphragmatic) and three in the anterior axillary line on ribs 5, 7 and 9. After sectioning the thoracic parietal pleura, the diaphragm was separated at the costal junction, carefully avoiding penetration into the peritoneal cavity. The anterior thoracic spine was approached, between T5 and T9, at the joint connecting the right rib and vertebral bodies.
Hemivertebral cauterisation was performed by electrocoagulation of the epiphyseal, superior and inferior and/or neurocentral cartilage of vertebrae T5 to T9, according to the established groups.
After surgery, the tubes and instruments were removed in reverse order of their introduction, always under camera view. Fluids were aspirated and the absence of postoperative pneumothorax was verified avoiding the placement of a chest tube. The parietal pleura was closed with absorbable 1/0 suture and in the skin with 3/0 silk suture. After closure, postoperative wounds were covered with Betadine cream to promote asepsis.
To achieve postoperative analgesia, fentanyl transdermal patches (Durogensic® 0.5 mcg) were used. Postoperative antibiotic coverage was achieved with 3 mg/kg ceftiofur hydrochloride administered intramuscularly twice daily for 3 days. Pigs were maintained in appropriate housing for 3 months.
Radiographic, morphologic and histologic study
Conventional X-rays—postero-anterior and lateral views—were used to assess vertebral growth 12 weeks after surgery. After animal’s euthanasia, the entire spine between T1 and L5, including the costal arches (sectioned at the mid-axillary line) was extracted for morphological and histological examinations. The extracted costovertebral pieces were also studied by CT scans. Computed axial tomography was carried out with a high-speed scanner (Somatom definition-Siemens), with a scanning power of 200 mA, 120 kV and a slice thickness of 1.4 mm. Two-dimensional reconstruction was performed in the coronal, sagittal and transverse planes for each surgical specimen. Three-dimensional spatial reconstruction was performed as well. In each vertebra with physiodesis of the EP, 3-mm-thick scans were obtained. Transversal cuts that were 3-mm-thick were obtained from physiodesis of the NCC as well. The development of scoliotic curves in the frontal plane as well as in the sagittal plane was evaluated in each animal. In the radiological study with or without computed tomography, deformities were analysed with the following parameters: magnitude of the curvature according to Cobb angle in coronal and sagittal planes; degree of vertebral wedging and height of the vertebral bodies on the injured and uninjured sides; vertebral rotation according to Ho’s method for computerised tomography; width and length of the pedicles in the section of each mediotransverse vertebra injured; and developmental abnormalities of the spinal canal.
Histologic examination of surgical specimens was performed following standard procedures for bone decalcification. The stains used were Masson trichrome and safranin-O. The microscopic study was performed with a DFC420 Leica microscope, with objectives of 2×, 4×, 10× and 20×. The parameters analysed included the study of condrogenic activity of the remaining epiphyseal plate and neurocentral cartilage.
Statistical analysis
Data were processed with SPSS for Windows version 14. The parameters analysed and compared among the three groups were vertebral wedging, vertebral body height of the uninjured side, height of the vertebral body on the injured side, vertebral rotation in the axial plane and pedicle width, because length could not be measured due to the alteration of this parameter in vertebral bodies. Within the first group, a comparative study was performed depending on the amount of EP damage: 0, 1, or 2 areas of the physeal cartilage. The first category included cases where destruction was less than 75% of the physeal cartilage.
Numerical variables were expressed as median values (25th–75th percentiles). Nonparametric tests were analysed with the Kruskal–Wallis and Mann–Whitney tests. Statistical significance was defined as p < 0.05. Comparisons of pedicle width were performed using Student’s t test with independent measures.
Results
A total of 26 animals could be evaluated after 12 weeks of follow-up. None of them displayed neurological deficits at the end of follow-up. There were four animals that could not complete the entire post-operative evaluation period. One animal belonging to group I developed an infection in one of the vertebrae operated upon and had to be killed at 6 weeks post-intervention. In group II, that is pigs undergoing NCC lesions, there were two animals that died before completing the study. One died in the immediate postoperative period due to hemo-pneumothorax related to surgery and one animal died on the fourth week after surgery due to unknown reasons. One animal of the group III died during surgery after developing malignant hyperthermia due to the inability to tolerate anaesthetics.
Epiphyseal plate damage (group I)
The nine pigs with hemicircumferential EP injury developed very slight scoliotic deformities, (mean 12.4° Cobb; range 10–16°). Curves were unstructured with concavity at the injured side. In lateral views, no deformities of interest were observed, with animals maintaining a straight spinal structure. One case presented severe kyphosis at one of the injured levels. Due to its gross deformity during growth, a radiological study was conducted at 4 weeks, revealing kyphosis of 30° that increased to 48° at 12 weeks. Vertebral wedging in the coronal plane ranging from 5 to 10° was observed in 62% of the damaged vertebrae. Wedging greater than 10° was observed in 11% of these animals, and no wedging was observed in the remaining 27% of animals. Median vertebral wedging was 6.3°. Pronounced wedging, up to 25°, was rare. The height of the vertebral bodies was lower on the injured side (concavity of the curve) than on the uninjured side (convexity of the curve). At the uninjured side the mean vertebral height was 24.8 mm (range 23–26 mm) and at the injured side it was 21.1 mm (range 14–25 mm). Some injured vertebral bodies showed a decrease in anterior vertebral height, maintaining the same vertebral height in the posterior half. CT scans revealed no rotation of the apical vertebra (Fig. 1a). No significant changes in the width of the vertebral pedicles were found when the injured side was compared with the non-injured side. Likewise, 3D reconstruction of the injured segment revealed no segmental rotation (Fig. 1b). We did not observe significant stenosis in the spinal canal or deviations of the spinous processes.

a Medial-transverse cut of apical vertebra showing injury of the epiphyseal cartilage without rotation, as determined by Ho’s method; b 3D reconstruction of the injured segment with physiodesis of the epiphyseal cartilage
Upon gross examination, there was a significant lack of formation of vertebral hemibody on the injured side (Fig. 2a). Microscopically, the physeal disruption was unstructured and displayed necrotic material as well as multinucleate cells from foreign body granulomas (Fig. 2b).

a Anatomical specimen with incomplete formation of one-side vertebral hemibody after physiodesis of the epiphyseal growing plates; b Masson trichrome stain displaying injury of the epiphyseal cartilages
Neurocentral cartilage damage (group II)
The eight animals with NCC injuries developed mild scoliotic curves with convexity contralateral to the lesions and with average Cobb angle of 19° (range 16–24°). Slight loss of physiological kyphosis was observed in the sagittal plane. Wedging in the coronal plane from 5 to 10° was found in 65% of the injuried vertebrae showing lower height on the operated side. Wedging lower than 5ª was detected in 10% of the injuries spine segments, and greater than 10° was found in other 10% of vertebrae. Average vertebral wedging was 6.7°/maximun 25°). The remaining 15% of cases did not present wedging although the injury of the NCC was present.
The average height of the vertebral bodies on the injured side was 23.6 mm (range 19–27). At the uninjured side, vertebral body height averaged 25.6 mm (range 24–27). Ho’s method in combination with CT scans revealed rotation in 73% of the injured vertebral bodies. The average rotation was 3.19°, with a maximum rotation of 11.6° (Fig. 3).

Medial transverse view of apical vertebrae with vertebral rotation, as determined by Ho’s method
The 27% of the vertebral bodies that did not present rotation had NCC injury, with residual undamaged physeal tissue near the spinal canal. In certain rotated vertebrae with adjacent vertebral bodies (superior or inferior), slight to moderate rotation did not result in rotation of the vertebra, even with injury to much of the neurocentral cartilage. Hypoplastic pedicles were common on the injured side. The vertebral width of the injured hemibody averaged 5.4 mm (range 4–7), while at the undamaged side was 7.4 mm (range 6.5–7.6). A slight narrowing in the spinal hemicanals of the injured side was noted where the spinal cord approached the injury edge, bending toward the concavity. In the rest of the injured vertebrae, no changes of interest in form or size of the spinal canal were detected. No disorganisation of the vertebral posterior arch was observed.
At the macroscopic level, a complete absence of one-third of the injured anterolateral vertebral body was observed (Fig. 4). At the microscopic level, destruction of the entire NCC occurred (Fig. 5). Few cases displayed cartilage in the area closest to the spinal canal that was completely or even partially free of injury. Residual cartilage appeared unstructured, with isolated regions of cartilage formation.

Lack of formation of the anterolateral third of the vertebral body after neurocentral cartilage lesion

Masson trichrome stain. Complete lesion of the neurocentral cartilage
Combined EPO and NCC injuries (group III)
Animals with combined physeal lesions developed slight scoliotic curves, ranging from 10 to 14° Cobb. Fifty-one per cent of injured vertebrae exhibited wedging in the coronal plane, ranging between 5–10°; 6.7% presented wedging over 10°, being the maximun 28°. These cases did not display significant axial deformity of the spine as an adaptation to offset the remaining upper and lower vertebral bodies. Overall average wedging was 4.8°. The remaining 33.3% did not exhibit any wedging.
Vertebral body height was reduced on the operated-on as compared with the uninjured side. At the uninjured side body height was lower than the other two groups, with an average of 23.1 mm (range 20–25 mm). Mean height of the injured side was 21.7 mm (range 17–25). In 57.7% of the injured vertebrae, minimum rotations were observed (average 1.5°). The maximum rotation (10.8°) was achieved in a vertebra with significant destruction of a hemibody, but without progressive rotation of adjacent vertebrae (Fig. 6).

Axial CT scan after neurocentral and epiphyseal cartilage injury
The pedicles were more hypoplastic on the injured side and less wide. The average pedicle width of the injured side was 4.8 mm (range 4–6.2), and 7.1 mm (range 6.9–7.4) at the non-injured side. Neither deviations of the spinous processes, nor alterations in the formation of the posterior arch, or narrowing of the spinal canal was found in the damaged spinal segments. At the macroscopic level, the absence of formation of the vertebral hemibodies of the injured side was very pronounced (Fig. 7). Microscopic examination showed that empty vertebral space was covered by fibrous scar tissue.

Gross lesions of the injured part with hemiphysiodesis of neurocentral and epiphyseal cartilage. Lack of tissue formation in the injured hemivertebra
Intergroup comparisons
The average coronal Cobb angle in group I was 12.4°; in group II 19°, and 13.1° for group III. Therefore, curves resulted from extensive isolated injury of the NCC injury were more severe than those induced by EP damage and combined PE and NCC injury (ANOVA, p < 0.05). A lesser vertebral wedging was observed in group III (4.8°) as compared with group I (6.3°) and group II (6.7°). Statistically significant differences in the degree of wedging were only found between group II and group III (p < 0.05). The height of the vertebral body on the injured side was lower than that on the non-injured side in all three groups. A statistically significant difference was found between groups I–II, I–III and II–III (p < 0.001).
After analyzing vertebral wedging and height in relation to the extent of vertebral EP destruction in group I, significant differences (p < 0.001) were seen between the injured and uninjured side. Vertebral wedging increased since more physeal cartilage was destroyed (p < 0.001). The height of the vertebral body on the injured side decreases as the extent of cartilage destruction increases (p < 0.001). However, vertebral height of the uninjured was not correlated with the extent of physeal cartilage destruction (p = 0.966).
As to vertebral rotation, it was not found in cases where EP injury was performed. When damage of the NCC was accomplished the average rotation of the vertebrae was higher in group II (3.2°) with isolated NCC lesion than in group III (1.5°) with combined injury of the EP and NCC; this difference was statistically significant (p < 0.005). Finally, the width of the pedicles was compared between the injured and uninjured sides in these last groups II and III. In both groups, the width of the pedicle was smaller on the injured side (p < 0.001).
Discussion
Although an asymmetric growth pattern of the vertebral end plates has been implicated in the development of idiopathic adolescent scoliosis, its role remains controversial. Even more, there is still disagreement in the literature regarding the time-point for closure of the vertebral growth plates [6]. Most studies on spinal growth modulation have proposed to slow down growth on the convex side in order to compensate for the deformity. Both Roaf and Piggot [7, 8] reported that the effectiveness of physeal stapling in the spine was limited, as the biomechanical properties of vertebral EP differ greatly from those of the growing ends of long bones. In fact, using staples, Roaf [7] caused a true spinal fusion and not a pure hemiphysiodesis because his technique involved partial destruction of the physeal plate, intervertebral disc excision and bone graft interposition, resulting in vertebral hemiartrodesis.
More recently, Barrios et al. [9] attempted to induce spinal deformities in the antero-posterior and sagittal planes in the immature pig by vertebral physiodesis using staples. The authors inserted three mediothoracic intervertebral staples through a minithoracotomy involving the intervertebral disc and EP of two adjacent vertebrae. Only some of the animals developed mild scoliosis (not exceeding 14°) of hyperlordotic-type scoliosis (<10°), representing the failure of clip-mediated vertebral fixation. Staples resulted in centrifugal migration by the forces of distraction during vertebral growth, thus rendering physiodesis ineffective.
Similarly, Wall et al. [1] implanted staples using thoracoscopy in seven mediothoracic vertebrae in immature pigs. Each assembly covered the intervertebral disc and the superior physeal cartilage of the lower vertebra, as well as the inferior physeal cartilage of the upper vertebra. The staples were fixed in the vertebrae with screws. Animals developed a significant increase in the curvature in the coronal plane (up to 22.4°), with convexity on the side opposite to the implants. In the sagittal plane, there was an increase in of the physiologic kyphotic curvature. The authors proposed the procedure to reduce or perhaps correct the deformity at early stages in order to avoid instrumentation with rods or fusion.
Experimental spinal growth modulation has been also attempted by implanting a fixed flexible cable, placed anterolaterally to the vertebral bodies of sheep’s thoracic spines [2]. As the authors mentioned, this technique allows manipulating vertebral growth, altering the morphology of the disc and subsequently the alignment of the spine. Segments connected by a flexible cable showed scoliotic deformity (11.6°) and an average increase of 5.1° in thoracic kyphosis. Apparently, 60% of these changes resulted from elastic deformation of the disc. After removing the flexible cable 3 months after implantation, the mobility of the disc in flexi-extension, lateralisation and rotation was not affected. However, the mobility of the involved segment was found to be asymmetric in lateralisation manoeuvres toward the concave side that supported the cable and toward the convexity of the curve.
The role of the NCC on the pathogeny of scoliosis has been newly investigated in a growing pig model. Zhang and Sucato [5] hypothesize that symmetrical growth of the left and right sides of the neurocentral cartilage causes abnormal spinal growth leading to scoliotic deformity. The authors proposed that fusion of the neurocentral synchondrosis might cause asymmetric spine segmental growth inducing vertebral rotation, ultimately resulting in spinal deformity. So, this research group performed asymmetric physiodesis of the NCC by putting one or two transpedicular screws through the neurocentral synchondrosis. A group of animals undergoing transpedicular fixation from T7 to T14 with one screw was compared with another group that underwent fixation with two screws at the same levels [5]. A direct correlation between the extension of epiphysiodesis and severity of the induced scoliosis was found. Fixation with two screws provided greater closure of the synchondrosis and developed curves of up to 42° Cobb with an average of 15° of axial rotation.
This issue had already been studied in 1980, when Beguiristáin et al. [3] performed a selective unilateral physiodesis by inserting a screw through the neurocentral cartilage on eight pigs at five thoracic levels, obtaining immature scoliotic curves of 30° with convexity towards the operated side. Studies such as those by Beguiristáin et al. and Zhang and Sucato [3, 5] emphasised NCC asymmetry as a cause of structured scoliosis. However, other studies demonstrated the effects of EP on vertebral wedging which in turn develop scoliotic deformity [1, 2]. The results from these studies prompted us to analyse the effectiveness of physiodesis of EP and/or NCC by direct electrocoagulation decreasing the possibilities of surgical complications due to the potential extrusion of the implants. Furthermore, our approach permits evaluating the development of spinal deformities by complete destruction of vertebral growth cartilages, using a minimally invasive thoracoscopic approach.
In the current experimental work, less severe curvatures were obtained by unilateral complete physiodesis of the NCC, as compared with the double-screw physiodesis, with an average of 19° in our group compared with 40° in the Zhang and Sucato’s study [6]. These results were obtained using double screw to achieve synchondrosis closure of 100% in the centre of the vertebra, 75% at the top and 50% at the bottom of the vertebra. Destruction of the NCC with electrocautery was complete in most of the treated vertebrae and was associated with a lack of vertebral hemibody development. Rotation in the two studies occurred on the side of the physiodesis. These results support the theory of spine rotation to the side of the convexity due to cartilage growth on the uninjured side, in combination with the cessation of activity on the injured NCC side. In fact, in Zhang and Sucato’s study [5], the curve concavity was towards the opposite side of the NNC physiodesis. On the contrary, in our experiment, curve concavity following NCC destruction by electrocoagulation was on the side of the injured physis. The lower height of the vertebral bodies on the injured side might possibly explain this unexpected finding.
In our animals with unilateral isolated NCC damage the average rotation was also significantly lower than that of the double screws (15.2° compared with 3.2°). It must be borne in mind that the Zhang and Sucato’s animals had a longer follow-up period (6 months) than ours (3 months).
Similar defects in pedicle development were seen in both Zahng and Sucato′s model and our animals, but with reduced vertebral rotation following electrocautery-mediated NCC destruction. This may be because the cessation of activity on the injured side and the growth of the uninjured side did not cause as much rotation to the convex side due to the defective formation of the vertebral body, which was replaced by fibrous and bone tissue.
Regarding wedging, no significant differences were found between that produced by the hemivertebral EP injury and that induced by NCC damage. Selective damage of the NCC secondarily alters epiphyseal cartilages. This may be the reason for similar wedging patterns.
After direct injury of the EP by electrocoagulation, the vertebral body on the uninjured side was 14.7% higher than on the injured side; this percentage is higher than that produced after NCC injury (7.7%). This difference nearly doubled because physeal cartilage affects longitudinal growth of the vertebral body. However, when performing unilateral physiodesis of the NCC, the vertebra longitudinal growth was also influenced presumably because the anatomic cross-linking of EP and NCC. Interestingly, when performing combined injuries of both cartilages, the decrease in vertebral height was almost similar to that found after isolated lesion of the EP. These physeal end plates seem to be more heavily involved in vertical growth that the NCC. As in Zhang and Sucato’s study, it is important not to neglect the contribution of the NCC to the development of vertebral height. In animals with isolated neurocentral cartilage physiodesis using transpedicular screws, wedging and lower heights of the vertebral body on the side with physeal lock were also detected.
The induction of asymmetric growth of the vertebrae, especially in combined EP and NCC lesions, appears to be a potential alternative for a fusionless surgical treatment of scoliosis. This could be especially relevant for cases of congenital scoliosis due to vertebral malformations such as hemivertebrae (lack of formation) or bars (segmentation absence), resulting in an asymmetry of spine growth. In most of these cases, the orthopaedic treatment is not effective except for the handling of compensatory curves. As scoliosis progresses, patients must undergo surgery, even at early ages. In those patients with combined congenital malformations exhibiting poor prognosis, a minimally invasive surgical procedure for epiphyseal thoracoscopic electrocoagulation could be a promising non-fusion alternative.
In summary, direct destruction of the vertebral physis by electrocoagulation via a thoracoscopic approach has proved to alter vertebral growth in pigs. Direct unilateral lesion of epiphyseal cartilages produces unstructured mild scoliotic curves without vertebral rotation. Direct injury to one-side neurocentral cartilage causes mild lordotic-type scoliotic curves, with slight vertebral axial rotation characterised by concavity towards the injured side. Destruction of both epiphyseal and neurocentral cartilages disrupts formation of one-side hemivertebrae, but results in limited vertebral axial rotation. Furthermore, both cartilages influence vertebral body height. The vertebral body height is as shorter as the number of physeal cartilage destroyed increases. Neurocentral cartilage destruction causes hypoplasia of the pedicle but does not cause spinal canal stenosis or morphological changes of the posterior arch. The vertebrae superior and inferior to the injured spinal segment compensate for the deformity produced by damaging the growth plates, maintaining a straight spine structure and avoiding the progression of curvature.
Finally, concerning the potential clinical relevance of this experimental research on pigs, we are aware of the limitations of this study, in the sense that conclusions regarding direct clinical application to human scoliosis cannot be drawn at present. This new concept of vertebral growth modulation using minimally invasive techniques must be regarded as the beginning of a new stage in the attempt to correct scoliosis without permanent arthrodesis.
References
Wall EJ, Bylski-Austrow DI, Kolata RJ, Crawford AH (2005) Endoscopic mechanical spinal hemiepiphysiodesis modifies spine growth. Spine 30:1148–1153
Newton PO, Fricka KB, Lee SS, Farnsworth CL, Cox TG, Mahar AT (2002) Asymmetrical flexible tethering of spine growth in an immature bovine model. Spine 27:689–693
Beguiristáin JL, De Salis J, Oriaifo A, Canadell J (1980) Experimental scoliosis by epiphysiodesis in pigs. Int Orthop 3:317–321
Vital JM, Beguiristain JL, Algara C, Villas C, Lavignolle B, Grenier N, Senegas J (1989) The neurocentral vertebral cartilage: anatomy, physiology and physiopathology. Surg Radiol Anat 11:323–328
Zhang H, Sucato DJ (2008) Unilateral pedicle screw epiphysiodesis of the neurocentral synchondrosis. Production of idiopathic-like scoliosis in an immature animal model. J Bone Joint Surg Am 90:2460–2469
Kouwenhoven JW, Castelein RM (2008) The pathogenesis of adolescent idiopathic scoliosis: review of the literature. Spine 33:2898–2908
Roaf R (1963) The treatment of progressive scoliosis by unilateral growth-arrest. J Bone Joint Surg [Br] 45:637–651
Piggott H (1987) Growth modification in the treatment of scoliosis. Orthopedics 10:945–952
Barrios C, Maruenda JL, Serra I (2005) Fracaso en la inducción de escoliosis experimental mediante hemiepifisiodesis vertebral por minitoracotomía. V Congreso General INVESCOT (Asociación Española de Investigación en Cirugía Ortopédica y Traumatología), Salamanca
Conflict of interest
None.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Caballero, A., Barrios, C., Burgos, J. et al. Vertebral growth modulation by hemicircumferential electrocoagulation: an experimental study in pigs. Eur Spine J 20 (Suppl 3), 367 (2011). https://doi.org/10.1007/s00586-011-1909-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00586-011-1909-0