
Abstract
Arbuscular mycorrhizal fungi (AMF) were surveyed for species richness and abundance in sporulation in six distinct land uses in the western Amazon region of Brazil. Areas included mature pristine forest and sites converted to pasture, crops, agroforestry, young and old secondary forest. A total of 61 AMF morphotypes were recovered and 30% of them could not be identified to known species. Fungal communities were dominated by Glomus species but Acaulospora species produced the most abundant sporulation. Acaulospora gedanensis cf., Acaulospora foveata, Acaulospora spinosa, Acaulospora tuberculata, Glomus corymbiforme, Glomus sp15, Scutellospora pellucida, and Archaeospora trappei sporulated in all land use areas. Total spore numbers were highly variable among land uses. Mean species richness in crop, agroforestry, young and old secondary forest sites was twice that in pristine forest and pasture. fungal communities were dominated in all land use areas except young secondary forest by two or three species which accounted for 48% to 63% of all sporulation. Land uses influenced AMF community in (1) frequency of occurrence of sporulating AMF species, (2) mean species diversity, and (3) relative spore abundance. Conversion of pristine forest into distinct land uses does not appear to reduce AMF diversity. Cultural practices adopted in this region maintain a high diversity of arbuscular mycorrhizal fungi.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The legal Amazon located in the northern region of Brazil covers 60% of the country's territory and embraces a wide variety of edaphic environments and ecosystems composed of distinct types of equatorial and tropical forests, savannahs, tropical grasslands, and flooded lands (BRASIL 2002). Because of its large area and diverse soil types, flora, and fauna, the Amazon represents a major hotspot of global biodiversity (Daly and Prance 1989). Anthropogenic disturbance has caused deforestation of pristine forest that either is degraded or converted to different land uses (LUs) such as domestic orchard, agroforestry, crop fields, or pastures (Mendonça-Santos et al. 2006).
Forest clearing causes loss of biodiversity and modifies soil physical, chemical, and biological properties (Mendonça-Santos et al. 2006). Agriculture and pastures for beef cattle ranching account for more than 90% of the deforestation that occurred in the 1980s (Amelung and Diehl 1992). Despite being highly weathered and nutrient-poor, Amazon soils support a dense evergreen rainforest composed of plants that have evolved distinct mechanisms for nutrient cycling and uptake. Nutrients are sequestered mostly in the vegetation and they are efficiently cycled through forest litter and the top soil layer. Once this vegetation is removed, however, carbon and nutrient cycles are disrupted and soils are exposed to intensive radiation, leaching, and erosion (Mendonça-Santos et al. 2006). Owing to loss of organic matter and the non-replenishment of soil nutrients, these highly weathered soils became unfertile and unproductive, often with high aluminum toxicity and severe phosphorus and nitrogen deficiency (Cochrane and Sanchez 1982). Once cleared, soils became unproductive and are left on fallow by local farmers, spontaneous re-vegetation is allowed to occur. This plant biomass represents a natural resource for the next cropping cycle, which is described as slash-and-burn agriculture.
Soil biota comprises much of the earth's biological stock and diversity and plays crucial roles in ecosystem functioning (Fitter et al. 2005). Amongst the soil functional groups are arbuscular mycorrhizal fungi (AMF) in Glomeromycota, which contribute to plant health and nutrition and are widespread in natural and man-made ecosystems worldwide. AMF have been shown to be beneficial to tropical plant species (Janos 1980; Siqueira et al. 1998; Siqueira and Saggin-Junior 2001; Zangaro et al. 2007; Pasqualini et al. 2007). Their potential influence on soil processes and on plant diversity (Rillig 2004) have increased interest on AMF of tropical forests where deforestation pressure and expansive agriculture may threaten conservation of these mutualists.
Analyses of AMF communities in tropical systems have focused on changes in abundance and richness of species based on sporulation patterns (Johnson and Wedin 1997; Cuenca et al. 1998; Picone 2000; Zhang et al. 2004; Mangan et al. 2004; Guadarrama-Chávez et al. 2007; Violi et al. 2008). Comprehensive studies are difficult because AMF reside only in soil, are obligate biotrophs, and both morphological and molecular identification of species can be problematic. In the Amazon region, little is known about general pattern of AMF occurrence and diversity or the impact of anthropogenic interference on AMF communities. Peña-Venega et al. (2007) recorded 18 AMF species in the Amazon forest of Colombia that were predominantly Glomus and Acaulospora.
This study is part of a broader project entitled “Conservation and Sustainable Management of Below-Ground Biodiversity” (CSM-BGBD), co-funded by the Global Environment Facility with implementation support of the United Nations Environment Programme and developed in Brazil, Mexico, Ivory Coast, Uganda, Kenya, India, and Indonesia. The aims of this project are to increase the general awareness, knowledge, and comprehension of biological diversity of soils important to sustainable agriculture in tropical soils.
In Brazil, the study was carried out at in the Alto Solimões region in the western portion of the Brazilian state of Amazon. This region was chosen because it is composed of a mosaic of pristine forest and secondary forest at distinct stages of regeneration adjacent to small crop areas and managed forest systems (Fidalgo et al. 2005). All of these areas represent land uses by traditional human communities in that region, but access is difficult. Therefore, anthropogenic influence is low compared to other Amazonian regions. In this paper, we report on AMF species richness and abundance based on spore abundance data. We tested the hypothesis that overall AMF community composition will be reduced by distinct land uses following deforestation in the Amazon forest.
Material and methods
Benchmark area
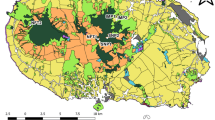
Three sites were studied. They are located in Benjamin Constant municipality (4°21′ and 4°26′ S, 69°36′ and 70°1′ W), in the Alto Solimões river region, western part of the Amazon state, Brazil (Fig. 1), at the border with Peru and Colombia. The climate of the area is tropical humid and super humid (Af type according to Köppen) with no dry season. Average annual temperature is 25.7°C and mean rainfall is 2,562 mm, with the majority of precipitation falling from December to April (Fidalgo et al. 2005). One study site was private land located in the neighborhoods of Benjamin Constant town, and the remaining two sites were located in the rural communities of Nova Aliança and Guanabara II, which are territories occupied by “Cocama” and “Ticuna” cultures (Fig. 1).

Map of Brazil showing the Legal Amazon (gray filled area), Amazon state (AM) indicating the localization of Benjamin Constant in the Alto Solimões region and the three study sites (grids 3–5, grids 1–2, grid 6). Grid 5 illustrates the distribution of sampling points per grid
Sampling design and soil collection
Soils were sampled from six distinct LUs areas categorized as follows: pristine forest, old secondary forest, young secondary forest, agroforestry system, crop, and pasture. Fidalgo et al. (2005) provides a detailed characterization of each land use area as summarized in Table 1. Soils are dominated by low fertility Inceptisols (Coelho et al. 2005) with the following properties: pH 4.5–5.4; P 2.8–4.5 mg dm−3; exchangeable base 3.8–11.7 cmolc dm−3 OM 1.6–1.9 dag kg−1; H+Al 8.2–22.1 cmolc dm−3, soil density 0.82 g cm−3.
Soil sampling was carried out in six grids of 16-points-spaced representing all land use area (Fig. 1). LUs were not evenly distributed in the landscape and therefore grids were set up in order to obtain a significant number of replicates (sampling points) per land use area. Each sampling point was geo-referenced and was distributed in grids to follow the norms of the CSM-BGBD projects for all countries involved (Huising et al. 2008). A wood stick was placed in each sample point as a center reference for all teams participating of the CSM-BGBD project (Research Group BiosBrasil - www.biosbrasil.ufla.br). A total of 98 points were sampled: 80 of them corresponding to the center reference point and 18 of them located between two main center points and 50 m distant. Each sample was composed of 12 subsamples: Four subsamples were collected 3 m from the center point in all cardinal directions. Another eight subsamples were collected 6 m from the center point, with four in cardinal directions and four between them. All subsamples were collected with a soil core sampler at 0–20-cm depth and then pooled. The soil core sampler was flame-sterilized in the field between sampling. Soil from each sample was homogenized in plastic trays, transferred to new plastic bags and transported to the laboratory. All samples were collected in a field campaign during 10–15 March 2004.
AMF spore extraction and species identification
Spores were isolated from a 100 cm3 subsample from each soil sample by wet sieving (Gerdemann and Nicolson 1963) followed by centrifugation in a 20%/60% sucrose gradient. The subsample first was placed in a 2 L bucket with tap water, swirled, and the soil suspension passed through two nested sieves, with 710 and 53 μm openings. Material retained on the 710-μm sieve was transferred to a large Petri dish and inspected under a dissecting microscope (Zeiss Stemi 2000) to collect large AMF spores and sporocarps. The material collected on the 53-μm sieve was poured into a 50-ml Falcon tube containing the sucrose gradient and centrifuged at 1000×g for 1 min. Supernatant was decanted into a 53-μm sieve, washed, transferred to Petri dishes, and spores inspected under a dissecting microscope. All spores were separated by morphotype according to macroscopic features such as size, color, hyphal attachment, and shape. After counting, the spores were mounted permanently on slides in polyvinyl-lacto-glycerol (PVLG) and PVLG mixed with Melzer's reagent (1:1, v/v). Slides were stored at room temperature for 7 days and then at 35°C for 3–4 days. Spores were examined under a compound microscope (Zeiss Axiostar plus) and identified by spore wall structure, Melzer's reaction, and other taxonomically informative characteristics. Phenotypes were compared with those described on original species protologues (Schenck and Perez 1990) and online references of species description of INVAM at the West Virginia University, USA (http://invam.caf.wvu.edu) and Department of Plant Pathology, University of Agriculture in Szczecin, Poland (http://www.agro.ar.szczecin.pl/~jblaszkowski/).
We were conservative in assigning a species name to a morphotype. Minor differences in spore phenotype were merged into one species to minimize inflation of species richness estimates. Reasons for this approach were (1) some AMF spores recovered from the field were in poor conditions or missing informative taxonomic features and (2) the range of morphological variation was ambiguous because of low spore numbers. Sporocarpic species of Glomus were found in many samples and only sporocarp numbers were recorded. Sporocarpic species were included in calculations of frequency of occurrence and species richness, but excluded from calculations of spore abundance.
Measurement of AMF diversity
Differences in AMF communities between LUs were investigated by using a suite of population measurements. AMF abundance (spores per 100 cm3 soil) was estimated by direct counts of field-recovered fungal spores. Abundance was recorded for each species and for each sample by summating counts of all species in a sample. Frequency of occurrence (FO) was calculated by the number of samples from which spores of a particular species was recovered and expressed as a percentage. We applied the approach of Zhang et al. (2004) to classify AMF species as dominant (FO > 50%), most common (30% < FO ≤ 50%), common (10% < FO < 30%), and rare (FO ≤ 10%). To calculate an overall frequency index (FI) for each AMF species, we assigned value of 1, 0.75, 0.5, or 0.25 to a species if it was dominant, most common, common, or rare, respectively. The sum of values over all LUs corresponded to the FI for each species. Therefore, the largest FI was 6.0 for a species dominant in soils at all six LUs. Sporulating AMF species also were ranked as generalists if they were detected in five to six LUs, intermediate if present in two to four LUs and exclusive if detected at only one land use.
Diversity for each LU was calculated using several descriptors according to Magurran (2004) and PAST (PAleontological STatistics) software (Hammer et al. 2001). Number of AMF species (S) was determined by counting fungal species per soil sample; species richness was estimated by the Margalef index (\( {D_{\rm{Mg}}} = S - {1}/{ \ln }\,N \), where N is the total number of AMF spores); Shannon diversity index (\( H\prime = - \sum\nolimits_{\rm{i}} {{{\hbox{p}}_{\rm{i}}}\left( {\log \,{{\hbox{p}}_{\rm{i}}}} \right)} \), where pi is the relative spore abundance of i-th species considering all species) and Shannon evenness were computed for each LU rather than individual soil samples. We generated species accumulation and first-order jackknife curves to estimate the expected number of AMF species in each land use using the software BioDiversity Professional version 2.0.
Statistical analysis
The number of sampling points collected in each land use was treated as replicates in a one-way analyses of variance following Tukey ad hoc test at a P value of <0.05. Spore abundance data were log (x + 1) transformed prior to analysis to meet normality and homogeneity of variance, but values were expressed as number of spores per 100 cm3 soil. Differences on the Shannon diversity of AMF between LU was tested using the statistical procedures for diversity indices in PAST (Hammer et al. 2001).
Results
AMF community composition
A total of 61 morphotypes of AMF were recovered from 98 soil samples collected over all LUs, of which 43 (70%) were identified to known species in the families Acaulosporaceae, Glomeraceae, Gigasporaceae, and Archaeosporaceae (Table 2). In all six LUs, 35 putative species were members of Glomus (Tul. and Tul.) Gerdemann and Trappe, 16 in Acaulospora Gerdemann and Trappe emend. Berch, two in Entrophospora Ames and Schneider, five in Scutellospora Walker and Sanders, one in Gigaspora Gerdemann and Trappe, and two in Archaeospora Morton and Redecker. Glomus and Acaulospora comprised 84% of all species. Eight species were found in all six LUs: Acaulospora gedanensis cf., Acaulospora foveata, Acaulospora spinosa, Acaulospora tuberculata, Glomus corymbiforme, Glomus sp15, Scutellospora pellucida, and Archaeospora trappei. Twenty two species (34.4%) were classified as generalists, 31 (48.4%) as intermediate, and 11 (17.2%) as exclusive (Table 2). Most exclusive species were present either in pasture or pristine forest. A brief morphological description of morphotypes identified to genus level is summarized in Table 3.
Twelve AMF species were classified in three or four frequency of occurrence groups depending on LUs. The most extreme species was G. corymbiforme, whose sporulation pattern was rare in pasture, common in crop, most common in agroforestry, old secondary forest, and pristine forest and dominant in young secondary forest (Table 2). Frequency index (FI) in general was higher for Acaulospora species than for Glomus species, averaging 2.67 and 1.62, respectively. FI was less than 4.0 for A. gedanensis cf., A. foveata, A. delicata, Acaulospora. mellea cf., G. corymbiforme, and Glomus sp15. FI was less than 3.0 for the 49 other species.
Among the 35 Glomus species, six were sporocarpic and formed spores in large numbers (Table 2). These species included Glomus sinuosum, Glomus clavisporum, Glomus coremioides, and Glomus rubiformis. Sporocarps were most abundant in crop (215), young secondary forest (279), and old secondary forest (156). Sporocarpic species were rare in pasture and in pristine forest.
Spore abundance and species richness
Total spore abundance varied markedly amongst LUs, with a 6.3-fold difference between pristine forest and secondary forests/crop. Abundance in pristine forest was very low compared to other sites, reaching a maximum in young secondary forest and crop (Table 4). Mean spore densities also were variable amongst LUs, differences were significant only in a comparison of pristine forest (51 ± 9) system and pasture (255 ± 58) and crop (233 ± 53; Fig. 2). Mean species richness ranged from 6.1 in pristine forest to 12.8 in young secondary forest. On average, species richness was twice as high in crop, agroforestry, young and old secondary forest as in pasture and pristine forest (Fig. 2). Although richness between the two extreme LUs did not differ, spore density was highest in pasture samples measuring 5-fold higher than in undisturbed pristine forest.

Mean spore number (bars) and mean species richness (line) of arbuscular mycorrhizal fungi in pasture (PS), crop (CR), agroforestry (AF), young secondary forest (YS), old secondary forest (OS), and pristine forest (FO) land use categories in the Amazon. Bars with the same letter are not statistically significant (P < 0.05)
In order to understand how LUs affect sporulation of AMF communities, we compared relative abundance of the five most prolific sporulators (Fig. 3). In the pristine forest, two Acaulospora species comprised for 63% of total spores counted. In the other extreme of interference represented by pasture area, the three most abundant sporulating species accounted for 73% of the total spores, each a member of a different genus: Acaulospora, Glomus, and Entrophospora. A. gedanensis cf. was the most prolific sporulator in pasture, agroforesty and young secondary forest and was the second most abundant species in other LUs. Except in young secondary forest, two AMF species accounted for 48% to 63% of sporulation in all LUs. Acaulospora delicata had the largest relative abundance in crop and pristine forest while Glomus glomerulatum was the most abundant species in old secondary forest. Two species commonly found as prolific sporulators were Acaulospora excavata and G. corymbiforme.

Relative abundance (% of the total spores) of AMF species occurring in distinct land use systems in the Amazon
Measures of diversity
The number of species varied twofold among some LUs. The lowest number of species (23) was in pasture. The number of species (50) was highest in young secondary forest followed by 44 species in crops (Table 4). The Shannon index of diversity differed significantly among LUs except for crop and old secondary forest (Table 4). Pristine forest and pasture measured the lowest H′ index while young secondary forest had the highest H′. Margalef's species richness followed the same trend of H′. Evenness values were low for all LUs, ranging from 0.25 in pristine forest to 0.42 in young secondary forest.
Observed species accumulation curves and Jacknife one estimated curves did not plateau in any LUs except Young Secondary Forest (Fig. 4), where the accumulation curve leveled after 25 samples. Based on jackknife one estimates, the expected number of species was 28 in pasture, 47 in crop, 42 in agroforestry, 52 in young secondary forest, 39 in old secondary forest and 38 in pristine forest.

Species accumulation curves (dotted lines) and Jacknife one estimate curves (solid lines) for AMF in different land use systems in the Amazon. Scale of the y axis adjusted to accommodate the estimated curve
Discussion
We assessed AMF spore diversity based on field-collected spores in distinct LUs in a small region in western Amazon that encompassed a pristine forest to old cultivated soils under tropical grass pasture. Evidence from this study suggests that this Amazon region harbors a diverse AMF community relative to that in other tropical ecosystems. AMF community differentiation among LUs was largely detected by changes in the frequency of occurrence of AMF species, in mean species diversity and in the relative abundance of the most abundant species. Differences in total spore abundance were not informative. Our data has some constraints due to: (1) diversity measurements were based only on field-collected spores, (2) samples were collected only at one time, and (3) spore diversity does not necessarily correlate with species diversity. Nevertheless, this is the first study to report AMF species richness and community structure based on an extensive sampling of a region in the Amazon forest.
The total number of AMF species recorded in this study is larger than that reported in previous studies of tropical ecosystems. In rainforests, number of species ranged from 27 species in China (Zhao et al. 2003), 13 in Costa Rica (Lovelock et al. 2003), 24 in Panama (Mangan et al. 2004), 16–25 species in Mexico (Guadarrama and Álvarez-Sánchez 1999; Guadarrama-Chávez et al. 2007). Number of AMF species similar to that found in this study was reported by Zhang et al. (2004) for evergreen broadleaved subtropical forests in China, where 47 fungal species were recognized. Comparisons between studies in tropical regions must be viewed with caution as for constraints listed above. Moreover, AMF species richness in tropical regions does not contrast remarkably from that recorded for temperate regions using more exhaustive approaches (Bever et al. 1996, 2001). The area sampled was large (54 ha) and LUs represented Amazonian environments, harboring a diverse plant community with distinct plant life history traits and soil conditions, factors known to affect AMF composition and abundance (Bever et al. 2001).
Land uses influenced mean species richness and spore abundance (Fig. 2). Pasture was highly disturbed, first with native vegetation completely removed, then soil under cultivation for 45 years with sugarcane, and finally as forage for cattle ranching. In contrast, pristine forest was a stable, undisturbed environment subject only to natural succession. Mean species richness was lowest in pasture and pristine forest, with 7.3 and 6.1 species per sample, respectively. However, plant species numbered 60 in the former and 599 in the latter. This is not in accordance to the general ecological dogma that biodiversity above ground corresponds with that below ground (Wardle et al. 2004). The number of sporulating AMF species is low compared to the large range of associated plant species (this study – tropical region). In contrast, AMF species richness (37) is roughly of the same magnitude as plant richness in temperate regions (Bever et al. 2001). AMF species richness in crops, agroforestry, and young secondary forest ranged from 11.6 to 12.8. These values are high, however, when compared to other tropical soils with species richness varying from 3.5 to 5.5 (Zhao et al. 2003; Mangan et al. 2004; Violi et al. 2008).
We can tentatively explain patterns of sporulation and species richness by evoking two distinct processes. Historical processes related to geograpy of the region and random dispersal pattern of AMF could account for some differences detected as species composition of local communities reflects processes such as vicariance and dispersal (Holt 1983). Local processes impacting on inoculum potential of fungi in each LUs, as affected by plant dependence on mycotrophy and plant density, could also partially explain the patterns observed. For instance, pristine forest is dominated by climax plant species which are known to exhibit low degree of mycotrophy in contrast to pioneers or early successional species from tropical rain forests (Siqueira et al. 1998).
The AMF community recovered from the six Amazonian systems was dominated by Glomus followed by Acaulospora species. This pattern is similar to that observed in other tropical systems, such as forests in China (Zhao et al. 2003; Zhang et al. 2004), Panama (Mangan et al. 2004), Brazil (Zangaro et al. 2007; Stürmer and Siqueira 2006), and Mexico (Guadarrama-Chávez et al. 2007). In contrast, Acaulospora dominated in a tropical forest in Costa Rica (Lovelock et al. 2003) and secondary vegetation in Brazil (Stürmer et al. 2006). Both genera were co-dominants in Costa Rica (Picone 2000; Johnson and Wedin 1997) and in tropical “cerrado” in Brazil (Siqueira et al. 1989). Low numbers of Gigaspora and Scutellospora spores also were found in these regions.
Species of Glomus producing spores in highly organized sporocarps also were common components in all LUs studied except pasture, where only one sporocarpic species was found. The number of sporocarps produced by these species was unexpectedly high. For example, 53 sporocarps of Glomus. rubiformis and 12 sporocarps of Glomus coremioides were collected from a single 100 cm3 soil sample from Young Secondary Forest and Crop, respectively. Sporocarpic Glomus species were recorded in tropical forest soils by Picone (2000), Mangan et al. (2004), and Guadarrama and Álvarez-Sánchez (1999) but not in such high densities. Some species were widely distributed, such as Acaulospora delicata, A. foveata, A. gedanensis cf., A. mellea cf., G. corymbiforme, and Glomus sp15. S. pellucida and Archaeospora trappei also were detected in all LUs as common or rarely occurring, the former species being one of the most frequently recovered in distinct Brazilian ecosystems (Stürmer and Siqueira 2006). Glomus sp15 seems to be very common in tropical forest soils (Stürmer, personal observation) and morphological similar to Glomus brohultii Sieverding and Herrera. This study is the first record of G. corymbiforme, Glomus trimurales cf., Acaulospora paulinae, and Acaulospora polonica in Brazil
Patterns of species richness, sporulation, and frequency of occurrence of Glomus and Acaulospora species indicate that the spatial distribution of both genera over LUs was distinct. Acaulospora species typically do not sporulate prolifically, yet they are commonly widespread (average FI = 2.67). Many Glomus species also produce only low numbers of spores, but they have a more limited distribution (average FI = 1.62). Overall, Acaulospora species accounted for 60% of total spore numbers while Glomus species accounted for only 33% of the total, a trend observed over all LUs, except old secondary forest. One underlying cause of Glomus dominance may be the phylogenetic history of the genus. Glomus is the most speciose taxon, even though it is comprised of several highly supported monophyletic clades (Schwarzott et al. 2001). The high relative abundance of Acaulospora species also detected in Nicaragua and Costa Rica soils (Picone 2000; Lovelock et al. 2003) and Brazilian “cerrado” (Siqueira et al. 1989) can be partly explained by adaptation of local isolates to highly weathered low pH and low fertility soils. Dominance of Acaulospora in tropical soils may be partially controlled by the even distribution of rainfall throughout the year (Lovelock et al. 2003), a condition found in our benchmark area. We cannot attribute differences in distribution and abundance of Acaulospora versus Glomus to preference by the former to slow-growing woody species (Lovelock et al. 2003). The spatial distribution of both genera was similar in pasture with fast growing grass species and in pristine forest and old secondary forest dominated by woody and herbaceous species.
LUs appeared to have some influence on AMF richness and abundance given the identity and the relative abundance of the five most prolific sporulating species. Acaulospora and Glomus species were among the most abundant regardless of LUs. However, E. colombiana and A. trappei dominated in pasture and in old secondary forest, respectively.
AMF communities in our study sites consisted of two or three species producing the majority of spores. This pattern of dominance was observed in distinct Brazilian ecosystems, from agroecosystems to forest (Stürmer and Siqueira 2006) and in tropical forest soils where Acaulospora morrowiae and Acaulospora mellea accounted for 91.5% of total spore numbers (Lovelock et al. 2003).
AMF in tropical soils appear to have resilience when facing shifts in habitats from pristine forest to other LUs. Most fungal species in this study appeared to be generalists (22) or of intermediate adaptability (31) while only 11 sporulated exclusively in one land use area. Johnson and Wedin (1997) and Picone (2000) also observed resilience of AMF in tropical ecosystems. Resilience to surviving drastic management practices may contribute to maintenance of high AMF diversity in Amazonian soils. Pristine forest, which represents the ancestral ecosystem in this region may represent a “storehouse” of AMF species. We hypothesize that low sporulation in the pristine forest may be the result of latent fungal associates or fungi with predominant vegetative growth rather than partitioning biomass to spores. Once forests are cleared and other LUs are developed, then soil exposure, disturbance, and shifts in plant composition create distinct environmental and biological traits that trigger or stimulate sporulation. Low sporulation also might reflect low propagule density and infectivity in forest soils (unpublished results). Indeed, 30 species were detected in pristine forest, of which only three were exclusive to this ecosystem.
Few studies in mycorrhizal ecology have used species accumulation curves based on non-parametric estimators to determine the expected number of species in ecosystems. Picone (2000) used species accumulation curves in Costa Rica to compare AMF species richness between pasture and tropical forest sites. Whitcomb and Stutz (2007) used the first-order jackknife estimator to compare diversity in two plots of an urban experimental landscaped site. We chose the jackknife 1 algorithm because it considers (1) the number of species occurring only in a single sample, and (2) rare species are most useful to estimate overall species richness (Magurran 2004). In our study, many AMF species were rare or common within a LU (Table 2) and some species were detected only in a single sample.
This remote region of the Amazon forest domain is home to diverse AMF species, including some that have yet to be described. Shifts from the pristine forest ecosystem to other LUs with strong anthropogenic input appear to influence frequency and abundance of some species but not overall community structure and species richness. This study suggests that distinct LUs and their management practices commonly utilized by traditional human communities in the western Amazon are not causing a loss in diversity of AMF species.
References
Amelung T, Diehl M (1992) Deforestation of tropical rainforest – economic causes and impact on development. Tubingen, Germany, Kieler Studien 241
Bever JD, Morton JB, Antonovics J, Schultz PA (1996) Host-dependent sporulation and species diversity of arbuscular mycorrhizal fungi in a mown grassland. J Ecol 84:71–82
Bever JD, Schultz PA, Pringle A, Morton JB (2001) Arbuscular mycorrhizal fungi:more diverse than meets the eye, and the ecological tale of why. Bioscience 51:923–931
BRASIL (2002) Cenários para a Amazônia Legal. Bases para discussão. Ministério do Meio Ambiente—DSD, Brasília, CD-Rom
Cochrane TT, Sanchez P (1982) Land resources, soils, and their management in the Amazon region. In: Hech SB (ed) Amazonia: agriculture and land-use research. CIAT, Cali, pp 137–209
Coelho MR, Fidalgo ECC, Araújo FO, Santos HG, Santos ML, Pérez DV (2005) Solos das áreas-piloto do Projeto GEF BIOS (Conservation and Sustainable Management of Below-Ground Biodiversity: Phase I), Município de Benjamin Constant, Estado do Amazonas. Embrapa Solos, Rio de Janeiro, p 158, Boletim de Pesquisa
Cuenca G, de Andrade Z, Escalante G (1998) Diversity of glomalean spores from natural disturbed and revegetated communities growing on nutrient-poor tropical soils. Soil Biol Biochem 30:711–719
Daly DG, Prance GT (1989) Brazilian Amazon. In: Campbell DG, Hammond HD (eds) Floristic inventory of tropical countries. New York Botanical Garden, Bronx, pp 401–426
Fidalgo ECC, Coelho MR, Araújo FO, Moreira FMS, Santos HG, Santos MLM, Huising J (2005) Land use and land cover survey in benchmark area of Projeto BiosBrasil (Conservation and sustainable management of below-ground biodiversity: Phase I), Benjamin Constant (AM). Boletim de Pesquisa e Desenvolvimento 71. EMBRAPA Solos, Rio de Janeiro, p 54, in Portuguese
Fitter AH, Gilligan CA, Hollingworth K, Kleczkowski A, Twyman RM, Pitchford JW (2005) Biodiversity and ecosystem function in soil. Funct Ecol 19:369–377
Gerdemann JW, Nicolson TH (1963) Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans Br Mycol Soc 46:235–244
Guadarrama P, Álvarez-Sánchez FJ (1999) Abundance of arbuscular mycorrhizal fungi spores in different environments in a tropical rain Forest, Veracruz, Mexico. Mycorrhiza 8:267–270
Guadarrama-Chávez P, Camargo-Ricalde SL, Henrández-Cuevas L, Castillo-Argüero S (2007) Los hongos micorrizógenos arbusculares de la region de Nizanda, Oaxaca, México. Bol Soc Bot Méx 81:133–139
Hammer O, Harper DAT, Ryan PD (2001) PAST: Paleontological Statistics Software Package for Education and Data Analysis. Paleontologica Electronica 4(1) art4: 9 pp, 178 kb
Holt RD (1983) Ecology at the mesoscale: the influence of regional processes on local communities. In: Ricklefs RE, Schluter D (eds) Species diversity in ecological communities – historical and geographical perspectives. The University of Chicago Press, Chigaco, p 416
Huising EJ, Coe R, van Noordwijk M, Huang SP, Cares J, Louzada J, Zanetti R, Moreira FMS, Susilo F, Konate S (2008) Sampling strategy and design to evaluate below-ground biodiversity. In: Moreira FMS, Bignell D, Huising J (eds) Handbook of tropical soil biology: sampling and characterization of below-ground biodiversity. Earthscan, London, pp 17–42
Janos DP (1980) Vesicular-arbuscular mycorrhizae affect lowland tropical rain forest plant growth. Ecology 61:151–162
Johnson NC, Wedin DA (1997) Soil carbon, nutrients, and mycorrhizae during conversion of dry tropical forest to grassland. Ecol Appl 7:171–182
Lima AS, Nóbrega RSA, Barberi A, Silva K, Ferreira DF, Moreira FMS (2009) Nitrogen-fixing bacteria communities occurring in soils under different uses in the Western Amazon Region as indicated by nodulation of siratro (Macroptilium atropurpureum). Plant Soil 319:127–145.
Lovelock CE, Anderson K, Morton JB (2003) Arbuscular mycorrhizal communities in tropical forests are affected by host tree species and environment. Oecologia 135:268–279
Magurran AE (2004) Measuring biological diversity. Blackwell, Oxford
Mangan SA, Eom A-H, Adler GH, Yavitt JB, Herre EA (2004) Diversity of arbuscular mycorrhizal fungi across a fragmented forest in Panama: insular spore communities differ from mainland communities. Oecologia 141:687–700
Mendonça-Santos ML, dos Santos HG, Coelho MR, Bernardi ACC, Machado PLOA, Manzatto CV, Fidalgo ECC (2006) Soil and land use in the Brazilian Amazon. In: Moreira FMS, Siqueira JO, Brussaard L (eds) Soil biodiversity in Amazonian and other Brazilian ecosystems. CAB International, Oxfordshire, pp 13–42
Pasqualini D, Uhlmann A, Stürmer SL (2007) Arbuscular mycorrhizal fungal communities influence growth and phosphorus concentration of woody plant species from the Atlantic rain forest in South Brazil. For Ecol Manage 145:148–155
Peña-Venega CP, Cardona GI, Rguelles JH, Arcos AL (2007) Micorrizas arbusculares del Sur de la Amazonia Colombiana y su relación com algunos factores fiscioquímicos y biológicos del suelo. Acta Amazonica 37:327–336
Picone C (2000) Diversity and abundance of arbuscular mycorrhizal fungus spores in tropical forest and pasture. Biotropica 32:734–750
Rillig MC (2004) Arbuscular mycorrhizae and terrestrial ecosystem processes. Ecol Lett 7:740–754
Schenck NC, Perez Y (1990) Manual for identification of VA mycorrhizal fungi. Synergistic, Gainsville
Siqueira JO, Saggin-Junior OJ (2001) Dependency on arbuscular mycorrhizal fungi and responsiveness of some Brazilian native woody species. Mycorrhiza 11:245–255
Schwarzott D, Walker C, Schüssler A (2001) Glomus, the largest genus of the arbuscular mycorrhizal fungi (Glomales), is nonmonophyletic. Mol Phylogenet Evol 21:190–197
Siqueira JO, Colozzi-Filho A, Oliveira E (1989) Ocorrência de micorrizas vesicular-arbusculares em agro e ecossistemas do estado de Minas Gerais. Pesqui Agropecu Bras 24:1499–1506
Siqueira JO, Carneiro MAC, Curi N, Rosado SCS, Davide AC (1998) Mycorrhizal colonization and mycotrophic growth of native woody species as related to successional groups in Southeastern Brazil. For Ecol Manage 107:241–252
Stürmer SL, Siqueira JO (2006) Diversity of arbuscular mycorrhizal fungi in Brazilian ecosystems. In: Moreira FMS, Siqueira JO, Brussaard L (eds) Soil biodiversity in Amazonian and other Brazilian ecosystems. CABI, Oxfordshire, pp 206–236
Stürmer SL, Filho OK, de Queiroz MH, Mendonça MM (2006) Occurrence of arbuscular mycorrhizal fungi in soils of early stages of a secondary succession of Atlantic Forest in South Brazil. Acta Bot Bras 20:513–521
Violi HA, Barrientos-Priego AF, Wright SF, Escamilla-Prado E, Morton JB, Menge JA, Lovatt CJ (2008) Disturbance changes arbuscular mycorrhizal fungal phenology and soil glomalin concentrations but not fungal spore composition in montane rainforests in Veracruz and Chiapas, Mexico. For Ecol Manage 254:276–290
Zangaro W, Nishidate FR, Vandresen J, Andrade G, Nogueira MA (2007) Root mycorrhizal colonization and plant responsiveness are related to root plasticity, soil fertility and successional status of native woody species in southern Brazil. J Trop Ecol 23:53–62
Zhang Y, Gui LD, Liu RJ (2004) Survey of arbuscular mycorrhizal fungi in deforested and natural forest land in the subtropical region of Dujiangyan, southwest China. Plant Soil 261:257–263
Zhao ZW, Wang GH, Yang L (2003) Biodiversity of arbuscular mycorrhial fungi in a tropical rainforest of Xishuangbanna, southwest China. Fungal Divers 13:233–242
Wardle DA, Bardgett RD, Klironomos JN, Setälä H, van der Putten WH, Wall DH (2004) Ecological linkages between aboveground and belowground biota. Science 304:1629–1633
Whitcomb S, Stutz JC (2007) Assessing diversity of arbuscular mycorrhizal fungi in local community: role of sampling effort and spatial heterogeneity. Mycorrhiza 17:429–437
Acknowledgements
JOS thanks CNPq, Brazil for scholarship and SLS thanks the Pró-Reitoria de Pesquisa, Pós-Graduação e Extensão (FURB) for Research Grant (Edital Interno). This publication presents part of the findings of the international project “Conservation and Sustainable Management of Below-Ground Biodiversity” (CSM-BGBD), implemented in seven countries—Brazil, Côte d'Ivoire, India, Indonesia, Kenya, Mexico, and Uganda. This project is coordinated by the Tropical Soil Biology and Fertility Institute of CIAT (TSBF-CIAT with co-financing from the Global Environmental Facility—GEF), and implementation support from the United Nations Environment Programme (UNEP), and coordinated in Brazil by Dr. F. M. S. Moreira (UFLA, Lavras, MG). Views expressed in this publication are those of their authors and do not necessarily reflect those of the authors' organization, the UNEP and the GEF. Carlos R. Grippa is also acknowledged for contribution to soil sampling. We are in debt to two anonymous reviewers for their valuable contribution to this paper.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Stürmer, S.L., Siqueira, J.O. Species richness and spore abundance of arbuscular mycorrhizal fungi across distinct land uses in Western Brazilian Amazon. Mycorrhiza 21, 255–267 (2011). https://doi.org/10.1007/s00572-010-0330-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00572-010-0330-6