
Abstract
At embryonic day 8.5, the LIM-homeodomain factor Lmx1a is expressed throughout the otic placode but becomes developmentally restricted to non-sensory epithelia of the ear (endolymphatic duct, ductus reuniens, cochlea lateral wall). We confirm here that the ears of newborn dreher (Lmx1a dr) mutants are dysmorphic. Hair cell markers such as Atoh1 and Myo7 reveal, for the first time, that newborn Lmx1a mutants have only three sensory epithelia: two enlarged canal cristae and one fused epithelium comprising an amalgamation of the cochlea, saccule, and utricle (a “cochlear-gravistatic” endorgan). The enlarged anterior canal crista develops by fusion of horizontal and anterior crista, whereas the posterior crista fuses with an enlarged papilla neglecta that may extend into the cochlear lateral wall. In the fused endorgan, the cochlear region is distinguished from the vestibular region by markers such as Gata3, the presence of a tectorial membrane, and cochlea-specific innervation. The cochlea-like apex displays minor disorganization of the hair and supporting cells. This contrasts with the basal half of the cochlear region, which shows a vestibular epithelium-like organization of hair cells and supporting cells. The dismorphic features of the cochlea are also reflected in altered gene expression patterns. Fgf8 expression expands from inner hair cells in the apex to most hair cells in the base. Two supporting cell marker proteins, Sox2 and Prox1, also differ in their cellular distribution between the base and the apex. Sox2 expression expands in mutant canal cristae prior to their enlargement and fusion and displays a more diffuse and widespread expression in the base of the cochlear region, whereas Prox1 is not detected in the base. These changes in Sox2 and Prox1 expression suggest that Lmx1a expression restricts and sharpens Sox2 expression, thereby defining non-sensory and sensory epithelium. The adult Lmx1a mutant organ of Corti shows a loss of cochlear hair cells, suggesting that the long-term maintenance of hair cells is also disrupted in these mutants.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The vertebrate ear has 3–9 sensory epithelia consisting of hair cells and supporting cells (Lewis et al. 1985). Mammals have three canal cristae, two gravistatic organs (utricle, saccule), an organ of Corti in the cochlea, and a papilla neglecta that varies in size (Fritzsch and Wake 1988). The initially continuous sensory epithelia become separated as a result of unknown developmental mechanisms (Fritzsch et al. 2002) by non-sensory epithelia that orient the sensory epithelia in space, channel fluid dynamics, and maintain the endolymphatic environment (Lewis et al. 1985). Sensory and non-sensory epithelia generate diffusible factors that govern the morphogenesis of nearby non-sensory epithelia (Chang et al. 2004a, 2008; Daudet et al. 2002; Sienknecht and Fekete 2008). Additional secreted factors originate from the hindbrain, ectoderm, and mesenchyme (Chang et al. 2004b; Fritzsch et al. 2006b; Ohyama et al. 2007; Pirvola et al. 2004). Thus, both global and local interactions of various diffusible factors regulate local transcription factors that govern the morphogenetic process of the non-sensory epithelium of the ear, a process that ultimately channels physical stimuli to specific sensory epithelia. Likewise, the differentiation of sensory epithelia into hair cells and supporting cells reflects temporal expression cascades of transcription factors (Fritzsch et al. 2006a; Kelley 2006; Kiernan et al. 2005). However, no single factor has been described that is exclusively associated with the non-sensory epithelia during development and throughout the ear (Chang et al. 2008; Kiernan et al. 1997; Raft et al. 2004), although the vast majority of Wnt transcripts are expressed in non-sensory domains (Sienknecht and Fekete 2008). Hence, a possible feedback loop between developing sensory and non-sensory areas of the ear could exist to fine-tune the morphogenesis of the ear to the histogenesis of the sensory epithelia.
Lmx1a is one of four members of the Islet-Lim homeodomain transcription factor family (Hunter and Rhodes 2005), which has three conserved members in triploblastic animals (Isl1, Lmx1a, Lmx1b; Drosophila orthologs: tailup, CG32105, CG4328, respectively). The Islet family belongs to a large family of Lim-homeodomain transcription factors that can bind to DNA in the form of monomers that form complexes with other transcription factors or in the form of heteromultimeres (Bhati et al. 2008b; Hunter and Rhodes 2005; Matthews and Visvader 2003). GATA, bHLH, and LMX factors interact with Lim-homeodomain factors during development. For example, such complexes are required during the development of reticular formation in the hindbrain (Alenina et al. 2006) and motoneuron formation in the spinal cord (Lee et al. 2008; Matthews and Visvader 2003). Likewise, in insect mechanosensory development, Isl and Gata/pannier antagonize each other to regulate the expression of bHLH genes necessary to develop sensory and non-sensory cells (Asmar et al. 2008; Biryukova and Heitzler 2005), presumably through competition for binding to another Lim-homeodomain factor. Consistent with the emerging concept of the molecular conservation of essential neurosensory developmental modules across phyla (Adam et al. 1998; Caldwell and Eberl 2002; Fritzsch et al. 2000, 2007; Pierce et al. 2008), Gata3 is necessary for neurosensory development of the vertebrate ear (Karis et al. 2001; Lillevali et al. 2006). Interestingly haploinsufficiency of Gata3 causes hearing loss (Van Esch and Devriendt 2001). In the ear, the expression of Isl1 (Radde-Gallwitz et al. 2004), Lmx1a (Failli et al. 2002), Lhx3 (Hertzano et al. 2007) and Lim only factors (LMOs) have been described (Deng et al. 2006), but no functional analysis using LoF or Gof as yet exists.
The dreherJ (Lmx1a dr) point mutation is one of 13 known spontaneous mutations in the Lmx1a gene causing neurological, skeletal, and otic abnormalities (Chizhikov et al. 2006; Millonig et al. 2000). The morphology of the dreherJ mutant ear was initially described by Deol (1964, 1983) and was thought to be a consequence of malformations in the hindbrain (Manzanares et al. 2000). More recent in situ expression studies show, however, a more robust and earlier expression of Lmx1a in the developing mouse (Failli et al. 2002) and chicken ear compared with expression in the hindbrain (Giraldez 1998). These in situ hybridization (ISH) data raise the possibility that local otic Lmx1a expression is required for ear development, and that its absence in the ear is causally linked to the ear defects. Since Lmx1a interacts with other Lim and LMO factors, an intriguing and likely possibility is that Lmx1a and other Isl family members co-operate to regulate the sensory and non-sensory development of the ear. Their role in ear morphogenesis could thus parallel that of tailup/pannier in fly mechanosensory development (Biryukova and Heitzler 2005) and could display a conserved interaction of bHLH, Gata and Lim transcription factors in the regulation of mechanosensory development across phyla (Fritzsch et al. 2007).
In agreement with this hypothesis, Lmx1a is predominantly expressed in the non-sensory otic epithelium, and Lmx1a dr mutant mice show fusion and enlargement of sensory epithelia, dysmorphogenesis of the ear, and disrupted histogenesis of sensory epithelia eventually leading to the degeneration of hair cells. These data suggest that Lim domain factors indeed play a possibly conserved role in regulating the distinction between sensory and non-sensory epithelia in mechanosensory development across phyla. Further work is needed to unravel the details of the molecular interactions that are regulated by Lmx1a in the developing mouse ear.
Materials and methods
Mice
Atoh1tm2Hzo mice were obtained from Dr. Huda Zoghbi (Bermingham et al. 2001) and Lmx1adr/J mice from Jackson Labs; they were and maintained in an AALAC approved facility under an IACUC approved protocol. Breeding and genotyping of the mice was as previously described (Bermingham et al. 2001; Millonig et al. 2000). Experimental animals were of mixed genetic stock. Timed breeding took place overnight, with midnight being considered time 0.0; noon of the first day was considered as embryonic day 0.5 (E0.5). Postnatal day 0 (P0) was the equivalent of embryonic day 19 (E19) regardless of the actual birth date.
Detection of β-galactosidase activity
To detect β-gal activity, ears were dissected, briefly (30 min) fixed in 4% paraformaldehyde/phosphate-buffered saline, rinsed in phosphate buffer, and stained with “X-gal.” as previously described (Fritzsch et al. 2005a). When required, we enhanced the “X-gal.” reaction by using two-photon photoactivation on whole-mounts and sections (Matei et al. 2006). Stained ears were mounted flat or, alternatively, embedded in a soft Epoxy resin, sectioned (3 μm) with a histology-grade diamond knife (Dumont), imaged by using a compound light microscope (Nikon Eclipse 800), and captured by means of a Coolsnap camera and Metamorph software.
Immunohistochemistry and ISH
Primary antibodies were rat anti-mouse β-tubulin (Sigma; #T6793, 1:800), Hoechst nuclear stain (Sigma) and MyoVII (gift of T. Hasson, San Diego) and chicken anti-BDNF (anti-brain-derived neurotrophic factor; R&D Systems; 3AF248, 1:100). Whole-mount ISH was carried out according to standard procedures (Pauley et al. 2003) with digoxigenin-labeled riboprobes specific for Sox2, Fgf8, Gata3, and Fgf10. Anti-dig-AP antibody and BM Purple (Roche) colorimetric signal detection was used. Some whole-mount reacted ears were subsequently embedded in epoxy resin, cut at a thickness of 5–10 μm, counterstained with toluidine blue, and viewed with a Nikon E800 microscope by using differential interference contrast.
Secondary Alexa 488-, 543-, and 634-conjugated antibodies (Molecular Probes) were used predominantly on whole-mounted microdissected sensory epithelia (Matei et al. 2005). Sections and whole-mounts were imaged by using a confocal system (Biorad 2000 mounted on a Nikon E800 or Zeiss LSM 510). Images were assembled into plates by means of CorelDraw software.
Lipophilic dye tracing
PTI lipophilic tracers (NV red, NV Maroon) were used for afferent and efferent neuronal fibers (Fritzsch et al. 2005b). Briefly, dyes were inserted into central targets, and the fibers were filled with the diffusible dye. The epithelia were microdissected and viewed with a confocal system (Zeiss LSM 510).
Imaging for scanning electron microscopy
Ears were microdissected, osmicated, dehydrated, and critical-point-dried as previously described by Ma et al. (2000). They were mounted on stubs and imaged with a Hitachi scanning electron microscope.
Results
Lmx1a expression is concentrated in certain non-sensory epithelia patches
Lmx1a expression was shown to be widespread throughout the ear between E8.5 and E10.5 (Failli et al. 2002), but these expression analyses were limited to only those early embryonic ages. We therefore extended these investigations of Lmx1a expression by using ISH. At E10.5, virtually the entire otocyst was positive for Lmx1a (Fig. 1a) except for a small anteroventral quadrant, the area of prosensory formation (Farinas et al. 2001; Fekete and Wu 2002; Ma et al. 1998). Over the next 2 days, Lmx1a expression became focused in the developing endolymphatic duct (Fig. 1c,f,g) and the lateral margin of the cochlear duct (Fig. 1g). Strong expression also persisted in the saccular roof (Fig. 1h–j), in the ductus reuniens (Fig. 1h,i), and near pigment cells in the utricular roof and the canal cristae (Fig. 1h,j). In the cochlea, Lmx1a was immediately lateral to the developing organ of Corti and medial to the pigment cells of the stria vascularis (Fig. 1k). These data suggest that Lmx1a outlines certain non-sensory epithelia of the ear and may be involved in specifying sharp boundaries between sensory and non-sensory epithelia.

Lmx1a expression, as shown by in situ hybridization (ISH) undergoes dynamic changes in wildtype (wt) and mutants (ant anterior, dors dorsal, AC anterior crista, PC posterior crista, S saccule, SL spiral limbus, U utricle). a In the wildtype at embryonic day 10.5 (E10.5), expression is in all but the neurosensory precursor epithelium (NSP). b, c Compare the wildtype endolymphatic duct (ED) with its vestigial mutant counterpart. dLmx1b is not expressed in the E10.5 ear (dotted line) but is strongly expressed in the hindbrain (labeling and be seen in and out of focus; top left). e f In the E11.5 mutant, Lmx1a expression is incompletely segregated to the lateral/posterior cochlea (CO), and no trace of an endolymphatic duct is apparent. g At E12.5, expression is becoming confined to the endolymphatic duct (ED), lateral cochlea, and non-sensory saccule. h–j By E14.5, expression centers on the ED and radiates to constricted spacer epithelia between sensory epithelial territories. Most conspicuous of these is the ductus reuniens (DR). k At E18.5, cochlear expression is localized to the outer spiral sulcus (OSS). Melanocytes are conspicuous in the adjacent stria vascularis (SV). l–n In wildtype mice, the organ of Corti (OC) is entirely free of Lmx1a expression (n). However, mutant Lmx1a mRNA is broadly expressed throughout the basal turn (l, m). All whole ears are oriented as in a. Bars 100 μm
Since Lmxa1 dr is a nonsense mutation (Millonig et al. 2000), the presence of the mutated Lmx1a mRNA permits detection of alterations in Lmx1a expression patterns in these functional null mutants. Alteration of normal Lmx1a expression became evident at E11 in the mutant ears. Unlike the obvious concentrated expression in the endolymphatic duct in the wildtype, Lmx1a expression remained widespread in the mutant (Fig. 1b,c). The endolymphatic duct in Lmx1a mutants never developed beyond a rudimentary structure (Fig. 1e,f). The pattern of Lmx1a distribution in the mutants suggests a loss of segregation to non-sensory epithelia as evidenced by the overlapping distribution of expression within the basal sensory region, instead of clear segregation to the outer spiral sulcus (Fig. 1l-n).
Although Lmx1a and Lmx1b diverged before the split of protostomia and deuterostomia, they both still share large areas of expression in the mammalian brain (Chizhikov et al. 2006). In order to determine whether a similar overlapping expression pattern exists in the inner ear, we investigated the expression of Lmx1b. The well-characterized expression of Lmx1b in the hindbrain and isthmus region was replicated, but no significant expression of Lmx1b was observed in the otocyst (Fig. 1d) or in later stages of the ear formation in wildtype or Lmx1a mutant mice. Based on these observations, Lmx1b could be eliminated as having a direct role in inner ear development. However, the strong and early expression in the adjacent hindbrain might indicate that Lmx1b expression indirectly affects ear development.
We next investigated the distribution of Lmx1a expression in near radial sections of epoxy-resin-embedded E18.5 ears analyzed by ISH for Lmx1a (Fig. 2). These data showed a clear medial to lateral organization of the cochlea in the mutant. However, the cellular organization of the organ of Corti did not show the single clear row of inner hair cells and the 3–4 rows of outer hair cells found in the wildtype (Fig. 2a-d). Consistent with our data on whole-mounted ears, we found a considerable expansion of the expression area of the mutated Lmx1a mRNA beyond the lateral wall area of the wildtype. Expression sometimes expanded to the stria vascularis and even to Reissner’s membrane (Fig. 2a’-d’). In the absence of functional Lmx1a protein, mechanisms that help to focus Lmx1a expression onto the narrow part of the lateral wall between Claudius cells and the stria vascularis did not appear to work properly.

Lmx1a expression is less restricted in mutants. The distribution of Lmx1a ISH reaction product is shown in wildtype (b, b’, d, d’) and Lmx1a mutant mice at embryonic day 18.5. Ears were reacted for whole-mount ISH (see Fig. 1), embedded in soft epoxy resin, and sectioned at 5–10 μm. These radial sections show a well-organized organ of Corti (OC) with 1 inner and 3–4 rows of outer hair cells in the wildtype (b, b’, d, d’). In contrast, hair cells are disorganized in the Lmx1a mutant mice (a, a’, c, c’). Nevertheless, the main medial-to-lateral areas of the cochlea such as greater epithelial ridge (GER) with a tectorial membrane (TM), OC, and lateral wall are distinct. In wildtype mice, the Lmx1a in situ signal is in the lateral wall (stars) adjacent to the stria vascularis (SV in b’, d’). In contrast, in the Lmx1a mutant mice, the strong Lmx1a in situ signal expands to include the SV (stars) or even Reissner’s membrane (RM in a’,c’). Bar 100 μm
Mutation of Lmx1a gene produces unique inner ear phenotype
The altered Lmx1a expression pattern in dr mutants suggests a basis for the observed disruption of sensory epithelium segregation cues. Since the previous studies described only inner ear dysmorphogensis in the Lmx1a mutant lines (Deol 1964, 1983; http://www.informatics.jax. org/Lmx1a alleles), we extended these preliminary observations to define the extent of this interference in sensory and non-sensory epithelium formation. The Atoh1 tm2Hzo allele carrying the targeted LacZ reporter (Atoh1 LacZ; Fritzsch et al. 2005a) was used to identify both differentiated and undifferentiated hair cell precursors and, in turn, assisted in defining the regions of the developing sensory epithelium.
Postnatal 1- and 2-week-old (P7 and P14) wildtype mice had six discrete AtohLacZ-positive sensory patches located in a complex three-dimensional labyrinth of ducts and recesses (Fig. 3a). The papilla neglecta was barely detectable and consisted of 5–8 hair cells (data not shown). In contrast, P7 and P14 Lmx1a dr/dr mutants had only a single undivided sac that was wide in the region of the canal cristae and continuously tapered toward the apical tip of the cochlea. Whereas each sensory epithelium of the wildtype ears resided within its own recess (three ampullae for the canal cristae, the utricular and saccular recesses being separated by the constricted utriculo-saccular foramen, and the cochlear duct being separated from the saccule by the ductus reuniens), none of these non-sensory constrictions was found in Lmx1a mutants, nor was the constriction that normally separates the posterior ampulla from the cochlea. The data suggest that Lmx1a is, directly or indirectly, involved in the morphogenesis of the specific constrictions and ducts that separate the individual sensory epithelia. These mutant ears also lacked an endolymphatic duct/sac (Figs. 1e,l, 3b).

Postnatal Lmx1a mutant ears reveal disorganized sensory epithelia. a In the wildtype, six discrete sensory epithelia are separated from one another by constricted non-sensory epithelial spacers. Two of three semicircular canals (Ant Canal anterior canal, Post Canal posterior canal) and an endolymphatic duct (Endo duct) can be identified as shadows (Common cruc common cruciate eminence, DR ductus reuniens, OC organ of Corti, PC posterior crista, S saccule, U utricle, USF utriculo-saccular foramen). b In the Lmx1a mutant, the anterior cristae (AC) and horizontal cristae (HC) are separated by a common cruciate eminence, whereas the posterior crista is grossly enlarged and extended by the presence of both embedded and detached papilla-neglecta-like sensory epithelia (PN). The utricle (U), saccule (S), and cochlear sensory epithelia seem to be continuous with one another. The basal turn of the organ of Corti (OC) appears as a uniform band of hair cells that is discretely separated (arrow) from an apex in which inner and outer hair cells can be identified. c Higher magnification view of the transition (arrow) in b. The densely packed hair cells of the basal cochlea lie above the arrow, with the apex (IHC inner hair cells, OHC outer hair cells) below it (black lines planes of section in e, f). d Same tissue as in c but stained for beta-tubulin to reveal nerve fibers (RF radial fibers), pillar cells (P), and Deiters’ cells (DC). Note the absence of tubulin-containing pillar and Deiters’ cells in the base and their conspicuous appearance at the transition to the apex. e. A medio-lateral near radial section across the base of the mutant cochlear duct (see dotted line in c). Note the presence of a tectorial membrane (TM). Up to 11 rows of hair cells are marked by the blue Atoh1LacZ reaction product; unlabelled supporting cells are present below the hair cells (HP habenula perforate, HC horizontal crista). f A recognizable organ of Corti with inner and outer hair cells is present in the apex (compare with wildtype, inset). g, h Beginning at ∼P14, hair cells disappear, starting in the base (arrow). In contrast, nerve fibers continue to mature as indicated by the osmium-tetroxide-stained myelin (black fibers). i, j By P114, the organ of Corti is grossly disorganized, lacks identifiable hair cells, and shows massive aberrations in almost all associated epithelia such as the spiral limbus (SL) and the stria vascularis (SV). These data suggest that absence of Lmx1a is ultimately incompatible with hair cell maintenance, although that its absence does not interfere with their initial formation. Bars 100 μm
Fusion of inner ear sensory endorgans
At P7/P14, only three distinct sensory epithelia instead of the usual six were found in Lmx1a mutants (Fig. 3a,b). Near the anterodorsal pole, a single crista consisting of two unequal sized hemicristae was separated by a non-sensory cruciate eminence, suggesting that the horizontal crista had fused with one hemicrista of the anterior crista. Likewise, the posterior crista consisted of two asymmetric hemicrista separated by a cruciated eminence. The enlarged hemicrista sometimes had a distinct extension or patches extending into the cochlear duct (Fig. 3b), possibly representing a fusion of the posterior canal hemicrista with the papilla neglecta. The papilla neglecta generally consists only of a few hair cells in mammals but can be large in elasmobranchs and becomes the comparatively large amphibian papilla in frogs (Fritzsch and Wake 1988, Lewis et al. 1985).
In addition to these two enlarged cristae, the Lmx1a dr ear contained one large continuous band of hair cells, with an extensive patch that lay adjacent to the anterior crista, and that we tentatively identified as a utricle-like region based on its topology (Fig. 3b, see also below). This utricular area blended into a smaller elongated patch that we identified as a putative saccule. The saccular region continued into a progressively tapering band of hair cells that stretched to the apex of what appeared to be a shortened cochlear duct with the organ of Corti. Whereas the hair cells of this patch were continuous, the pattern of innervation indicated four distinct regions in this epithelium (Figs. 3b,g, 4c). The utricle region was innervated by fibers that were accompanied by others that continued on to the anterior crista, as in wildtype animals. Likewise, the saccular portion received fibers from both the superior and inferior vestibular ganglion, as in the wildtype (Fig. 4c). In contrast, the cochlea organ of Corti showed two distinct regions with respect to its innervation: the base received only a patchy and reduced density of nerve fibers, whereas the apex was densely innervated with radial fibers, like the wildtype (Figs. 3d, 4a–c). These fibers however arose from a typical spiral ganglion.

Late innervation and sensory epithelia are disorganized in mutants. Afferent radial fibers to the base of mutant cochlea stained with the lipophylic dye, NV Maroon, at the E18.5 (a, b), and with anti-acetylated tubulin at P7/P14 (c–i). a, b Note that the fibers enter the organ of Corti(OC) but do not extend to the outer hair cells (OHC) as they would in a comparable wildtype ear. There is a notable difference between the packing density of radial fibers (RF) in the base and the apex consistent with the reduced presence of spiral ganglia (Spg) in the base. c There is a complete absence of pillar/Deiters’ cells basal to the cochlear transition (arrow) and a clear distinction of innervation between the densely innervated saccule (S) and the poorly innervated basal turn of the cochlea (AC anterior crista, HC horizontal crista, PC posterior crista, U utricle). d Disorganized Deiters’ cell processes (D) in the apex of the ear at P7 stained for β-tubulin and showing longitudinal extension along the cochlea (P pillar cells). e Myosin VIIa (Myo7a) staining revealing close proximity of the OC and hair cells of the papilla neglecta (PN) in the basal cochlea of a P7 mutant. f Basal/apical cochlear transition of the ear in c, with supporting cells stained for β-tubulin and hair cells stained for Myo7a. Note that the packing density of hair cells is inversely related to supporting cell labeling. This pattern is maintained for as long as hair cells are labeled by Myo7a antibodies (i). g Apical cochlea of the ear in c, d. Inner hair cells (IHC) and outer hair cells (OHC) can be recognized, but the organization is inferior to that of the wildtype (see Atoh1LacZ-stained hair cells in inset; top mutant apex, bottom wildtype). h Grossly enlarged and elongated posterior crista (PC) of a P14 mutant showing fibers targeted to the Myo7a-positive hair cells. Bars are 100 μm
In addition to the differences in density of innervation, older Lmx1a mutant mice showed tubulin-positive pillar and Deiters’ cells only in the apex (Figs. 3d, 4d,f,g,i). The absence of these cells in the base correlated with a different distribution of hair cells. In the apex, both inner and outer hair cells could be identified and were separated from one another by unusually positioned but otherwise typical pillar cells (Figs. 3c-f, 4e-i). This was therefore identifiable as having an organ of Corti-like organization of cells. In contrast, in the base, hair cells formed multiple rows of uniform cells that defied any histological characterization as inner or outer hair cells (Figs. 3c,e, 4f,i). Despite their unusual pattern of distribution, all were hair cells as revealed by both Atoh1 LacZ histochemistry (Fig. 3b,c,g) and immunofluorescent detection of the hair cell-specific marker, myosin VIIa (Myo7a; Fig. 4d,f,h,i). The common utriculo-sacculo-cochlear sensory epithelium (Figs. 3b, 5a) of dr mutant mice is thus reminiscent of the common macula of jawless vertebrates (Lewis et al. 1985). This common macula is then the precursor of the several sensory epithelia that segregate from one another during the course of development and evolution (Fritzsch 2003; Fritzsch et al. 2002). Clearly, Lmx1a is a major molecular player in this process during development and may have played the same role during ear evolution.

Atoh1LacZ staining and Prox1 immunodetection showing disorganization of hair and supporting cells. In these confocal micrographs of E16.5 Lmx1a null mutants, Atoh1LacZ is UV-activated and false-colored blue. Prox1 is immunostained red. a The fields shown in b, c are boxed (OC organ of Corti, PC posterior crista, PN papilla neglecta, S saccule, U utricle). Inset: Wildtype (Wt) Prox1 staining pattern of supporting cells at E18. Note that the supporting (pillar, Deiters’) cells are precisely organized. b Note the basal-apical transition (arrow). The basal OC lies upper right of the arrow. Note that an unorganized mass of Prox1-positive cells is located just basal to the transition. Blue hair cells are located medial to the Prox1-positive cells. c Mutant E16.5 cochlear apex demonstrating a disorganized, but otherwise continuous band of supporting cells in the OC. Bars 100 μm
Absence of hair cells in adult Lmx1a mutant cochlea
Using Atoh1LacZ histochemistry, we found gaps in the distribution of cochlear hair cells as early as P14 (Fig. 1g,h, arrow). Notably, these gaps tended to appear at the boundaries between the utricle, saccule, and basal region of the cochlea. Likewise, the apex showed a patchy distribution of hair cells in the older epithelia (Fig. 3g). Investigation of the cochlear histology at 2–3 months of age or older revealed a complete loss of all hair cells and severe dysmorphogenesis of the organ of Corti, including the adjacent areas such as the spiral limbus, Reissner’s membrane, and stria vascularis (Fig. 3i,j). A few hair cells remained in the vestibular organs (data not shown). These data suggest that Lmx1a not only plays a major role in early histogenesis and morphogenesis of the ear, but is also essential for the long-term maintenance of hair cells.
Amalgamated cochlear-gravistatic endorgan boundary
In a series of experiments, we next characterized the unique morphological and histological phenotype of the fused cochlear-gravistatic endorgan. Scanning electron microscopy was used to define the apical specializations in the area in which the saccular and cochlear regions merged (Fig. 6a,b). Despite our best efforts, we could not mechanically remove the tectorial membrane near this transition site. Importantly, the presence of a tectorial membrane defines a molecular transition between the saccule (no tectorial membrane) and the cochlea (tectorial membrane), despite the continuity of hair cells. Closer examination revealed distinct vestibular-type hair cells in the saccular and utricular regions (Fig. 6c–e). Near the tectorial membrane, we found that medial cells displayed vestibular-type long organ-pipe-like stereocilia. In contrast, more C-shaped, shorter stereocilia reminiscent of the inner hair cells of the organ of Corti prevailed more laterally, indicating an introgression zone of the vestibular and organ-of-Corti-type of hair cell differentiation. Ventral to this zone, radial histological sections (Fig. 3e) showed that 8–11 rows of hair cells were present in the basal part the cochlear region. Interestingly, the polarity of these inner-hair-cell-like cells was normal in the more medial cells, but the more lateral cells were rotated 90° toward the base, much as in mice mutant for the Foxg1 (Pauley et al. 2006) and Neurog1 (Ma et al. 2000) genes.

Continuity of hair cells in late embryos; Atoh1 LacZ staining and scanning electron microscopy. a Sensory epithelia of the E16.5 Lmx1a mutant ear revealed by Atoh1 LacZ staining of hair cells. Note that the juxtaposition of sensory epithelia found at P7 in Fig. 1b is apparent even at E16.5 (AC anterior crista, CO cochlea, HC horizontal crista, PC posterior crista, S saccule, U utricle). b A P0 Lmx1a mutant ear oriented similar to that in a and viewed by scanning electron microscopy. The positions of the micrographs in g (G), h (H) are indicated bottom (OC organ of Corti, TM tectorial membrane). c Higher power micrograph centered on the saccule (S) in b. The positions of micrographs in d–f (D-F) are indicated, as is the smooth flat surface of the TM. d Saccular macula. The sterocilia appear as long and immature short variations of the pipe-organ arrangement characteristic of vestibular hair cells. e Region of the basal cochlea (note the adjacent TM in c) close to the saccule. A mix of hair cells with long vestibular-like and short C-shaped cochlear inner-hair-cell-like sterocilia are present. f Further toward the apex, but still in the basal cochlea, vestibular-type hair cells are found adjacent to the tectorial membrane, with cochear-like hair cells lying further laterally. g Still in the base, but further toward the apex, multiple “rows” of hair cells all possess short cochlea-like sterocilia bundles. h Variously oriented cochlear hair cells are present (arrow indicates polarity) adjacent to g in the base. Medial is left and lateral right in d–h. Bars 100 μm
This non-cochlear organization of hair cells and supporting cells in the cochlea-like basal region might result from mis-expression of transcription factors uniquely associated with the organ of Corti. One transcription factor associated with supporting cells is Prox1 (Bermingham-McDonogh et al. 2006; Fritzsch et al. 2008; Puligilla et al. 2007). Prox1 distribution in Lmx1adr mice showed a marked deviation from the wildtype, even in the apex, which was more typical in its supporting cell and hair cell organization (Figs. 5a-c, 7a-d). Instead of a regular pattern of supporting and hair cells, Prox1-positive cells in the apex of Lmx1a mutants showed an irregular distribution with an inconsistent number of rows. To obtain more information regarding the co-localization of hair cells and Prox1-positive supporting cells, hair cells were labeled by immunodetection for BDNF. In Lmx1a mutants, the innermost row of hair cells showed strong BDNF staining, much like that observed in wildtype mice. However, unlike in the wildtype, BDNF was just above background levels in the most lateral outer hair cells of the mutants (Fig. 7b”). The mutant distribution of Prox1 near the transition from the near normal apical organ of Corti to the more vestibular-like basal region was the most interesting. Prox1 could not be detected in the base, except for a large aggregate of labeled cells near the cochlear-saccular transition zone (Fig. 7c,d). We sought to determine how early this organization became apparent and found that it occurred as early as E16.5 by using Prox1 immunodetection and Atoh1LacZ labeling of hair cells (Fig. 5). As in later stages, little to no Prox1 was expressed throughout the base, except at this transition zone (Figs. 5, 7). In wildtype mice, outer hair cells are regularly interspersed with Prox1-expressing Deiters’ cells (Fig. 7a’’’). However, in Lmx1a mutants at these stages, all hair cells were medial to the Prox1-expression area, in both the apical and basal regions (Fig. 7c’’’). Combined, these data suggest that Prox1- and Atoh1-expressing cells partially segregate in the absence of Lmx1a expression further implying that Lmx1a functions in the setting up of the topology for these two cell types and their specific distribution along the cochlea. A possible candidate could be diffusible factors emanating from the lateral spiral sulcus (Fig. 1k, 2b’) where Lmx1a expression is most prominent.

Hair cells, supporting cells, and nerve fibers are disorganized in mutant mice. Organ of Corti at E18.5 stained immunohistochemically for Prox1, β-tubulin, brain-derived neurotrophic factor (BDNF), and all three combined (a’’’–d’’’), in the wildtype (a–a’’’), mutant apex (b–b’’’), mutant base (c–c’’’), and high magnification of mutant base (d–d’’’). a–d Immunohistochemistry for Prox1. Note the orderly expression in supporting cells in the wildtype, some organization in the apex in the mutant, but clumping of PROX1-positive supporting cells in the mutant base (see also a’’’-d’’’). a’-d’ Immunohistochemistry for β-tubulin. Note that nerve fibers in the base stop short of the OC (see also a’’’-d’’’). a’’-d’’ Immunohistochemistry for BDNF stains inner hair cells (IHC) strongly and outer hair cells (OHC) weakly in the wildtype. A row of IHCs can be recognized in the mutant apex, but strongly stained cells are scattered in the base. a’’’-d’’’ The superimposed images show that hair cells and supporting cells overlap and orderly nerve fibers project between the supporting cells in the wildtype, whereas hair cells and supporting cells do not overlap in mutants with disorganized fibers projecting between the labeled supporting cells (apex) or reaching the hair cells (base). Bars 100 μm
Distinct basal cellular organization can be defined by gene expression
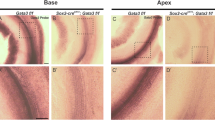
We next examined Gata3, Fgf8, and Fgf10 transcript distribution to elucidate further the extent of the disorganization of the basal region at the level of gene expression. The distribution of the transcript for Gata3, the gene for a zinc finger transcription factor, is uniquely associated with the cochlea, but it is not observed in the vestibular (specifically, saccular) sensory epithelia (Karis et al. 2001). It is known to interact with the Lim-homeodomain and bHLH factors needed for cell-fate specification (Matthews and Visvader 2003). In addition, the two fibroblast growth factors (Fgf) have distinct expression patterns, Fgf8 being associated with the inner hair cells and Fgf10 being expressed in the greater epithelial ridge just adjacent to the inner hair cells (Pauley et al. 2003; Pirvola et al. 2002). Expression of both Fgf10 and Gata3 showed that the base region gradually merged into the apex with respect to these markers but was distinct from the saccular region (Fig. 8a,b,e,g). There was also an expansion of Gata3 expression across the cochlea in Lmx1a mutants compared with wildtype littermates (Fig. 8e,g). In Lmx1a mutants, Fgf8 expression expanded across most of the several rows of hair cells in the basal organ of Corti, whereas only inner hair cells were labeled in the apical region, as in wildtype littermates (Fig. 8c,d,f). Most interestingly, Lmx1b, which shares high sequence homology with Lmx1a, is a known regulator of Fgf8 in the isthmic region of mice and zebrafish (Alexandre et al. 2006; Guo et al. 2007; O’Hara et al. 2005). Since our data show an expansion of Fgf8 expression in the base (Fig. 8c), but no Lmx1b expression has been detected in the ear, this suggests that wildtype Lmx1a somehow restricts Fgf8 expression in the basal cochlea to the inner hair cells. This might result from its early expression in this region during the otocyst stage or from diffusible factors released as a result of its later expression in the outer spiral sulcus (Daudet et al. 2002).

Fgf10, Fgf8, and Gata3 mRNA expression reveal disorganization of the organ of Corti in mutants. a In the E18.5 Lmx1a mutant, Fgf10 is strongly expressed in the entire organ of Corti (OC), but more weakly in the vestibular sensory epithelia (PC posterior crista, S saccule, Spg spiral ganglion, U utricle). b Mutant Fgf10 expression extends from the greater epithelial ridge (GER) laterally to the inner hair cells in a pattern similar to that of the wildtype. This pattern is the same in the base and the apex. c, dFgf8 expression in the base and apex of the mutant, respectively. Note that expression is confined to the inner hair cells (IHC) in the apex but is scattered among the excess hair cells of the base (HC horizontal crista). f In the wildtype (Wt), Fgf8 expression is neatly confined to the inner hair cells (IHC), with none in the outer hair cells (OHC). e In the E18.5 mutant, Gata3 expression is present in the basal cochlea, but not the saccule. g Similar Gata3 expression in the wildtype cochlea. Bars 100 μm
The Fgf8 data support the impression derived from the scanning electron-microscopic data of the Lmx1a mutant, viz., that all hair cells in the basal portion of the cochlear region develop an inner hair cell phenotype. Furthermore, alterations in Fgf8 and Gata3 expression and the near complete absence of Prox1-expressing cells in the base suggest that these too play roles in generating the altered phenotype of the basal-turn hair cells. Consistent with the Lmx1a expression pattern (Fig. 1n), Lmx1a is apparently necessary to define the lateral boundaries of the organ of Corti (which is more irregular in these mutants) and to enhance the interaction of supporting cells and outer hair cells. The absence of functional Lmx1a protein may underlie the disruption of outer hair cells and Deiters’ cells observed in the dr mutant mice (Fig. 3), possibly through indirect effects on the integrity of sensory/non–sensory boundaries. Proper cellular restriction of the early expression of the Lmx1a transcription factor appears to be necessary for the coordinated development of the organ of Corti.
Absence of segregation of utricle, saccule, and organ of Corti during development in the Lmx1a mutant
Given that our Lmx1a expression analysis suggested a possible role in ear formation as early as E11 (Fig. 1b,c), the initial upregulation of Atoh1 LacZ was compared in Lmx1a mutants and wildtype mice (Chen et al. 2002; Matei et al. 2005). As early as E14.5, the hair cells of the six epithelia of the wildtype ear were distinctly labeled (Fig. 9a). In the mutant, and in contrast to later stages, three cristae could be recognized. However, a single cochlear–gravistatic endorgan was in place, even at this stage, although distinct hair cell patches were observed within the common endorgan. Specifically, an area of constricted hair cell formation indicated the utricular/saccular regional transition (Fig. 9b,c), and the cochlea showed little to no upregulation of Atoh1LacZ in the base. In spite of this absence of staining, a basal cochlear prosensory precursor epithelium could be identified by using differential interference microscopy (Fig. 9c). These data suggested a delay in hair cell maturation in the base of the organ of Corti that could contribute (1) to the misexpression of the several of the factors that we have described above and (2) to the histological defects observed in the basal region of the organ of Corti in Lmx1a mutant mice.

Atoh1LacZ staining of hair cells and tracer dye studies showing initial segregation of sensory epithelia and their innervation. a Wildtype Atoh1LacZ-stained ear (left anterior, up dorsal, AC anterior crista, DR ductus reuniens, HC horizontal crista, OC organ of Corti, PC posterior crista, S saccule, U utricle, UCSF utriculo-saccular foramen). b An identically stained mutant ear. Note that the cristae are separated at this age and the posterior crista (PC) is not grossly enlarged, although a tiny papilla neglecta (PN) is present. The utricle (U) and saccule (S) are, however, joined. c Higher magnification of the red boxed field in b. The basal termination of the unstained though translucent organ of Corti (OC) overlaps the stained horizontal cristae of the saccule (S). d-g Lipophylic dye tracings in E14.5 mutant ears. The red dye was placed in the solitary nucleus and descending vestibular nucleus. The dye then backfilled collaterals to the vestibular sensory epithelia and sensory neurons of the 7th cranial nerve in the geniculate ganglion (Genggl). The green dye was placed in the (otic) efferent nerve fibers near the floor of the fourth ventricle to both the vestibular and cochlear sensory epithelia (CO cochlea). d Overview in which the green channel dominates. All sensory epithelia receive efferents. e Vestibular afferent (red) and efferent innervation (green) to the anterior crista (AC), horizontal crista (HC), utricle (U), and saccule (S) are almost normal, except for the limited segregation of the saccule from the utricle. f Innervation to the posterior crista (PC) is abnormally widespread and includes a large branch to the papilla neglecta (PN). g Innervation to the basal cochlea (CO) lacks a vestibular (yellow) component. Bars 100 μm
In order to verify the presence of distinct subcompartments within the cochlear-gravistatic epithelium, vestibular afferents were labeled from the brainstem, and vestibule-cochlear efferents were labeled from rhombomere 4, at which point the olivocochlear bundle crosses (Bruce et al. 1997; Fritzsch and Nichols 1993). We had previously shown that, with such double labeling, vestibular and cochlear fibers could be distinguished throughout development (Tessarollo et al. 2004). As expected, brainstem vestibular projections labeled all afferents to the vestibular organs and showed discrete innervation of a large utricular area and a smaller saccular portion (Fig. 9d,e). In contrast, the cochlear region received only efferent fibers and was thus identifiable based on this specific innervation. Whereas anterior and horizontal cristae were normal in their innervation pattern, the posterior crista innervation was expanded by fibers targeting the papilla neglecta.
In summary, hair cell formation in the cristae and papilla neglecta of Lmx1a mutant mice starts as discrete patches, which only later fuse into composite structures. In contrast, the utricle, saccule, and organ of Corti form as a common sensory epithelium. This common epithelium nevertheless shows differences in hair cell organization, maturation, and innervation warranting their labeling as run-on precursors of the utricle, saccule, and organ of Corti.
Unique phenotype of the Lmx1a mutant is foreshadowed by early alterations in Sox2 expression
In the chick (Giraldez 1998), as in mice (Failli et al. 2002), Lmx1a has been shown to be excluded from the neurogenic region of the otic placode. In addition, inhibition of the Notch signaling pathway allows the spread of Lmx1a expression into the neurogenic region (Abello et al. 2007). This is reminiscent of the “non-neural” Tbx1 gene of mice, which when mutated allows neural genes, including Neurog1, to expand their expression domains, and which when overexpressed causes the retreat of neural gene expression (Raft et al. 2004).
To obtain further insights into the molecular organization of supporting cells, we next studied the distribution of Sox2, which is first necessary for prosensory specification (Fritzsch et al. 2006a; Holmberg et al. 2008) and is only later restricted to supporting cells (Kiernan et al. 2005). We therefore sought to determine whether prosensory gene expression would expand into non-neural territory in the Lmx1a null mice.
In E10.5 wildtype animals, Sox2 expression occurred in presumptive sensory areas of the antero-ventral quadrant and in the anlage of the posterior crista (Fig. 10a). This general pattern was retained in the mutant, but the areas of expression were greatly expanded, with more diffuse margins. This was most apparent in the region of the posterior crista (Fig. 10b). By E11.5, however, whereas expression in the wildtype remained strong and neatly confined to future sensory areas, that in the mutant remained broader in the region of the posterior crista (Fig. 10c,d). We noted that segregation of the anterior cristae from the utricle was completed in both wildtype and mutant mice. In contrast, no horizontal crista prosensory patch appeared as a discrete entity in the Lmx1a mutants (Fig. 10c,d). Both wildtype and mutant showed a continuous expression band with focal increases in intensity in the utricle, saccule, and organ of Corti (Fig. 10c,d). By E14.5, all six epithelia were distinct in the wildtype, but the mutant showed a combined anterior and horizontal crista, a combined posterior crista and papilla neglecta (including the extension along the cochlear lateral wall), and the fused cochlear-gravistatic endorgan. The pattern and intensity of Sox2 expression in the cochlear region compared to the contiguous saccular portion were most interesting. In both wildtype and mutant, the cochlear expression was much reduced in intensity compared with that of the saccule. In addition, the cochlear expression was extremely broad in the base of the mutant and much less focused than in the wildtype (Fig. 10e,f). This broader and more diffuse pattern of expression persisted in the mutant base through E18.5. The mutant apex developed focused expression reminiscent of the entire wildtype cochlea (Fig. 10h–k).

Sox2 mRNA expression shows that Lmx1a defines early prosensory patch development (ant anterior, dors dorsal, AC anterior crista, ED HC horizontal crista, OC organ of Corti, PC posterior crista, PN papilla neglecta, S saccule, Spg spiral ganglion, SV stria vascularis, U utricle) a, bSox2 expression is similar, but broader in the mutant without a distinct gap between posterior and anterior prosensory patch expression (star in a, b; see also lower magnification insets). c–e At E11.5, wildtype expression is localizing to individual sensory epithelia (note the separate AC and HC), whereas in the mutant, the AC and HC are joined. Note also the PC/PN extending ventrally toward the cochlea (arrows in d, e, g, i). f, g By E14.5, the wildtype sensory epithelia are clearly segregated (by Lmx1a-expressing epithelia; cf. Fig. 8h), whereas continuities and near continuities are apparent in the mutant. h In the E18.5 wildtype cochlea, Sox2 expression is most intense in the inner pillar cells. i Mutant Sox2 expression identifies the fused utricle-saccule-cochlear and enlarged posterior crista/papilla neglecta. j, k At E18.5, Sox2 expression in the mutant cochlear base (j) is broad and diffuse, whereas that in the apex (k) is, as in the wildtype, focused on the inner pillar cells (P). All whole ears oriented as in a. Bars 100 μm (a-g), 50 μm (h, k)
In summary, the wildtype and Lmx1a dr mutant exhibited major differences in initial hair cell formation as revealed by Atoh1LacZ staining (Fig. 9b), and in prosensory patch formation as revealed by Sox2 expression (Fig. 10g). Whereas prosensory patch formation maintained, from its onset, the fusion/combination of sensory epithelia, hair cell formation initially showed all six sensory epithelia as discrete entities embedded in the unsegregated prosensory patches specified by Sox2. These differences suggest that the initial Atoh1 upregulation and withdrawal of the hair cells from the cell cycle (Matei et al. 2005) and the expression of Sox2 might be under different regulatory control. However, Atoh1 expression and initial cell-cycle exit take place in the Sox2 expression domains. These data also suggest that, in the mutant, the initial expression changes in Sox2 foreshadow the later expansion and disorganization of the hair cells in the mutant cochlea and the overgrowth of the papilla neglecta. Notably, in the wildtype animal, an Lmx1a-expressing caudal constriction separates the posterior crista from the cochlea and thus contributes to the formation of the Lmx1a-expressing ductus reuniens. This constriction remains rudimentary, and the ductus is nonexistent in the Lmx1a mutant, whereas the papilla neglecta overgrows the site preserved by the failure to form the constriction. Thus, in the wildtype, an enlarged papilla neglecta does not form and fuse with the posterior crista, both because Lmx1a expression confines Sox2 expression, and because it creates a non-sensory constriction at the site where the enlarged papilla would form in the mutant.
Discussion
Lmx1a mutants show that non-sensory otic epithelium facilitates ear morphogenesis
Our Lmx1a expression data show a unique association with specific areas of non-sensory otic epithelium in older ears (>E10.5). These Lmx1a-expressing non-sensory epithelia are involved in: (1) separating and constricting the endolymphatic sac from the saccule by the formation of the endolymphatic duct, (2) separating and constricting the utricle from the saccule by forming the utriculo-saccular foramen, and (3) separating and constricting the cochlea from the saccule and posterior crista by forming the ductus reuniens. None of these non-sensory constrictions occur in Lmx1a null mutants, implying that Lmx1a plays a direct or indirect role in all of these morphogenetic events, and that these events are driven in part by Lmx1a expression in the non-sensory epithelia. In addition Lmx1a is expressed in the outer spiral sulcus separating the organ of Corti from the stria vascularis (Figs. 1k, 2a-d). A lack of Lmx1a expression adjacent to developing sensory epithelia might interfere with signaling from the sensory epithelia to govern coordinated morphogenesis of the non-sensory otic epithelia. Alternatively, Lmx1a expression might initiate the secretion of diffusible factors(s) from the non-sensory otic epithelium required for proper sensory epithelia maturation.
The hypothesis that Lmx1a interferes with a sensory epithelium signal is in agreement with the emerging concept that crista epithelia express diffusible factors such as Fgf10 and Bmp4 (bone morphogenetic protein 4) that regulate the growth of the non-sensory part of the vertical canals (Chang et al. 2004b, 2008; Fritzsch et al. 2006b; Pauley et al. 2003). In contrast, the cochlea grows by the intercalation of sensory and non-sensory epithelia (Wang et al. 2006) and does so even when differentiating hair cells never form (Fritzsch et al. 2005a) or when the prosensory anlage of the organ of Corti is disrupted (Kiernan et al. 2005). The data support models that propose more sophisticated molecular interactions and possible feedback loops of sensory and non-sensory epithelia to complete morphogenesis (Chang et al. 2008). These hypotheses conform with the known interactions that occur in the inner ear otic mesenchyme, and that require bilateral signals, only some of which are presently understood (Pirvola et al. 2004).
Overall, Lmx1a mutant dysmorphogenesis is a more exaggerated form of Otx1 null dysmorphogenesis in which the utricle and saccule remain in communication via an open utriculo-saccular foramen, and no ductus reuniens forms (Fritzsch et al. 2001; Morsli et al. 1999). The two phenotypes differ, however, in that the organ of Corti in the Otx1 mutant remains distinct from the saccule in most cases and develops a normal histology. In contrast to Lmx1a expression, Otx1 expression is found in both non-sensory and sensory compartments during ear morphogenesis (Morsli et al. 1999) and thus does not permit the contention that its non-sensory expression is uniquely involved in ear morphogenesis. Several other genes also affect ear morphogenesis (Chang et al. 2004b; Fritzsch et al. 2007). However, the limited characterization of their expression patterns does not allow the distinctions that we can make here for Lmx1a gene expression and function. In addition, the primary action of some of these genes is in the brain, with the ear being secondarily affected. Other genes are overlappingly expressed in both sensory and non-sensory parts of the ear, as in the case with Otx1 and Foxg1 (Pauley et al. 2006; Raft et al. 2004). Based on the Lmx1a expression pattern and defects in null mutants, we show here, for the first time, that a gene expressing a non-diffusible factor exclusively in non-sensory areas of the differentiating ear is essential for aspects of ear morphogenesis and sensory organ histogenesis.
Lmx proteins are known to regulate Fgfs, Wnts, and Bmps in parts of the central nervous system (Adams et al. 2000; Alexandre et al. 2006; Chizhikov and Millen 2004; Guo et al. 2007; Matsunaga et al. 2002; O’Hara et al. 2005). These secreted factors are major players for ear morphogenesis (Chang et al. 2008; Pauley et al. 2003; Riccomagno et al. 2005; Wright and Mansour 2003). Clearly, the expression of Fgf8 is altered in the basal organ of Corti of Lmx1a mutants, showing that disregulation of at least one Fgf indeed occurs in the ear and that the severe dysmorphogenesis might be related to disregulation of both Fgfs and Bmps. Wnts have also been shown to be crucial for ear placode formation (Ohyama et al. 2006) and to play a major role in ear morphogenesis (Riccomagno et al. 2005). Moreover, both Wnts and some Fgfs are secreted by the non-sensory part of the developing ear. Therefore, a Wnt or Fgf agent such as Fgf9 (Pirvola et al. 2004) or Wnt4 (Daudet et al. 2002) might be directly regulated by Lmx1a, and these and other factor(s) released from the Lmx1a-expressing non-sensory areas of the developing ear might regulate crucial aspects of ear morphogenesis and provide a feed-back loop for sensory epithelium development. The absence of the endolymphatic duct in the Lmx1a mutant might certainly be related to the similar absence of the endolymphatic duct in Ffg3 null mice (Hatch et al. 2007) and could explain the ultimate loss of all hair cells in Lmx1a null mice as a consequence of disturbed endolymphatic homeostasis, such as that reported in Foxi1 and Pendrin mutant mice (Blomqvist et al. 2006; Hulander et al. 2003). The absence of an endolymphatic duct does indeed result in the absence of Foxi1 expression (data not shown) and a likely lack of pendrin expression in the missing endolymphatic duct. Further studies are needed to test for ionic disregulation in Lmx1a mutant mice as this might also play a role in adult hair cell loss.
In this context, we should stress that no pigment cells reach the lateral wall of the cochlea but rather accumulate near the radial fibers in Lmx1a null mutant. Wnts, in interaction with Wnt signaling modulators, such as the Dickkopf (DKK) family of secreted factors, set up gradients along which cells migrate. For example, in the skin, DKK interacts with Wnts in a reaction-diffusion mechanism that sets up the spacing of hair follicles (Sick et al. 2006). It also interacts with Wnts in head morphogensis (Lewis et al. 2008). Similar issues of spacing between sensory epithelia are in part the cause of dysmorphogenesis in the Lmx1a mutant ear. The similarity of several aspects of the Otx1 and Lmx1a phenotypes combined with the finding that Dkk1 can rescue the Otx2 phenotype (Kimura-Yoshida et al. 2005; Lewis et al. 2008) implies that Lmx1a plays an unspecified part in Wnt-Otx mediated morphogenesis. Similarly, the close proximity of pigment cells to Lmx1a-expressing areas suggests modulated Wnt signaling. Such signaling may guide pigment epithelial cells to known areas of endolymph production (stria vascularis, dark cells of the utricle, and canal cristae) and resorption (endolymphatic duct). This possible involvement of the Lmx1a protein in the formation and resorption of endolymph, once verified in the Lmx1a mutant ear, could reflect a conserved function of this gene in the ear and hindbrain where Lmx1a is involved in the formation of the cerebrospinal-fluid-secreting choroid plexus (Chizhikov et al. 2006; Elsen et al. 2008). Wnts and their intracellular effector, β-catenin, play important roles in ear formation (Ohyama et al. 2006; Riccomagno et al. 2005). However, more work is needed to elucidate the role of Lmx1a in local otic Wnt secretion (Daudet et al. 2002) and its modulation.
Histological defects of Lmx1a mutants relate to gene misexpression
Our data show that the enlarged anterior and posterior canal cristae of Lmx1a mutants result from the fusion of the horizontal crista and papilla neglecta with the anterior and posterior crista, respectively. Interestingly, the initial upregulation of Atoh1 for the differentiation of hair cells is discrete but is embedded in enlarged or fused Sox2 expression domains (Fig. 10). The hair cell patches fuse into these enlarged epithelia only in late embryonic stages (Figs. 3b, 9b). These data imply that two independent, but topographically related processes focus Sox2 expression to the prosensory patches and Atoh1 expression to the first differentiating hair cells inside the Sox2 expression domains. Obviously, the effect of the absence of Lmx1a expression is more profound on prosensory patch formation, which foreshadows the later phenotype that simply fills in the prosensory domain with differentiated hair cells. Neither focal prosensory patch formation nor focal Atoh1 upregulation is understood at the molecular level (Kiernan et al. 2005; Matei et al. 2005). Like the Lim domain factor tailup in insect sensory development (Biryukova and Heitzler 2005), Lmx1a may counteract the bHLH gene upregulation mediated by Gata3 (Karis et al. 2001) and other factors. However, a complete inventory of Lim-homeodomain factors in ear development is necessary before such multimeric interactions (Bhati et al. 2008a; Matthews and Visvader 2003) can be understood.
We suggest that the common function of the Lim genes Isl/tailup (flies) and Lmx1a (mice) is to define non-sensory cells. This suggestion is in agreement with existing models comparing insect and vertebrate mechanosensory development (Caldwell and Eberl 2002; Fritzsch et al. 2000). However, whereas the sorting of insect sensory and non-sensory cells is reinforced by delta-notch signaling, we argue here that diffusible factor gradients may be altered in the multicellular non-sensory spacers that form between sensory epithelia of mice, complementing the delta-notch function in sensory epithelium segregation (Daudet and Lewis 2005).
In contrast to canal cristae, the wildtype utricle, saccule, and cochlea form as a single elongated epithelium that only segregates later in development (Farinas et al. 2001; Fritzsch et al. 2002; Morsli et al. 1998). Neither prosensory epithelium formation nor hair cell differentiation demonstrate any such segregation in Lmx1a mutants. Segregation appears only during senescence of the adult ear when hair cells die. The blending of vestibular with cochlear hair cell types near the saccular-cochlear transition might be attributable to cell-type mingling across an undetected border. However, this might also suggest that proximity prevents specific factors associated with vestibular and cochlear hair cell differentiation or segregation from functioning normally. In addition, the absence of Lmx1a might change radial gradients of Wnts or other diffusible factors and cause the altered morphology. Certainly, the broad expression of Fgf8 in Lmx1a mutant basal-turn hair cells combined with the wider expression of Sox2 and the absence of Prox1 could result in sophisticated changes in signaling that would lead to the observed dismorphogenesis. Obviously, changing the expression of Fgf8 from a single to multiple rows can affect the differentiation of pillar and Deiters’ cells, as shown for mutations of the Fgfr3 receptor (Puligilla et al. 2007). Hence, the next step is to define those factors that are disregulated by Lmx1a and cause dysmorphogenesis and/or altered histogenesis.
Evolutionary implications
The morphological and histological defects reported here suggest that, in the absence of Lmx1a protein, the mouse ear reverts to a hagfish-like ear consisting of a simple torus with two canal cristae and a single common macula for the gravistatic organs (Fritzsch et al. 2006b). Once DNA sequencing efforts in hagfish have been completed, we should be able to begin a determination of the way that Lmx1a expression differs across vertebrate phyla. Lmx1a might play the same role for the segregation of the common macula into multiple end organs that Otx1 (Fritzsch et al. 2001) and Foxg1 (Pauley et al. 2006) play for the formation of the horizontal canal and horizontal canal crista, respectively, in development and evolution. Further work is needed to show whether the fly Lmx1a ortholog, CG32105, is associated with chordotonal organ development. Alternatively, Lmx1a might be involved in ionic homeostasis, which is critical for the proper function of ear and mechanosensors alike (Fritzsch et al. 2000, 2007; Todi et al. 2004; Walker et al. 2000). Irrespetive of the details, our results suggest that local Lmx1a expression is a major player in ear morpho- and histogenesis, and that this function is probably conserved at the level of interacting modules of transcription factors.
References
Abello G, Khatri S, Giraldez F, Alsina B (2007) Early regionalization of the otic placode and its regulation by the Notch signaling pathway. Mech Dev 124:631–645
Adam J, Myat A, Le Roux I, Eddison M, Henrique D, Ish-Horowicz D, Lewis J (1998) Cell fate choices and the expression of Notch, Delta and Serrate homologues in the chick inner ear: parallels with Drosophila sense-organ development. Development 125:4645–4654
Adams KA, Maida JM, Golden JA, Riddle RD (2000) The transcription factor Lmx1b maintains Wnt1 expression within the isthmic organizer. Development 127:1857–1867
Alenina N, Bashammakh S, Bader M (2006) Specification and differentiation of serotonergic neurons. Stem Cell Rev 2:5–10
Alexandre P, Bachy I, Marcou M, Wassef M (2006) Positive and negative regulations by FGF8 contribute to midbrain roof plate developmental plasticity. Development 133:2905–2913
Asmar J, Biryukova I, Heitzler P (2008) Drosophila dLMO-PA isoform acts as an early activator of achaete/scute proneural expression. Dev Biol 316:487–497
Bermingham NA, Hassan BA, Wang VY, Fernandez M, Banfi S, Bellen HJ, Fritzsch B, Zoghbi HY (2001) Proprioceptor pathway development is dependent on Math1. Neuron 30:411–422
Bermingham-McDonogh O, Oesterle EC, Stone JS, Hume CR, Huynh HM, Hayashi T (2006) Expression of Prox1 during mouse cochlear development. J Comp Neurol 496:172–186
Bhati M, Lee C, Nancarrow AL, Lee M, Craig VJ, Bach I, Guss JM, Mackay JP, Matthews JM (2008a) Implementing the LIM code: the structural basis for cell type-specific assembly of LIM-homeodomain complexes. EMBO J 27:2018–2029
Bhati M, Lee M, Nancarrow AL, Bach I, Guss JM, Matthews JM (2008b) Crystallization of an Lhx3-Isl1 complex. Acta Crystallogr Sect F Struct Biol Cryst Commun 64:297–299
Biryukova I, Heitzler P (2005) The Drosophila LIM-homeo domain protein Islet antagonizes pro-neural cell specification in the peripheral nervous system. Dev Biol 288:559–570
Blomqvist SR, Vidarsson H, Soder O, Enerback S (2006) Epididymal expression of the forkhead transcription factor Foxi1 is required for male fertility. EMBO J 25:4131–4141
Bruce LL, Kingsley J, Nichols DH, Fritzsch B (1997) The development of vestibulocochlear efferents and cochlear afferents in mice. Int J Dev Neurosci 15:671–692
Caldwell JC, Eberl DF (2002) Towards a molecular understanding of Drosophila hearing. J Neurobiol 53:172–189
Chang W, Brigande JV, Fekete DM, Wu DK (2004a) The development of semicircular canals in the inner ear: role of FGFs in sensory cristae. Development 131:4201–4211
Chang W, Cole LK, Cantos R, Wu DK (2004b) Molecular genetics of vestibular organ development. In: Highstein SM, Fay RR, Popper AN (eds) The vestibular system, vol 19. Springer, New York, pp 11–56
Chang W, Lin Z, Kulessa H, Hebert J, Hogan BL, Wu DK (2008) Bmp4 is essential for the formation of the vestibular apparatus that detects angular head movements. PLoS Genet 4:e1000050
Chen P, Johnson JE, Zoghbi HY, Segil N (2002) The role of Math1 in inner ear development: uncoupling the establishment of the sensory primordium from hair cell fate determination. Development 129:2495–2505
Chizhikov VV, Millen KJ (2004) Control of roof plate development and signaling by Lmx1b in the caudal vertebrate CNS. J Neurosci 24:5694–5703
Chizhikov V, Steshina E, Roberts R, Ilkin Y, Washburn L, Millen KJ (2006) Molecular definition of an allelic series of mutations disrupting the mouse Lmx1a (dreher) gene. Mamm Genome 17:1025–1032
Daudet N, Lewis J (2005) Two contrasting roles for Notch activity in chick inner ear development: specification of prosensory patches and lateral inhibition of hair-cell differentiation. Development 132:541–551
Daudet N, Ripoll C, Moles JP, Rebillard G (2002) Expression of members of Wnt and Frizzled gene families in the postnatal rat cochlea. Brain Res Mol Brain Res 105:98–107
Deng M, Pan L, Xie X, Gan L (2006) Differential expression of LIM domain-only (LMO) genes in the developing mouse inner ear. Gene Expr Patterns 6:857–863
Deol MS (1964) The origin of the abnormalities of the inner ear in dreher mice.. J Embryol Exp Morph 12:727–733
Deol MS (1983) Development of auditory and vestibular systems in mutant mice. In: Romand R (eds) Development of vestibular and auditory systems. Academic Press, New York, pp 309–333
Elsen GE, Choi LY, Millen KJ, Grinblat Y, Prince VE (2008) Zic1 and Zic4 regulate zebrafish roof plate specification and hindbrain ventricle morphogenesis. Dev Biol 314:376–392
Failli V, Bachy I, Retaux S (2002) Expression of the LIM-homeodomain gene Lmx1a (dreher) during development of the mouse nervous system. Mech Dev 118:225–228
Farinas I, Jones KR, Tessarollo L, Vigers AJ, Huang E, Kirstein M, de Caprona DC, Coppola V, Backus C, Reichardt LF, Fritzsch B (2001) Spatial shaping of cochlear innervation by temporally regulated neurotrophin expression. J Neurosci 21:6170–6180
Fekete DM, Wu DK (2002) Revisiting cell fate specification in the inner ear. Curr Opin Neurobiol 12:35–42
Fritzsch B (2003) The ear of Latimeria chalumnae revisited. Zoology (Jena) 106:243–248
Fritzsch B, Nichols DH (1993) DiI reveals a prenatal arrival of efferents at the differentiating otocyst of mice. Hear Res 65:51–60
Fritzsch B, Wake MH (1988) The inner ear of gymnophione amphibians and its nerve supply: a comparative study of regressive events in a complex sensory system. Zoomorphology 108:210–217
Fritzsch B, Beisel KW, Bermingham NA (2000) Developmental evolutionary biology of the vertebrate ear: conserving mechanoelectric transduction and developmental pathways in diverging morphologies. Neuroreport 11:R35–R44
Fritzsch B, Signore M, Simeone A (2001) Otx1 null mutant mice show partial segregation of sensory epithelia comparable to lamprey ears. Dev Genes Evol 211:388–396
Fritzsch B, Beisel KW, Jones K, Farinas I, Maklad A, Lee J, Reichardt LF (2002) Development and evolution of inner ear sensory epithelia and their innervation. J Neurobiol 53:143–156
Fritzsch B, Matei VA, Nichols DH, Bermingham N, Jones K, Beisel KW, Wang VY (2005a) Atoh1 null mice show directed afferent fiber growth to undifferentiated ear sensory epithelia followed by incomplete fiber retention. Dev Dyn 233:570–583
Fritzsch B, Muirhead KA, Feng F, Gray BD, Ohlsson-Wilhelm BM (2005b) Diffusion and imaging properties of three new lipophilic tracers, NeuroVuetrade mark Maroon, NeuroVuetrade mark Red and NeuroVuetrade mark Green and their use for double and triple labeling of neuronal profile. Brain Res Bull 66:249–258
Fritzsch B, Beisel KW, Hansen LA (2006a) The molecular basis of neurosensory cell formation in ear development: a blueprint for hair cell and sensory neuron regeneration? Bioessays 28:1181–1193
Fritzsch B, Pauley S, Beisel KW (2006b) Cells, molecules and morphogenesis: the making of the vertebrate ear. Brain Res 1091:151–171
Fritzsch B, Beisel KW, Pauley S, Soukup G (2007) Molecular evolution of the vertebrate mechanosensory cell and ear. Int J Dev Biol 51:663–678
Fritzsch B, Srinivasan RS, Harvey NL, Nichols DH, Oliver G (2008) Canal cristae growth and fiber extension to the outer hair cells require Prox1 activity. Dev Dyn (in press)
Giraldez F (1998) Regionalized organizing activity of the neural tube revealed by the regulation of lmx1 in the otic vesicle. Dev Biol 203:189–200
Guo C, Qiu HY, Huang Y, Chen H, Yang RQ, Chen SD, Johnson RL, Chen ZF, Ding YQ (2007) Lmx1b is essential for Fgf8 and Wnt1 expression in the isthmic organizer during tectum and cerebellum development in mice. Development 134:317–325
Hatch EP, Noyes CA, Wang X, Wright TJ, Mansour SL (2007) Fgf3 is required for dorsal patterning and morphogenesis of the inner ear epithelium. Development 134:3615–3625
Hertzano R, Dror AA, Montcouquiol M, Ahmed ZM, Ellsworth B, Camper S, Friedman TB, Kelley MW, Avraham KB (2007) Lhx3, a LIM domain transcription factor, is regulated by Pou4f3 in the auditory but not in the vestibular system. Eur J Neurosci 25:999–1005
Holmberg J, Hansson E, Malewicz M, Sandberg M, Perlmann T, Lendahl U, Muhr J (2008) SoxB1 transcription factors and Notch signaling use distinct mechanisms to regulate proneural gene function and neural progenitor differentiation. Development 135:1843–1851
Hulander M, Kiernan AE, Blomqvist SR, Carlsson P, Samuelsson EJ, Johansson BR, Steel KP, Enerback S (2003) Lack of pendrin expression leads to deafness and expansion of the endolymphatic compartment in inner ears of Foxi1 null mutant mice. Development 130:2013–2025
Hunter CS, Rhodes SJ (2005) LIM-homeodomain genes in mammalian development and human disease. Mol Biol Rep 32:67–77
Karis A, Pata I, Doorninck JH van, Grosveld F, Zeeuw CI de, Caprona D de, Fritzsch B (2001) Transcription factor GATA-3 alters pathway selection of olivocochlear neurons and affects morphogenesis of the ear. J Comp Neurol 429:615–630
Kelley MW (2006) Regulation of cell fate in the sensory epithelia of the inner ear. Nat Rev Neurosci 7:837–849
Kiernan AE, Nunes F, Wu DK, Fekete DM (1997) The expression domain of two related homeobox genes defines a compartment in the chicken inner ear that may be involved in semicircular canal formation. Dev Biol 191:215–229
Kiernan AE, Pelling AL, Leung KK, Tang AS, Bell DM, Tease C, Lovell-Badge R, Steel KP, Cheah KS (2005) Sox2 is required for sensory organ development in the mammalian inner ear. Nature 434:1031–1035
Kimura-Yoshida C, Nakano H, Okamura D, Nakao K, Yonemura S, Belo JA, Aizawa S, Matsui Y, Matsuo I (2005) Canonical Wnt signaling and its antagonist regulate anterior-posterior axis polarization by guiding cell migration in mouse visceral endoderm. Dev Cell 9:639–650
Lee S, Lee B, Joshi K, Pfaff SL, Lee JW, Lee SK (2008) A regulatory network to segregate the identity of neuronal subtypes. Dev Cell 14:877–889
Lewis ER, Leverenz EL, Bialek WS (1985) The vertebrate inner ear. CRC Press, Boca Raton
Lewis SL, Khoo PL, De Young RA, Steiner K, Wilcock C, Mukhopadhyay M, Westphal H, Jamieson RV, Robb L, Tam PP (2008) Dkk1 and Wnt3 interact to control head morphogenesis in the mouse. Development 135:1791–1801
Lillevali K, Haugas M, Matilainen T, Pussinen C, Karis A, Salminen M (2006) Gata3 is required for early morphogenesis and Fgf10 expression during otic development. Mech Dev 123:415–429
Ma Q, Chen Z, Barco Barrantes I del, Pompa JL de la, Anderson DJ (1998) Neurogenin1 is essential for the determination of neuronal precursors for proximal cranial sensory ganglia. Neuron 20:469–482
Ma Q, Anderson DJ, Fritzsch B (2000) Neurogenin 1 null mutant ears develop fewer, morphologically normal hair cells in smaller sensory epithelia devoid of innervation. J Assoc Res Otolaryngol 1:129–143
Manzanares M, Trainor PA, Ariza-McNaughton L, Nonchev S, Krumlauf R (2000) Dorsal patterning defects in the hindbrain, roof plate and skeleton in the dreher (dr(J)) mouse mutant. Mech Dev 94:147–156
Matei V, Pauley S, Kaing S, Rowitch D, Beisel KW, Morris K, Feng F, Jones K, Lee J, Fritzsch B (2005) Smaller inner ear sensory epithelia in Neurog1 null mice are related to earlier hair cell cycle exit. Dev Dyn 234:633–650
Matei VA, Feng F, Pauley S, Beisel KW, Nichols MG, Fritzsch B (2006) Near-infrared laser illumination transforms the fluorescence absorbing X-Gal reaction product BCI into a transparent, yet brightly fluorescent substance. Brain Res Bull 70:33–43
Matsunaga E, Katahira T, Nakamura H (2002) Role of Lmx1b and Wnt1 in mesencephalon and metencephalon development. Development 129:5269–5277
Matthews JM, Visvader JE (2003) LIM-domain-binding protein 1: a multifunctional cofactor that interacts with diverse proteins. EMBO Rep 4:1132–1137
Millonig JH, Millen KJ, Hatten ME (2000) The mouse Dreher gene Lmx1a controls formation of the roof plate in the vertebrate CNS. Nature 403:764–769
Morsli H, Choo D, Ryan A, Johnson R, Wu DK (1998) Development of the mouse inner ear and origin of its sensory organs. J Neurosci 18:3327–3335
Morsli H, Tuorto F, Choo D, Postiglione MP, Simeone A, Wu DK (1999) Otx1 and Otx2 activities are required for the normal development of the mouse inner ear. Development 126:2335–2343
O’Hara FP, Beck E, Barr LK, Wong LL, Kessler DS, Riddle RD (2005) Zebrafish Lmx1b.1 and Lmx1b.2 are required for maintenance of the isthmic organizer. Development 132:3163–3173
Ohyama T, Mohamed OA, Taketo MM, Dufort D, Groves AK (2006) Wnt signals mediate a fate decision between otic placode and epidermis. Development 133:865–875
Ohyama T, Groves AK, Martin K (2007) The first steps towards hearing: mechanisms of otic placode induction. Int J Dev Biol 51:463–472
Pauley S, Wright TJ, Pirvola U, Ornitz D, Beisel K, Fritzsch B (2003) Expression and function of FGF10 in mammalian inner ear development. Dev Dyn 227:203–215
Pauley S, Lai E, Fritzsch B (2006) Foxg1 is required for morphogenesis and histogenesis of the mammalian inner ear. Dev Dyn 235:2470–2482
Pierce ML, Weston MD, Fritzsch B, Gabel HW, Ruvkun G, Soukup GA (2008) MicroRNA-183 family conservation and ciliated neurosensory organ expression. Evol Dev 10:106–113
Pirvola U, Ylikoski J, Trokovic R, Hebert J, McConnell S, Partanen J (2002) FGFR1 is required for the development of the auditory sensory epithelium. Neuron 35:671–685
Pirvola U, Zhang X, Mantela J, Ornitz DM, Ylikoski J (2004) Fgf9 signaling regulates inner ear morphogenesis through epithelial-mesenchymal interactions. Dev Biol 273:350–360
Puligilla C, Feng F, Ishikawa K, Bertuzzi S, Dabdoub A, Griffith AJ, Fritzsch B, Kelley MW (2007) Disruption of fibroblast growth factor receptor 3 signaling results in defects in cellular differentiation, neuronal patterning, and hearing impairment. Dev Dyn 236:1905–1917
Radde-Gallwitz K, Pan L, Gan L, Lin X, Segil N, Chen P (2004) Expression of Islet1 marks the sensory and neuronal lineages in the mammalian inner ear. J Comp Neurol 477:412–421
Raft S, Nowotschin S, Liao J, Morrow BE (2004) Suppression of neural fate and control of inner ear morphogenesis by Tbx1. Development 131:1801–1812
Riccomagno MM, Takada S, Epstein DJ (2005) Wnt-dependent regulation of inner ear morphogenesis is balanced by the opposing and supporting roles of Shh. Genes Dev 19:1612–1623
Sick S, Reinker S, Timmer J, Schlake T (2006) WNT and DKK determine hair follicle spacing through a reaction-diffusion mechanism. Science 314:1447–1450
Sienknecht UJ, Fekete DM (2008) Comprehensive Wnt-related gene expression during cochlear duct development in chicken. J Comp Neurol 510:378–395
Tessarollo L, Coppola V, Fritzsch B (2004) NT-3 replacement with brain-derived neurotrophic factor redirects vestibular nerve fibers to the cochlea. J Neurosci 24:2575–2584
Todi SV, Sharma Y, Eberl DF (2004) Anatomical and molecular design of the Drosophila antenna as a flagellar auditory organ. Microsc Res Tech 63:388–399
Van Esch H, Devriendt K (2001) Transcription factor GATA3 and the human HDR syndrome. Cell Mol Life Sci 58:1296–1300
Walker RG, Willingham AT, Zuker CS (2000) A Drosophila mechanosensory transduction channel. Science 287:2229–2234
Wang J, Hamblet NS, Mark S, Dickinson ME, Brinkman BC, Segil N, Fraser SE, Chen P, Wallingford JB, Wynshaw-Boris A (2006) Dishevelled genes mediate a conserved mammalian PCP pathway to regulate convergent extension during neurulation. Development 133:1767–1778
Wright TJ, Mansour SL (2003) Fgf3 and Fgf10 are required for mouse otic placode induction. Development 130:3379–3390
Acknowledgement
We are grateful to Jennifer Kersigo, Anne Lindgren, Yuriko Mishima, and Amanda Branch for technical assistance and collection of mouse embryos, to Dr. Garret Soukup, Marsha Pierce, and Jason Pecka for help in designing and preparing primers, to Drs. Engel, Ornitz, Hogan and Chea for providing plasmids with probes, and to Dr. Huda Zoghbi for mice.
Author information
Authors and Affiliations
Corresponding author
Additional information
This work was supported by grants from the NCRR/COBRE (P20 RR 018788; D.H.N.) and NIH (RO1 DC 005590; B.F.). Parts of this investigation were conducted in a facility constructed with support of a Research Facilities Improvement Program Grant from the National Center for Research Resources, National Institutes of Health. We acknowledge the use of the confocal microscope facility of the NCCB, supported by EPSCoR EPS-0346476 (CFD 47.076), and of the University of Nebraska microarray facility, supported by NCRR/COBRE.
Rights and permissions
About this article
Cite this article
Nichols, D.H., Pauley, S., Jahan, I. et al. Lmx1a is required for segregation of sensory epithelia and normal ear histogenesis and morphogenesis. Cell Tissue Res 334, 339–358 (2008). https://doi.org/10.1007/s00441-008-0709-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00441-008-0709-2