
Abstract
This study aimed to demonstrate the ability of Argulus japonicus to infect a wide range of freshwater fishes, as well as to understand the effects of fish origin and host body size on the incidence of A. japonicus. Samples of cultured and wild fish were collected randomly from July 2010 to March 2013, using angling, long-lining, gill-netting, and trapping from rivers and fish farms in Guangdong province, South China. Eight fish species were found to be heavily infected including the common carp, the goldfish, the black carp, the silver carp, the brown trout, the rainbow trout, the mandarin fish, and the perch. Furthermore, the black carp, the brown trout, and the mandarin fish were recorded as new hosts for the first time. During the present study, a total of 2,271 fishes were examined, out of which 712 fishes were found to be infected by a total of 1,443 A. japonicus. Abundance and intensity of A. japonicus infection were significantly influenced by origin of fishes (cultured and wild) and total length (class I, <250 mm; class II, 250–350 mm; and class III, >350 mm) of fish species, whereas varied impacts on prevalence of infection were observed. The correlation between total length of fishes and prevalence of A. japonicus infection was variable, where no significant correlation was observed in the black carp, the silver carp, the mandarin fish, and the perch. In spite of the weak negative correlation between body size of the silver carp and prevalence of infection, A. japonicus was the most abundant and intensive in the silver carp. Thus, aquaculturists should pay particular attention to the control of these fish lice due to its host biodiversity.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Argulus japonicus has the greatest economic impact of any parasites in cultured fish and is also a threat to wild fish and increase the susceptibility of its host to secondary infections (Bandilla et al. 2006; Walker et al. 2011b). They result in economic losses not only in mortality, but also from treatment expenses and growth reduction during and after the outbreak of disease, and this militate against expansion of aquaculture (Cowx 1992; Omeji et al. 2011). The number of parasites necessary to cause harm to a host varies considerably with species, size of host, and its health status (Carpenter et al. 2001). The relationship between fish and parasites in the natural environment is an important way to understand pathologies in hosts and are also relevant to pisciculture (Tavares–Dias et al. 2000). Direct effects, such as host impairment, can induce indirect consequences (Crowden and Broom 1980; Milinski 1984), greater vulnerability to predators (Arme and Owen 1967), and decreased resistance to environmental stress (Lewis and Hettler 1968).
Host specificity is resulting from vicariance events or a long co-evolutionary history between some parasites and their host (Poulin et al. 2011). Argulid parasites exhibit low host specificity (Walker et al. 2011a); many authors as well have commented on the lack of specificity of argulid parasites, sharing the opinion that individual species from this group can infect a wide range of host species (Kearn 2004; Walker et al. 2004; Alsarakibi et al. 2012).
The objectives of current work were to investigate the host biodiversity of A. japonicus, as well as to compare the prevalence, abundance, and intensity of A. japonicus on wild and cultured fish, in addition to study the relationship between host size (total length) and the occurrence of A. japonicus in freshwater fish of Guangdong province, China.
Materials and methods
Sample collection
Samples of cultured and wild fish were collected randomly from July 2010 to March 2013, using angling, long-lining, gill-netting, and trapping from rivers and fish farms in Guangdong province, South China. After capture, the fish were immediately labeled, and total length (to the nearest millimeters) was recorded and categorized into three classes (class I, <250 mm; class II, 250–350 mm; and class III, >350 mm). The external surface of each fish was examined thoroughly for ectoparasites using a hand lens. Argulus sp. was detected and detached individually from infected fish. A. japonicus specimens were identified according to Wadeh et al. (2008) and counted from each sample separately. Parasite prevalence, abundance, and mean intensity were chosen as comparative parameters based on the definition in Bush et al. (1997).
Statistical analysis
Effect of fish origin (wild and cultured) on abundance and intensity of A. japonicus infection was examined using Mann–Whitney U test. The difference in parasite prevalence was tested by using Chi-square tests (χ 2). Kruskal–Wallis test was applied to test the difference in infection among host length classes. Spearman’s correlation (r) was used to examine relationships between host length and prevalence, abundance, and intensity of infection for each fish species. The differences were considered significant at P < 0.05. All the statistical tests were performed by using Statistica 10.0 for Windows, StatSoft Inc.
Results
Host biodiversity
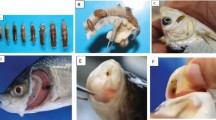
Eight fish species, representing four families, encountered A. japonicus (Fig. 1) during the sampling period. There were four species of Cyprinidae, the common carp Cyprinus carpio (L. 1758), the goldfish Carassius auratus (L. 1758), the black carp Mylopharyngodon piceus (Richardson 1846), and the silver carp Hypophthalmichthys molitrix (Valenciennes 1844). The brown trout Salmo trutta (L. 1758) and the rainbow trout Oncorhynchus mykiss (Walbaum 1792) represented Salmonidae, the mandarin fish Siniperca chuatsi (Basilewsky 1855), and the perch Perca fluviatilis (L. 1758) represented Percichthyidae and Percidae, respectively. Caught fish species were unevenly presented in our samples, indicating that there are probably greater numbers of some species than others within the resident fish community. Among them, the A. japonicus infection of the black carp M. piceus, the brown trout S. trutta, and the mandarin fish S. chuatsi is the first report.

Light micrograph of a female specimen of A. japonicus. a dorsal view; a antenna, ce compined eye, fm first maxialla, sm second maxilla, ra resparatory area, ps proboscis. b ventral view; ne nupilus eye, ab abdomen
Parasitism of A. japonicus
A total of 2,271 individual fishes were examined. Seven hundred twelve of these fishes (31.4 %) were infected with a total of 1,443 A. japonicus. The number of fish per species was varied from 168 to 425, a reflection of both relative abundances and differences in distribution of different fish species. Variation in prevalence of infection was observed between fish species examined, where A. japonicus was most prevalent on the mandarin fish S. chuatsi (42.1 %), whereas the prevalence of infection on brown trout S. trutta was the lowest (20 %). Overall, prevalence of infection was varied significantly within fish species examined (χ 2 = 66.32, P < 0.01). Furthermore, the highest abundance (1.70 ± 0.11) and the highest intensity (4.07 ± 0.14) were found on the silver carp H. molitrix. Abundance (χ 2 = 115.277, P < 0.01) and intensity (X 2 = 159.367, P < 0.01) of A. japonicus infection was significantly different between fish species captured (Table 1).
Overall, prevalence of infection was significantly higher in cultured fish (n = 487, 68.4 %) than in wild fish (n = 218, 31.6 %) (Z adj = 6.630, P < 0.01) (Fig. 2a). Nevertheless, prevalence of infection was significantly influenced by fish origin within fish species, with an exception of the silver carp H. molitrix which showed insignificant influence. However, A. japonicus was most prevalent in cultured (46.2 %) and wild (33.3 %) Mandarin fish S. chuatsi, whereas the lowest prevalence was observed in S. trutta in both cultured (22.5 %) and wild (16.7 %) fish (Fig. 2b). Mean abundance of cultured fishes (0.89 ± 0.04) was significantly higher than that in wild fishes (0.53 ± 0.04) (Z adj = 7.303, P < 0.01); significant differences as well were found on intensity of infection between cultured (2.18 ± 0.06) and wild fishes (1.75 ± 0.08) (Z adj = 4.374, P < 0.01).

Differences between prevalence of infection in cultured and wild fish (a) and between fish species (b) collected from freshwater of Guangdong province, China, during July 2010 to March 2013
However, A. japonicus was most abundant on cultured (1.84 ± 0.12) and wild (1.20 ± 0.13) silver carp H. molitrix, whereas the lowest abundance was observed on S. trutta in both cultured (0.55 ± 0.06) and wild (0.35 ± 0.10) fish combined. It is noteworthy that intensity of infection was also the highest in both cultured (4.17 ± 0.16) and wild (3.73 ± 0.14) silver carp H. molitrix, whilst the brown trout O. mykiss showed the lowest intensity (Table 2). Spearman’s correlation coefficient showed that prevalence, mean abundance, and intensity of A. japonicus infection were varied within fish species (Table 3).
In the present study, the relationship between total length of fishes and prevalence of infection was varied, generally; smaller-sized fishes were less infected than the larger-sized fishes (class III, 48.0 %; class II, 34.2 %; and class I, 31.7 %) (H = 41.033, P < 0.01). Moreover, mean abundance and mean intensity of A. japonicus infection were significantly varied by host length class, being higher in the length class III compared with class II (H = 152.491, P < 0.01) and class I (H = 130.390, P < 0.01), respectively (Fig. 3a and b). It is noteworthy that A. japonicus was more prevalent within class I new hosts, the silver carp H. molitrix and mandarin fish S. chuatsi. Mean parasitological indices (prevalence, abundance, and intensity) within fish species examined are shown in Table 4.

Abundance (a) and intensity (b) of A. japonicus infection within hosts’ length classes of freshwater fishes collected from Guangdong province, China, during July 2010 to March 2013
Based on all fishes examined, the prevalence was positively correlated with total host length of six fish species, whilst two new hosts, the silver carp H. molitrix and the mandarin fish S. chuatsi, showed negative correlation. Moreover, total host length was positively correlated with abundance and intensity of all fish species examined, as well; most correlations were significant as shown in Table 5.
Discussion
A range of different freshwater ecosystems around Guangdong province were investigated during the period studied, and on each sampling trip, several sampling methods were employed to ensure that any bias in fish species caught could not be attributed to sampling technique. Our data showed eight fish species representing four families were heavily infected by A. japonicus, indicating this fish louse is a generalist parasite, which is in agreement with many authors who have stated that argulid parasites infect a wide range of freshwater fish hosts (Kearn 2004; Walker et al. 2004, 2011a; Nagasawa 2009; Alsarakibi et al. 2012).
Regardless of fish species variety in Guangdong province, cyprinid were most frequent fish captured, as well as the infected fish during our study. Among more than 40 fish species reported worldwide as host of A. japonicus, there are up to 31 species belonged to Cyprinidae (Alsarakibi, unpublished data), thus, it makes sense to say that A. japonicus prefer cyprinid. However, the available information indicates that fish responds to parasite infections by activating different innate and adaptive immune mechanism. Studies have been mainly focused on some innate activities, most of which were demonstrated to be involved in the immune response to parasitoses (Alvarez–Pellitero 2008). Generally, fish immune response to A. japonicus is varied within species, which is initially restricted to the site of infection but extended to a generalized response throughout the skin as a whole organ at a later stage of the infection (Forlenza et al. 2008). However, it could be attributed to the fact that adult argulids are quite mobile, being able to glide over the surface of their hosts with relative ease using their maxillary suckers (Kearn 2004), demonstrating the ability of A. japonicus to infect a wide range of fish species.
The current study presents first record of A. japonicus infected the black carp, the brown trout, and the mandarin fish. It is possibly due to either infection transmission between fish hosts in same aquatic system or introduction of A. japonicus with introduced infected fish, especially under conditions of culture. Three new hosts belonged to three different families. However, comparison of the prevalence of infection between these hosts revealed that prevalence was independently associated with fish species; moreover, abundance and intensity of those three new hosts imply that fish species is not a key factor of A. japonicus infection.
Obviously, the prevalence, mean abundance, and mean intensity of A. japonicus were higher in cultured fish than in wild fish, which might strongly be attributed to the fish population in cultured facilities compared with that in wild. In addition, under cultured conditions, the over-stocking, inadequate nutrition, and poor water circulation can cause parasitic outbreaks (Khan 2009). This is consistent with the findings of Taylor et al. (2006, 2009) and McPherson et al. (2012), which concluded that slow stock turnover was an important key in determining the abundance of the Argulus sp., as well as the threshold at which it could survive. Furthermore, the changeable biotic and abiotic factors in the aquatic system may strongly impact on the fish ectoparasite outbreak (Walker et al. 2004; Violante–Gonzalez et al. 2009; Ibrahim 2012; Alsarakibi et al. 2012). In the case of new hosts recorded, A. japonicus infections were higher than expected; it may be attributed to host immune responses.
Parasite abundance can vary by more than one order of magnitude among host populations (Poulin 2006); however, our data on abundance and intensity hint towards a trend of silver carps and black carps being favored as hosts by A. japonicus. Notwithstanding this finding, the silver carp has not been considered traditionally as potential hosts of A. japonicus, where only Kimura (1970) and Qizhong and Chenglin (1994) have reported infections of A. japonicus on sliver carp, which were fewer infections as compared with other fish species. For clarification, we offer a plausible suggestion, similar to the host fish and other animals which have adaptive immune mechanisms; this parasite has also the ability to develop offensive mechanisms; this capability could be attributed to genetic changes, where the host selection in this ectoparasite does not depend exclusively on the features of the host/parasite but also on the surrounding conditions. On the other hand, Mikheev et al. (2007) have suggested that the innate ontogenetic shift in host preference maintains the major part of the parasite population on its principal host, ensuring successful reproduction within suitable habitats. Generally, if the parasite and various host fish species were evenly distributed within the water body, the parasite distribution should be related to host population (Walker et al. 2008). However, in our study, A. japonicus distribution within the host community does not appear to be related with the number of available hosts from each fish species.
Only four fish species showed a significant relationship between their prevalence and total host length, while mean abundance and mean intensity of A. japonicus showed a positive significant correlation with total host length in all captured fish species. Generally, parasite loads on fish have been shown to be influenced by host size (Kabata 1981; Grutter 1994). The general trend is that larger individuals have higher physical (ventilation volume) and chemical (mucus) stimuli, which increase their attractiveness to parasites (Lo et al. 1998; Tucker et al. 2002). The reason may be partly related to the host age, older hosts having longer to accumulate parasites (Bush et al. 2003; Zander 2004). However, some fish species appeared to tolerate multiple number of parasites without exhibiting debility, whilst young fish were infected with fewer parasites than older fish, and in addition, prevalence of the infection and mean abundance increased with the host length (Khan 2012). It also may be partly related to the surface area of host as a larger host has a greater surface area, making it easier for parasites to locate and attach to them (Walker et al. 2007). The negative correlation between the total length of silver carp and its prevalence may be the result of host immune responses and likewise of the insignificant correlation between three other fish species and total length. Similar to our finding, Bakke et al. (2002) have reported negative correlation between the population of Gyrodactylus salaries and age of Salmonidae.
Conclusion
A. japonicus infects a wide range of freshwater fish. Cyprinids are preferred species of A. japonicus, where silver carps and black carps were the most susceptible. Cultured fish are more infected than wild fish. Furthermore, A. japonicus were more abundant in larger-sized fish than smaller-sized fish. Therefore, fish population and body size are important keys in determining the abundance and intensity of A. japonicus. The black carp, the brown trout, and the mandarin fish were three new hosts found in the current study. Hence, aquaculturists should pay particular attention to the control of fish lice due to their economic importance.
References
Alsarakibi M, Wadeh H, Li GQ (2012) Freshwater abiotic components’ impact on the viability of fish lice, Argulus sp., in Guangdong province, China. Parasitol Res 111(1):331–339
Alvarez–Pellitero P (2008) Fish immunity and parasite infections: from innate immunity to immunoprophylactic prospects. Vet Immunol Immunopathol 126:171–198
Arme C, Owen RW (1967) Infections of the three spined stickleback, Gasterosteous aculeatus L. with the plerocercoid larvae of Schistocephalus solidus (Muller, 1776), with special reference to pathological effects. Parasitology 57:301–314
Bakke TA, Harris PD, Cable J (2002) Host specificity dynamics: observations on Gyrodactylids monogeneans. Int J Parasitol 32:281–308
Bandilla M, Valtonen ET, Suomalainen LR, Aphalo PJ, Hakalahti T (2006) A link between ectoparasite infection and susceptibility to bacterial disease in rainbow trout. Int J Parasitol 36:987–991
Bush AO, Lafferty KD, Lotz JM, Shostak AW (1997) Parasitology meets ecology on its own terms: Margolis et al. revisited. J Parasitol 83:575–583
Bush AO, Fernandez J, Esch GW, Seed J (2003) Parasitism: the diversity and ecology of animal parasites. Cambridge University, Cambridge, England
Carpenter JW, Mashima TY, Rupiper DJ (2001) Exotic animal formulary. W.B. Saunders Company, St. Louis
Cowx IG (1992) Aquaculture development in Africa: training and reference manual for aquaculture extensionists. Food production and rural development Division, London
Crowden AE, Broom DM (1980) Effects of the eyefluke, Diplostomum spathaceum, on the behaviour of dace (Leuciscus leuciscus). Anim Behav 28:287–294
Forlenza M, Walker PD, de Vries BJ, Bonga SEW, Wiegertjes GF (2008) Transcriptional analysis of the common carp (Cyprinus carpio L.) immune response to the fish louse Argulus japonicus Thiele (Crustacea: Branchiura). Fish Shellfish Immunol 25:76–83
Grutter AS (1994) Spatial and temporal variations of the ectoparasites of seven reef fish species from Lizard Island and Heron Island, Australia. Mar Ecol Prog Ser 115:21–30
Ibrahim MM (2012) Variation in parasite infracommunies of Tilapia zillii in relation to some biotic and abiotic factors. Int J Zool Res 8:59–70
Kabata Z (1981) Copepoda (Crustacea) parasitic on fishes: problems and perspectives. Adv Parasitol 19:1–71
Kearn GC (2004) Leeches, lice and lampreys: a natural history of skin and gill parasites of fishes. Springer, Dordrecht, the Netherlands
Khan RA (2009) Parasites causing disease in wild and cultured fish in Newfoundland. Icel Agric Sci 22:29–35
Khan RA (2012) Host–parasite interactions in some fish species. J Parasitol Res, article ID 237280, 7 pages. doi:10.1155/2012/237280
Kimura S (1970) Notes on the reproduction of water lice (Argulus japonicus Thiele). Bull Freshw Fish Res Lab 20:109–126
Lewis RM, Hettler WF (1968) Effects of temperature and salinity on the survival of young Atlantic menhaden, Brevoortia tyrannus. Trans Am Fish Soc 97:344–349
Lo CM, Morand S, Galzin R (1998) Parasite diversity/host age and size relationship in three coral–reef fishes from French Polynesia. Int J Parasitol 28:1695–1708
McPherson NJ, Norman RA, Hoyle AS, Bron JE, Taylor NGH (2012) Stocking methods and parasite–induced reductions in capture: modelling Argulus foliaceus in trout fisheries. J Theor Biol 312:22–33
Mikheev VN, Pasternak AF, Valtonen ET (2007) Host specificity of Argulus coregoni (Crustacea: Branchiura) increases at maturation. Parasitology 134(Pt 12):1767–1774
Milinski M (1984) Parasites determine a predator’s optimal feeding strategy. Behav Ecol Sociobiol 15:35–37
Nagasawa K (2009) Synopsis of branchiurans of the genus Argulus (Crustacea, Argulidae), ectoparasites of freshwater and marine fishes, in Japan (1900–2009). Bull Biogeogr Soc Jpn 64:135–148
Omeji S, Solomon SG, Idoga ES (2011) A comparative study of the common protozoan parasites of clarias gariepinus from the wild and cultured environments in Benue State, Nigeria. J Parasitol Res, Article ID 916489, 8 pages. doi:10.1155/2011/916489
Poulin R (2006) Variation in infection parameters among populations within parasite species: intrinsic properties versus local factors. Int J Parasitol 36:877–885
Poulin R, Krasnov BR, Mouillot D (2011) Host specificity in phylogenetic and geographic space. Trends Parasitol 27:355–361
Qizhong Z, Chenglin M (1994) The parasitic crustaceans of fishes from Sichuan Province. J Southeast China Normal Univ (Nat Sci) 19(1):58–61
Tavares–Dias M, Martins ML, Moraes FL (2000) Condition factor, hepatosomatic and splenosomatic relation of freshwater fishes naturally parasitized. Acta Sci Biol Sci 22:533–537
Taylor NGH, Sommerville C, Wootten R (2006) The epidemiology of Argulus spp. (Crustacea: Branchiura) infections in stillwater trout sheries. J Fish Dis 29:193–200
Taylor NGH, Wootten R, Sommerville C (2009) Using length–frequency data to elucidate the population dynamics of Argulus foliaceus (Crustacea: Branchiura). Parasitol 1023–1032
Tucker CS, Sommerville C, Wooten R (2002) Does size really matter? Effects of fish surface area on the settlement and initial survival of Lepeophtheirus salmonis, an ectoparasite of Atlantic salmon Salmo salar. Dis Aquat Org 49:145–152
Violante–Gonzalez J, Garcia–Varela M, Rojas–Herrera A, Guerrero S (2009) Diplostomiasis in cultured and wild tilapia Oreochromis niloticus in Guerrero State, Mexico. Parasitol Res 105:803–807
Wadeh H, Yang JW, Li GQ (2008) Ultrastructure of Argulus japonicus Thiele, 1900 (Crustacea: Branchiura) collected from Guangdong, China. Parasitol Res 102(4):765–770
Walker PD, Flik G, Bonga SEW (2004) The biology of parasites from the genus Argulus and a review of the interactions with its host. Symp Soc Exp Biol 55:107–129
Walker PD, Harris J, Van der Velde G, Bonga SEW (2007) Size matters: stickleback size and infection with Argulus foliaceus (L., 1758) (Branchiura, Arguloida). Crustaceana 80(11):1397–1401
Walker PD, Harris JE, Van der Velde G, Bonga SEW (2008) Differential host utilisation by different life history stages of the fish ectoparasite Argulus foliaceus (Crustacea: Branchiura). Folia Parasitol 55:141–149
Walker PD, Russon IJ, Duijf R, Van der Velde G, Bonga SEW (2011a) The off–host survival and viability of a native and non–native fish louse (Argulus, Crustacea: Branchiura). Curr Zool 57(6):828–835
Walker PD, Russon IJ, Haond C, Van der Velde G, Bonga SEW (2011b) Feeding in adult Argulus japonicus Thiele, 1900 (Maxillopoda, Branchiura), an ectoparasite on fish. Crustaceana 84(3):307–318
Zander CD (2004) Four-year monitoring of parasite communities in gobiid fishes of the south–western Baltic. II: infracommunity. Parasitol Res 93:17–29
Acknowledgments
This work was supported by grants from National Natural Science Foundation of China (grant no. 30972179, 31272551) and the PhD Programs Foundation of Ministry of Education of China (200805640004).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Alsarakibi, M., Wadeh, H. & Li, G. Parasitism of Argulus japonicus in cultured and wild fish of Guangdong, China with new record of three hosts. Parasitol Res 113, 769–775 (2014). https://doi.org/10.1007/s00436-013-3708-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-013-3708-5