
Abstract
The seasonal dynamic of the metazoan parasite community of the blue sea catfish (Sciades guatemalensis) from Tres Palos Lagoon, Guerrero, Mexico, was studied at the component community and infracommunity levels. A total of 382 fish were collected during the regional dry and rainy seasons (a total of seven seasons) between April 2000 and September 2007. Nine helminths were collected: Neotetraonchus sp., Pseudoacanthostomum panamense, Austrodiplostomum compactum, Clinostomum complanatum, Metadena sp., Pseudoleptorhynchoides lamothei, Neoechinorhynchus cf. golvani, Hysterothylacium perezi, and Contracaecum sp. The infection dynamics of some dominant helminths was influenced by environmental changes generated by the dry/rainy season cycle. Nested (non-random) species composition was observed in the infracommunities during almost all of the sample period. Variation in the intensity of nestedness was attributed to a sequential colonization process over time by the dominant helminths.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The blue sea catfish (Sciades guatemalensis) is found along the Pacific coast of Mesoamerica and Central America from the Gulf of California to Panama (Castro-Aguirre et al. 1999). Due to constant local demand, it is a major fishery catch in the coastal lagoons of Guerrero state, Mexico. Considered to be a carnivorous secondary consumer, it is a characteristic estuarine species of the Guerrero coastal lagoon system (Yáñez-Arancibia 1978). The species’ parasite fauna from two coastal lagoons has been reported previously (Violante-González and Aguirre-Macedo 2007; Violante-González et al. 2007), but no studies have focused on the stability of its parasite community over time in terms of species composition and abundance.
The majority of research in Mexico on temporal variation in tropical parasite communities has involved freshwater cichlids (Salgado-Maldonado 1993; Pineda-López 1994; Jiménez 2003; Vidal-Martinez and Poulin 2003), although one study was made of the parasite community of an estuarine eleotrid, Dormitator latifrons, (Violante-González et al. 2008). Some of these studies have shown high variation in parasite abundance over time in response to alterations generated by the seasonal dry/rain cycles, which presumably affect the parasite species recruitment process (Salgado-Maldonado 1993; Violante-González et al. 2008). Abundance has also been shown to respond to seasonal increases in host feeding and reproductive activity influenced by seasonal water temperature fluctuations (Jiménez 2003; Jiménez-García and Vidal-Martínez 2005; Violante-González et al. 2008). Pineda-López (1994), in contrast, suggested that helminth communities in tropical climates are stable or change very little over time. Therefore, it is still unclear, if tropical estuarine parasite communities experience temporal changes in abundance and species composition over time.
For several years we have been collecting parasite community data from Tres Palos Lagoon, Guerrero, during the two seasons that occur regionally (dry and rainy). The objective of this study was to determine if helminth community structure and species composition vary over time in blue sea catfish, S. guatemalensis, from Tres Palos Lagoon.
Materials and methods
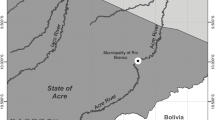
Tres Palos Lagoon (16°47’ N, 99°39’ W; Fig. 1) is located on the Pacific coast of Mexico, 25 km east of Acapulco. It covers 55 km2 (5,500 ha) and ranges in depth from 0.5–8 m. Continuous discharge of urban waste into the lagoon via the Sabana River have made it eutrophic (Violante-González 2006) and consequently primary productivity is very high (80 to 106 μg L−1 chlorophyll-a concentration), particularly during the rainy season (Violante-González et al. 2008). Water in the lagoon is brackish with salinity that ranges from 3 to 5 ppm year round.

Location of Tres Palos Lagoon in the state of Guerrero, Mexico. (Inserts temperature and precipitation recorded during 8 years)
The region experiences two distinct seasons during the year; a rainy season from June to November and a dry season from December to May (Violante-González 2006). For this study, temperature and precipitation data for the area were obtained from the local meteorological station. Differences in environmental parameters between seasons were determined with a one-way analysis of variance (ANOVA) and a Student’s t-test.
A total of 382 adult specimens of S. guatemalensis (total length = 23.4 ± 2.2 cm to 26.4 ± 4.3 cm) were collected using gill nets between April 2000 and September 2007. The number of specimens examined per season each year ranged from 45 to 87 (Table 1). A complete necropsy was made for all fish and helminths were collected from internal and external organs according to Lamothe-Argumedo (1997) and Vidal-Martínez et al. (2001). Voucher specimens of most taxa were deposited in the National Helminth Collection, Institute of Biology, National Autonomous University of Mexico, Mexico City (CNHE).
Autogenic species of parasite were defined as those which reach maturity in aquatic hosts and thus have a limited ability to colonize new locations. Allogenic species were those with birds or mammals as definitive hosts and whose natural migrations favor helminth dispersion, providing them a wide geographic distribution (Esch et al. 1988). Active transmission was defined as movement of a parasite to a host by its own means, whereas passive transmission was defined as movement of a parasite from one host to another without energy expenditure by the parasite (Bush et al. 2003). The host range concept (Rohde 2005) was used to classify the species of parasite as specialists or generalists. According to this concept, a specialist parasite is one that has a marked affinity for a specific host family, genus, or species, while a generalist is one that parasitizes a number of families. Based on their prevalence and abundance, the species of parasite also were classified as dominant (abundant and frequent, prevalence >40% and mean abundance = 6–7.43 parasites per fish), common (low abundance but frequent, prevalence 20–40% and mean abundance = 1–6), or rare (low abundance and low frequency, prevalence <20 % and mean abundance <1 per fish).
Variation in species composition over time was described using prevalence (percent infected hosts) and mean abundance (mean number of parasites per examined fish), calculated and applied according to Bush et al. (1997), for each species of helminth per season. Possible differences in infection parameters between seasons were evaluated using G-tests (Sokal and Rohlf 1998) for prevalence and a χ 2 test for abundance. Significance for all the statistical analyses was established at P = 0.05, unless stated otherwise.
Analyses were made at the component community (all helminth parasites in all hosts collected per season) and infracommunity (all helminth parasites in each individual host) levels (Holmes and Price 1986). Component community parameters included total number of parasite species, total number of individual parasites, the Shannon–Wiener Index (H) as a measure of diversity, species evenness (equitability) (Krebs 1999), and the Berger–Parker Index (BPI) as a measure of numerical dominance (Magurran 1991). A χ 2 test was used to determine differences in component community parameters between seasons and correlations were made using the Spearman range coefficient (r s; Krebs 1999). Infracommunities were described in terms of mean number of helminth species per fish host, mean number of parasite individuals, and mean value of the Brillouin Diversity Index (H′) per fish. A one-way analysis of covariance (ANCOVA) was used to determine differences in infracommunity parameters between seasons.
The presence of nestedness in species composition between seasons was determined with the nestedness temperature calculator (Atmar and Patterson 1995), which employs a randomness matrix based on disorder, or entropy. The relationship between entropy and heat is used to quantify the randomness matrix in terms of temperature, with 0°C representing minimum entropy (perfect nestedness) and 100°C representing maximum entropy (randomness). This is done by building a presence–absence matrix of parasite species in each infracommunity for each sample. The nestedness temperature calculator packs the observed data into the matrix in such a way that presences are concentrated in the upper left-hand corner as much as possible, while primary information (species-per-site data) is maintained within the matrix. The packed matrix is then compared with a maximum nestedness equivalent to identify unexpected presences and absences. For each observed matrix a T° value is calculated where 0°C represents a perfectly nested matrix and 100°C a completely random matrix. The significance of a derived T° was estimated with a t test, using a normal distribution of 500 Monte Carlo simulated T° values. As a complement to this analysis, the infracommunity index (ICI) (Zander 2004), which describes the frequency of double and multiple infections by a single parasite species in a distinct host, was calculated:
where Ij = mean number of parasite species in host j (= mean infracommunity); M ij = the ratio (numbers) of multiple-infected host j with parasite i and other parasites; N j = the ratio (numbers) of infected host j.
Results
Abiotic factors
During the 8-year study period, temperature varied significantly (P < 0.001): 25.2 ± 1.0°C (2000) to 28.8 ± 0.8°C (2007; one-way ANOVA F = 15.11). Precipitation also varied significantly (P < 0.001): 165.10 ± 76.5 mm (2004) to 295.6 ± 255.2 mm (2007; t = 14.4, Fig. 1).
Species composition
Nine species of helminth were identified in 382 specimens of S. guatemalensis collected during seven individual seasons (April 2000 to September 2007): one monogenean, four digeneans, two acanthocephalans, and two nematodes (Table 1). Four of the species recovered as larval stages were classified as allogenic since they mature in ichthyophagous aquatic birds living around the lagoon. The autogenic species were Neotetraonchus sp. (Monogenea), Pseudoacanthostomum panamense (Digenea), Pseudoleptorhynchoides lamothei, and Neoechinorhynchus cf. golvani (Acanthocephala), and Hysterothylacium perezi (Nematoda) (Table 1). All nine were classified as generalists because they have been recorded in other hosts from different families living in the same lagoon (Violante-González et al. 2007).
The frequency distribution of prevalence values for each species in all seasons produced three separate groups: dominant species (prevalence >40%), H. perezi and P. lamothei; common species (20–40%), Neotetraonchus sp., P. panamense, and Austrodiplostomum compactum; and rare species (prevalence <20%). Considering all sampling periods, prevalence and abundance differed significantly between seasons and years in four species (Table 1). Relatively high percentages of infection were observed for H. perezi in the 2002 dry season and the 2002 and 2007 rainy seasons (G = 46.11, P < 0.05); for P. lamothei in the 2002 and 2003 dry seasons (G = 73.63, P < 0.05); and for P. panamense and Neotetraonchus sp. in the 2007 rainy season (G = 87.75, P < 0.05 and G = 108.04, P < 0.05, respectively). The highest mean abundance values for these same species were recorded in these same sampling periods (χ 2 test, P < 0.05; Table 1), indicating a positive correlation between prevalence and average abundance values in each case (P < 0.05).
Component community
Species collected by season varied from seven to nine, although the number did not vary significantly between any seasons (χ 2 = 0.44, P > 0.05; Table 2). Total number of individual parasites ranged from 84 (2000 dry) to 2,885 (2007 rainy), and varied significantly between seasons and years (χ 2 = 520.50, P < 0.05; Table 2). P. lamothei was numerically dominant during six consecutive seasons, while H. perezi was numerically dominant only during the 2007 rainy season (Table 2). Shannon–Wiener diversity index values ranged from 1.65 to 2.43, although these did not differ between seasons and years (χ 2 = 0.26, P > 0.05). Evenness was positively correlated with diversity values during all the seasons sampled (r s = 0.89, P < 0.01).
Infracommunities
All fish collected were adults, though body size differed significantly between sampling periods (one-way ANOVA; F = 4.47, P < 0.05). The largest fish (26.6 ± 4.2) were collected in the 2000 rainy season, and the smallest (23.4 ± 2.2) in the 2003 dry season. Length of fish was positively correlated to mean number of individual parasites (r s = 0.300, P < 0.05, n = 307).
The mean number of species of helminth ranged from 1.52 ± 0.63 (dry-2001) to 2.89 ± 1.1 (rainy-2007), and the mean number of individual parasites from 4.0 ± 2.7 (dry-2000) to 45.08 ± 97.51 (rainy-2007; Table 2). The Brillouin diversity index values varied from 0.25 ± 0.35 (rainy-2000) to 0.81 ± 0.45 (rainy-2007). The mean number of species of helminth was highest in the 2002 dry season and 2007 rainy season (one-way ANCOVA F = 28.41, P < 0.001), as was mean number of individuals (one-way ANCOVA F = 4.25, P < 0.001) and mean diversity (one-way ANCOVA F = 5.96, P < 0.001).
Nestedness
Nestedness occurred during six of the sampling periods, the exception being the 2000 dry season (Table 3). Nestedness intensity (lowest value T°) was highest in the 2007 rainy season (T° = 11.45°). Species composition remained almost unchanged, although the rank assigned to each species within the packed component community matrix varied between seasons and years (Table 3). H. perezi and P. lamothei were top-ranked in the highest number of seasons (three each) (Table 3).
The infracommunity index values (ICI) indicated that H. perezi, P. lamothei, and Neotetraonchus sp. had the highest number of double or multiple co-occurrences with other species of helminth (ICI > 0.20) in two or more seasons. Nestedness intensity depended on the higher number of double or multiple co-occurrences between these three species and the remaining six species in the S. guatemalensis parasite community.
Discussion
The results indicate that although species composition and species richness of helminths varied little during the seven-season study period, some species exhibited seasonal changes in their infection dynamic. Two biotic factors may have influenced these seasonal variations. Environmental variations caused by the seasonal dry/rainy cycle influence intermediate host availability and can lead to consequent alterations in the recruitment processes of some allogenic species. Changes in the feeding and reproductive behavior of S. guatemalensis may also influence the recruitment of intestinal species.
Five members of the parasite community of S. guatemalensis were autogenic species: Neotetraonchus sp., P. panamense, P. lamothei, N. cf. golvani, and H. perezi. The remaining four species were classified as allogenic because they mature in the aquatic birds, Casmerodius albus egretta (great egret) and Phalacrocorax olivaceus (Neotropic cormorant), which inhabit Tres Palos Lagoon (Violante-González 2006; Table 1). All five of the autogenic species are considered to be generalists because they have been reported in other hosts from the same lagoon (Violante-González 2006; Violante-González et al. 2007), although at least P. panamense, P. lamothei, and H. perezi mature only in S. guatemalensis, indicating that this species is their preferential host (Holmes 1976).
The seasonal variations observed in the infection parameters of the four most frequent and abundant species of helminth suggest that the transmission process of some parasites can experience interseasonal or interannual fluctuations that may be due to variations in intermediate host availability (Moravec et al. 2002; Violante-González 2006; Violante-González et al. 2008). During the dry season (December–May), the environment of Tres Palos Lagoon is relatively stable (Violante-González 2006). This could favor transmission of some parasites (both active and passive transmission forms) through higher availability of intermediate hosts (snails or crustaceans). The feeding activity of S. guatemalensis is also higher during the dry season than in the rainy season, when it reproduces (Violante-González 2006). This favors acquisition of parasites during the dry seasons by passive transmission. Two species, P. lamothei and H. perezi, that use passive transmission had higher infection levels during some dry seasons (Table 1). In previous studies, temporal variation in the infection levels of the acanthocephalans Neoechinorhynchus agilis and N. cf. golvani were found to be linked to reproductive aspects of its hosts Liza abu (Mhaisen et al. 1988) and D. latifrons (Violante-González et al. 2008). Indeed, many studies have shown that seasonal variation in the diet and amount of food ingested by hosts is clearly related to the population dynamics and community structure of parasites (Esch et al. 1988; Zander and Kesting 1998; Fiorillo and Font 1999; Zander et al. 2002; Klimpel et al. 2003; Fellis and Esch 2004; Zander 2003, 2004, 2005).
The stable environmental conditions in the lagoon during the dry season become unstable during the rainy season (July to November) in response to high input of organic matter from the contiguous Sabana River and other tributaries. This influx causes high mortality among benthonic invertebrates, such as snails, which are buried by silt (Violante-González 2006). However, rainfall can vary widely from 1 year to the next (Fig. 1), producing an inherent annual instability in addition to the interannual variations, possibly explaining the lack of any clear pattern in seasonal variation in the community structure of the parasites of this fish.
At both the component and infracommunity levels, low species abundance and diversity of helminths were found in S. guatemalensis (Table 2). Richness (nine species of helminth) was lower than the 14 species reported for the catfish Ariopsis asssimilis (Vidal-Martínez et al. 2003), but similar to that reported in Cathorops seemani (nine species; Pérez-Ponce et al. 1999).
The low complexity food web structure in Tres Palos Lagoon may explain the low species richness in the parasite community of S. guatemalensis since food web structure is known to affect species richness and diversity of parasites in many freshwater systems (Carney and Dick 2000; Marcogliese 2001). The community of zooplankton is not very diverse, and the macrobenthoic community, including molluscs, is represented by only six species (Stuardo and Villarroel 1976). This low level of biodiversity clearly restricts intermediate host availability in the food web, reducing opportunities for parasite transmission (Carney and Dick 2000; Marcogliese 2001).
At the component community level, both community composition and number of helminth species were similar across the sampling periods. Parasite load (i.e., total number of parasite individuals), however, was significantly higher in the 2002 dry season and 2007 rainy season (Table 2). This seasonal variation is probably related to the higher abundance of P. lamothei and H. perezi in each of these seasons.
The main factors structuring the parasite infracommunities (Holmes 1990; Zander 2004) in S. guatemalensis were the same ones structuring the component communities; that is, feeding behavior and seasonal variation in the availability of infective stages (cercariae or oncomiracidia), both influenced in turn by the seasonal dry/rainy cycle. These two biotic factors structure infracommunities primarily through accumulation of temporarily free living cercariae (P. panamense, A. compactum, and C. complanatum) or ingestion of infected crustaceans (P. lamothei and H. perezi). Acquisition of some rare species completes the structure.
The presence of nestedness in almost all the seasons that were studied suggests a certain degree of structure in infracommunity species composition over time (Poulin and Valtonen 2001), in other words, species composition exhibited a certain degree of order. Some species of helminth were prevalent at high levels in all infracommunity classes (rich and poor), whereas rare species occurred mainly in species-rich infracommunities.
Nestedness within infracommunities can be generated by several processes, including passive sampling and sequential colonization of parasite species (Carney and Dick 2000; Timi and Poulin 2003). The passive sampling hypothesis is based on probability and predicts that common species will occur in many hosts and rare species in fewer hosts, therefore producing a nested pattern. The nestedness observed in the current study may have been caused by stability in species composition over time. However, some of the dominant species occurred in a higher number of double or multiple co-occurrences with other species (ICI > 0.20, Table 3) and were more abundant in different seasons, suggesting that a sequential colonization process may better explain the variation in intensity of nestedness observed here.
Thus, nestedness can be considered a feature of the parasite infracommunity composition in S. guatemalensis in Tres Palos Lagoon. Nestedness has been reported in the parasite communities of fish from other locations (Carney and Dick 2000; Poulin and Valtonen 2001; Valtonen et al. 2001; Timi and Poulin 2003; Vidal-Martinez and Poulin 2003; Norton et al. 2004), including one eleotrid from this same lagoon (Violante-González et al. 2008).
Overall, the results indicate that even though species composition and richness of helminths remained relatively constant over time, the infection dynamics of some dominant helminths were influenced by regional environmental conditions. Environmental factors (dry/rainy cycles) influence intermediate host availability in Tres Palos lagoon and generate alterations in the recruitment processes of some autogenic parasite species. Environmental variations also cause changes in the feeding and reproductive behavior of S. guatemalensis, which in turn influences recruitment of intestinal helminth species.
References
Atmar W, Patterson BD (1995) The nestedness temperature calculator: a visual basic program, including 294 presence–absence matrices. AICS Research, Inc., University Park New Mexico, and The Field Museum, Chicago
Bush AO, Lafferty KD, Lotz JM, Shostak AW (1997) Parasitology meets ecology on its own terms: Margolis et al. revisited. J Parasitol 83:575–583
Bush AO, Fernandez J, Esch GW, Seed JR (2003) Parasitism: the diversity and ecology of animal parasites. Cambridge University Press, Cambridge, p 566
Carney JP, Dick TA (2000) Helminth communities of yellow perch Perca flavescens (Mitchill): determinants of pattern. Can J Zool 78:538–555
Castro-Aguirre JL, Espinosa-Pérez HS, Schmitter-Soto JJ (1999) Ictiofauna estuarino-lagunar y vicaria de México. Limusa, México, p 711
Esch GW, Kennedy CR, Bush AO, Aho JM (1988) Patterns in helminth communities in freshwater fish in Great Britain: alternative strategies for colonization. Parasitol 96:519–532
Fellis KJ, Esch GW (2004) Community structure and seasonal dynamics of helminth parasites in Lepomis cyanellus and L. macrochirus from Charlie’s Pond, North Carolina: host size and species as determinants of community structure. J Parasitol 90:41–49
Fiorillo RA, Font WF (1999) Seasonal dynamics and community structure of helminths of spotted sunfish, Lepomis miniatus (Osteichthyes: Centrarchidae) from an oligohaline estuary in southeastern Louisiana, USA. J Helminthol Soc Wash 66:101–110
Holmes JC (1976) Host selection and its consequences. In: Kennedy CR (ed) Ecological aspects of parasitology. North-Holland, Amsterdam, pp 21–39
Holmes JC (1990) Helminth communities of marine fishes. In: Esch GW, Bush AO, Aho JM (eds) Parasite communities: patterns and process. Chapman and Hall, London, pp 101–130
Holmes JC, Price PW (1986) Communities of parasites. In: Anderson DJ, Kikkawa J (eds) Community ecology: patterns and processes. Blackwell Scientific, Oxford, pp 187–213
Jiménez MI (2003) Estructura temporal de las comunidades de parásitos metazoarios en poblaciones de cíclidos silvestres (Cichlasoma urophthalmus) e introducidos en Mitza, Yucatán: Un enfoque experimental. Tesis Doctoral Cinvestav-IPN, Mérida, p 174
Jiménez-García MI, Vidal-Martínez VM (2005) Temporal variation in the infection dynamics and maturation cycle of Oligogonotylus manteri (Digenea) in the cichlid fish, Cichlasoma urophtalmus, from Yucatán, México. J Parasitol 91:1008–1014
Klimpel S, Seehagen A, Palm HW (2003) Metazoan parasites and feeding behavior of four small-sized fish species from the central North Sea. Parasitol Res 91:290–297
Krebs CJ (1999) Ecological methodology. Addison-Welsey Educational, Boston, p 654
Lamothe-Argumedo R (1997) Manual de técnicas para preparar y estudiar los parásitos de animales silvestres. México City, D F, p 43
Magurran A (1991) Ecological diversity and its measurement. Princeton University Press, Princeton, p 178
Marcogliese DJ (2001) Pursuing parasites up the food chain: implications of food web structure and function on parasite communities in aquatic systems. Acta Parasitol 46:82–93
Mhaisen FT, Al-Salim NK, Khamees R (1988) Occurrence of parasites of the freshwater mugilid fish Lisa abu (Heckel) from Basrah, Southern Iraq. J Fish Biol 32:525–532
Moravec F, Mendoza-Franco E, Vivas-Rodríguez C, Vargas-Vázquez J, González-Solís D (2002) Observations on seasonal changes in the occurrence and maturation of five helminth species in the pimelodid catfish, Rhamdia guatemalensis, in the cenote (= sinkhole) Ixin-há, Yucatan, Mexico. Acta Societ Zool Bohemosl 66:121–140
Norton J, Lewis JW, Rollison D (2004) Temporal and spatial patterns of nestedness in eel macroparasite communities. Parasitol 129:203–211
Pérez-Ponce de León G, García-Prieto L, Mendoza-Garfias B, Léon-Règagnon V, Pulido-Flores G, Aranda-Cruz C, García-Vargas F (1999) IX. Biodiversidad de helmintos parásitos de peces marinos y estuarinos de la Bahía de Chamela, Jalisco. Listados faunísticos de México Universidad Nacional Autónoma de México, p 51
Pineda-López R (1994) Ecology of the helminth communities of cichlid fish in the flood plains of Southeastern Mexico. Ph. D. Thesis. Univ Ex, Exeter U K, p 236
Poulin R, Valtonen ET (2001) Nested assemblages resulting from host size variation: the case of endoparasite communities in fish hosts. Int J Parasitol 31:1194–1204
Rohde K (2005) Ecology of marine parasites. CSIRO, Wallingford, p 565
Salgado-Maldonado G (1993) Ecología de helmintos parásitos de Cichlasoma urophthalmus (Pisces: Cichlidae) en la Península de Yucatán, México. Tesis Doctoral. Cinvestav-IPN, Mérida, p 357
Sokal RR, Rohlf FJ (1998) Biometry, 2nd edn. W. H. Freeman, San Francisco, p 859
Stuardo J, Villarroel M (1976) Aspectos ecológicos y distribución de los moluscos en las lagunas costeras de Guerrero, México. Anales Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México. México City, DF. 1:70–90
Timi TT, Poulin R (2003) Parasite community structure within and across host populations of a marine pelagic fish: how repeatable is it? Int J Parasitol 33:1353–1362
Valtonen ET, Pulkinen K, Poulin R, Julkunen M (2001) The structure of parasite component communities in brackish water fishes of the northeastern Baltic Sea. Parasitol 122:471–481
Vidal-Martinez VM, Poulin R (2003) Spatial and temporal repeatability in parasite community structure of tropical fish hosts. Parasitol 127:387–398
Vidal-Martínez VM, Aguirre-Macedo L, Scholz T, González-Solís D, Mendoza-Franco EF (2001) Atlas of helminth parasites of Cichlid fish of Mexico. Academy of Sciences of the Czech Republic, p 165
Vidal-Martínez VM, Aguirre-Macedo ML, Noreña-Barroso E, Gold-Bouchot G, Caballero-Pinzón PI (2003) Potential interactions between metazoan parasites of the Mayan catfish Ariopsis assimilis and the chemical pollution in Chetumal Bay, Mexico. J Helmintol 77:173–184
Violante-González J (2006) Comunidades de parásitos metazoarios de peces, en dos lagunas costeras del Estado de Guerrero, México. Tesis Doctoral. Cinvestav-IPN, Mérida, p 153
Violante-González J, Aguirre-Macedo ML (2007) Metazoan parasites of fishes from Coyuca Lagoon, Guerrero, Mexico. Zoot 1531:39–48
Violante-González J, Aguirre-Macedo ML, Mendoza-Franco EF (2007) A checklist of metazoan parasites of fish from Tres Palos lagoon, Guerrero, Mexico. Parasitol Res 102:151–161
Violante-González J, Aguirre-Macedo ML, Vidal-Martínez VM (2008) The temporal variation in the helminth parasite communities of pacific fat sleeper Dormitator latifrons from Tres Palos lagoon, Guerrero, Mexico. J Parasitol 94:326–334
Yáñez-Arancibia A (1978) Taxonomía, ecología y estructura de las comunidades de peces en lagunas costeras con bocas efímeras del Pacífico de México. Centro de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México Publicaciones Especiales 2:1–306
Zander CD (2003) Four-year monitoring of parasite communities in gobiid fishes of the south-western Baltic. I. Guild and component community. Parasitol Res 90:502–511
Zander CD (2004) Four-year monitoring of parasite communities in gobiid fishes of the south-western Baltic. II. Infracommunity. Parasitol Res 93:17–29
Zander CD (2005) Four-year monitoring of parasite communities in gobiid fishes of the southwest Baltic. III. Parasite species diversity and applicability of monitoring. Parasitol Res 95:136–144
Zander CD, Kesting V (1998) Colonization and seasonality of goby (Gobiidae, Teleostei) parasites from the southwestern Baltic Sea. Parasitol Res 84:459–466
Zander CD, Kocoglu Ö, Skroblies M (2002) Parasite populations and communities from the shallow littoral of the Orther Bight (Fehmarn, SW Baltic Sea). Parasitol Res 88:734–744
Acknowledgements
The authors thank the PROMEP: Programa de Reincorporación de Exbecarios for partially funding this research and the students of the Marine Ecology Academic Unit (UAG) for their assistance in the field and laboratory work. We thank two anonymous reviewers whose extensive and thoughtful comments substantially improved the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Violante-González, J., Aguirre-Macedo, M.L., Rojas-Herrera, A. et al. Metazoan parasite community of blue sea catfish, Sciades guatemalensis (Ariidae), from Tres Palos Lagoon, Guerrero, Mexico. Parasitol Res 105, 997–1005 (2009). https://doi.org/10.1007/s00436-009-1488-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-009-1488-8