
Abstract
This study tested the effects of “living high-training low” (Hi–Lo) on aerobic performance and economy of work in elite athletes. Forty endurance athletes (cross-country skiers, swimmers, runners) performed 13–18 consecutive days of training at 1,200 m altitude, by sleeping at 1,200 m (LL, n = 20) or in hypoxic rooms with 5–6 nights at 2,500 m followed by 8–12 nights at 3,000–3,500 m (HL, n = 20). The athletes were evaluated before (pre-), one (post-1) and 15 days (post-15) after Hi–Lo. Economy was assessed from two sub-maximal tests, one non-specific (cycling) and one specific (running or swimming). From pre- to post-1: \(\dot{V}\hbox{O}_{2{\rm max}}\) increased both in HL (+ 7.8%, P < 0.01) and in LL (+ 3.3%, P < 0.05), peak power output (PPO) tended to increase more (P=0.06) in HL (+ 4.1%, P < 0.01) than in LL (+ 1.9%). At post-15, \(\dot{V}\hbox{O}_{2{\rm max}}\) has returned to pre-values in both groups, PPO increased more (P < 0.05) in HL (+ 8.3%, P < 0.01) than in LL (+ 3.8%), \(\dot{V}\hbox{O}_{2}\) and power at respiratory compensation point (RCP) increased more (P < 0.05) in HL (+ 9.5%, P < 0.01 and + 11.2%, P < 0.01) than in LL (+ 3.2 and + 3.3%). Cycling mechanical efficiency (8–5%) and economy during specific locomotion (7–7%) increased (P < 0.05) in both groups. This study shows that, for a similar increase in \(\dot{V}\hbox{O}_{2{\rm max}},\) HL had a greater increase in PPO than LL. The efficiency of Hi–Lo is also evidenced 15 days later by higher \(\dot{V}\hbox{O}_{2}\) and power at RCP. This study emphasizes that during the post-altitude period, economy of work greatly increases in both groups.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Altitude training has been used for several decades by athletes. In the 1990s, a new model “living high–training low” (Hi–Lo) was proposed as a more efficient way to enhance sea-level performance in trained endurance athletes (Levine and Stray-Gundersen 1997). The underlying mechanisms were an increase in erythropoiesis, red cell mass and blood oxygen carrying capacity, leading to an increase in \(\dot{V}\hbox{O}_{2{\rm max}}\) in parallel to a preservation of the training intensity, O2 flux and muscle function while training at or near sea level (Levine and Stray-Gundersen 1997; Melissa et al. 1997). Other studies rather support a “peripheral” hypothesis (local changes in skeletal muscle) for explaining the improvement in performance detected with Hi–Lo, through an increase in muscle buffer capacity (Gore et al. 2001) or economy of exercise (Saunders et al. 2004). The exact underlying mechanisms have been debated with some studies demonstrating no improvement in performance, maximal oxygen transport or erythropoietic responses (Ashenden et al. 1999). It has even been reported that Hi–Lo decreases maximal O2 transport (Gore et al. 2001). And to date, the debate on the efficiency of Hi–Lo is not closed as shown it the recent Point-Counterpoint on the Hi–Lo strategy (Levine and Stray-Gundersen 2005; Gore and Hopkins 2005). Among the potential mechanisms underlying the enhancement in endurance performance after Hi–Lo, some points have received little attention. The change in physiological variables at submaximal velocities such as at the onset of blood lactate accumulation (OBLA) or at respiratory compensation point (RCP) after Hi–Lo are poorly documented, although these factors are key determinants of endurance performance (Meyer et al. 2004; Tanaka 1990). Levine and Stray-Gundersen (1997) first showed an improvement in velocity at the ventilatory threshold after four Hi–Lo weeks. Economy of exercise was shown to be related to the level of performance in the main endurance sports such as swimming (Toussaint and Hollander 1994), running (Saunders et al. 2004) or cross-country skiing (Millet et al. 2003). To date, the effects of altitude training on the sea-level economy are still debated: Levine and Stray-Gundersen (1997) and Telford et al. (1996) reported no change in submaximal \(\dot{V}\hbox{O}_{2}\) just after a period of altitude training. Conversely, it was reported that running economy (Saunders et al. 2004), or cycling efficiency (Gore et al. 2001) were significantly improved after Hi–Lo, when compared to a matched control group. Green et al. (2000) suggested that after altitude acclimatization the muscular efficiency was improved either by the increase in ATP per mole O2 or by the decrease in ATP necessary at a given intensity. It is also unknown if the post-altitude changes in economy are transferable to other means of locomotion, as expected if due to central cardiovascular adaptations. One major question regarding Hi–Lo is the duration of the potential effects on performance. Some studies showed positive effects on \(\dot{V}\hbox{O}_{2{\rm max}}\) and performance at the end of Hi–Lo (Levine and Stray-Gundersen 1992, 1997). However, it is unknown if the optimal time period after Hi–Lo is similar to those traditionally defined by coaches with empirical observation after the living high-training high method (e.g. Dick 1992), i.e. 12–20 days.
Therefore, the primary objective of this study was to determine the effects of Hi–Lo on endurance performance and economy of work and its underlying factors in various sports. The second objective was to evaluate the persistence of these effects 15 days after Hi–Lo.
Methods
This study brought together three experiments based on the Hi–Lo model, described by Levine and Stray-Gundersen (1992, 1997). The first experiment involved cross-country skiers and took place during the summer of 2002. The second experiment with swimmers was realized during the winter of 2002–2003. The third experiment with runners was performed during the autumn of 2003. Modifications in procedures took into account the results of the preceding study. This work was part of a research project funded by the International Olympic Committee and the French Ministry of Sports. The Ethics Committee of Paris Necker Hospital approved these studies.
Subjects
All subjects were classified as elite by their participation to national and international competitions and were registered on a national elite list of the French ministry of sports. All subjects gave their voluntary written informed consent. The selected characteristics of the subjects are shown in Table 1. In each experiment, subjects were divided into two groups, a “living high–training low” group (HL) and a “living low–training low” group (LL). After the determination of maximal oxygen uptake \(\left(\dot{V}\hbox{O}_{2{\rm max}} \right)\) at the altitude of 1,200 m, the subjects were ranked according to their \(\dot{V}\hbox{O}_{2{\rm max}},\) and assigned to two equivalent \(\dot{V}\hbox{O}_{2{\rm max}}\) groups.
Experiment 1: Cross-country skiers
Eleven international-level athletes (seven biathletes, two nordic-combined skiers and two cross-country skiers) were divided in the HL (n = 6) or in the LL (n = 5) groups. There were three females in each group.
Experiment 2: Swimmers
Eighteen national-level swimmers were divided in the HL (n = 9) or in the LL (n = 9) groups. There was one female athlete in each group.
Experiment 3: Runners
Eleven male runners were divided in the HL (n = 5) or to the LL (n = 6) groups.
Experimental design
Each experiment was performed at the same location (Prémanon, France), at the ‘French national training center for Nordic ski’ at the altitude of 1,200 m and consisted in the following phases: pre-training tests (pre-), Hi–Lo or “living low–training low”(Lo–Lo) training period and post-training tests after one day (post-1) and 15 days (post-15) following the training period. Each experiment was conducted during the base training preparation of the competitive season of the subjects at least 2 months before the beginning of their respective national or international season. All tests were conducted at the altitude of 1,200 m.
Training period
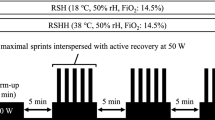
It was based on the “Hi–Lo” model for the HL groups with an introductory period of 5–6 days at 2,500 m followed by 12–18 consecutive nights at altitude between 3,000 and 3,500 m. The LL group slept at 1,200 m for the whole period. Details of the three experimental protocols are shown in Table 2.
Post-training period
After the study training period and the post-1 tests, all subjects returned to sea-level for 2 weeks and maintained their usual training activities. Then, they came back for 2 days at 1,200 m for performing the post-15 tests.
Training
Since the subjects were elite and this experiment was supported by the Ministry of Sports, the training program was carefully designed for matching the best standards in their sports, in collaboration with the respective national coaches. The experiments took place during the preparatory training period after three mesocycles of four aerobic training weeks and a minimum of 2 months prior the competitive period, as required by the International Olympic Commitee (IOC).
Training quantification
The quantification of the training stimulus, in term of volume (min) and intensity (%HRmax) was performed in all experiments. The training load was calculated according to the method of Banister et al. (1975) revised by Busso et al. (1997) and expressed in training impulses (TRIMPS). The athletes being in the same team (national team for nordic-skiers, elite club for swimmers and military national team for runners), they carried out the same training in the weeks preceding the Hi–Lo training camp. The “groups” effects were thus controlled.
Hypoxic rooms
Each HL or LL subject slept in the same type of double room. In the hypoxic rooms, the normobaric hypoxia was obtained by extracting oxygen from ambient air (OBS, Husøysund, Norway). Altitudes of 2,500, 3,000 and 3,500 m were obtained with an O2 fraction (F IO2) of 0.174, 0.164 and 0.154, respectively. For safety reasons and reliability of measurement of the hypoxic level, fractions of O2, CO2, humidity rate and barometric pressure were continuously monitored and controlled. A visual and acoustic alarm was set at levels of 14% O2 and 0.08% CO2. The hypoxic rooms were connected to a central computer located in an adjacent room and were under the permanent control of a medical doctor.
Measurements
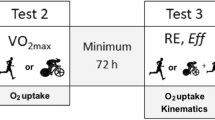
In each experiment, several testing trials were performed at pre- and repeated at post-1 and post-15 (Table 2): the first day, an incremental test to exhaustion to determine \(\dot{V}\hbox{O}_{2{\rm max}},\) peak power output (PPO), \(\dot{V}\hbox{O}_{2}\) and power output at the respiratory compensation point \(\left(\dot{V}\hbox{O}_{2{\rm RCP}}\; \hbox{and}\; {P}_{\rm RCP} \right)\) and the aerobic energy cost in specific locomotion (ECspe); the second day, a submaximal constant cycling test to determine the mechanical efficiency in non-specific locomotion (MEbic).
Incremental test to exhaustion
The skiers and runners performed an incremental test on treadmill (Chrono Run Electronic 500 ES®, Air Machine, Cesena, Italy) with the following protocol, which is routinely used for the medical follow-up of the French cross-country skiers: after a warm-up at 6 km h−1 at 6%-grade for 2 min, subjects ran at 7 km h−1 at 6%-grade for 2 min, then at 8 km h−1 at 6%-grade for another 2 min. Thereafter, the speed was kept constant at 8 km h−1 and the slope was increased by 2%-grade every 2 min up to 14%. Then, the slope was kept constant at 14%-grade and the speed was increased by 1 km h−1 every 2 min until exhaustion. The use of a 14%-slope during maximal treadmill running was in agreement with previous protocols involving elite cross-country skiers, in which slopes ranged from 10 to 21% (Ingjer and Myhre 1992). Cardio-respiratory measurements were performed in breath-by-breath (M VMAX series 29C®, SensorMedics Corporation, Yorba Linda, CA, USA). \(\dot{V}\hbox{O}_{2{\rm max}}\) corresponded to the peak in \(\dot{V}\hbox{O}_{2}\) averaged over 30 s. Peak power output (PPO) was determined from the speed and slope corresponding to the attainment of \(\dot{V}\hbox{O}_{2{\rm max}}\) and calculated starting from the last min average values of the \(\dot{V}\hbox{O}_{2{\rm max}}\) stage. PPO was expressed in watts.
where
- M :
-
the mass of the subject in kg
- g :
-
the acceleration of gravity (9.81 m s−2)
- s :
-
the treadmill speed in m s−1
- α:
-
the angle of the treadmill with the horizontal.
\(\dot{V}\hbox{O}_{2{\rm RCP}}\) and P RCP were determined at the respiratory compensation point (RCP) which corresponds to the onset of hyperventilation during incremental exercise (Morgan et al. 1989). The RCP was established when \(\dot{V}\hbox{E}/\dot{V}\hbox{CO}_{2}\) increased with the first decrease in PETCO2. It followed the steady-state phase after the ventilatory threshold (VT) corresponding to the point where \(\dot{V}\hbox{E}/ \dot{V}\hbox{O}_{2}\) and \(\hbox{PET}_{{\rm O}_2}\) began to increase without a simultaneous increase in \(\hbox{PET}_{{\rm CO}_2}\) All measurements of RCP were made by visual inspection of graphs of power output plotted against each relevant respiratory variable. The visual inspection was made for the three experiments by two experienced exercise physiologists. The results were then averaged.
The swimmers performed an incremental swimming test to exhaustion with gas exchange analysis (K4b2, Cosmed Rome, Italy) in an indoor 25 m swimming pool with temperature at 26°C. The K4b2 was connected to a snorkel (Aquatrainer), validated by Keskinen et al. (2003). The test was preceded by a standardized 400 m warm-up. The test comprised 5 × 200 m repetitions of increasing velocity with 15 s rest interval. Velocity of each swim was determined from each swimmer’s personal best competition time (PCT) in that distance measured in the preceding month. The first 200 m effort was set at 30 s < PCT, with the time to complete the remaining efforts decreasing by 5 s with each stage. The final 200 m were performed at maximal velocity. Swimmer’s velocity was controlled with the subject adjusting the velocity to auditory signals at 12.5 m intervals, delimited by visual marks along the bottom of the pool. Exhaustion coincided with: (a) heart rate (HR) approaching the maximal theoretical HR (220, age), (b) \(\dot{V}\hbox{O}_{2}\) leveling off even with an increase in intensity. Maximal speed (V max, m s−1) was defined as the speed obtained during the last 200 m. HR was continuously recorded with a heart rate monitor (S810®, Polar, Kempele, Finland).
Specific aerobic energy cost
The net energy cost (EC) was calculated in swimming and running as proposed by di Prampero (1986). In each trial the resting \(\dot{V}\hbox{O}_{2}\) value was the 1-min average values measured in a standing position prior the beginning of the test.
Cross-country skiers and runners
For each subject, EC was calculated from the treadmill incremental test at a constant 8%-grade. The \(\dot{V}\hbox{O}_{2}\) value was the 1-min average values measured during the last minute of the stage preceding the VT stage. Thus, the intensity of exercise was lower than VT for all the subjects and allowed a calculation of EC in an exercise intensity expected to be mainly aerobic. EC was expressed in ml kg−1 km−1
Swimmers
EC was calculated at the first stage of the swimming incremental test from the ratio of \(\dot{V}\hbox{O}_{2}\) above resting (ml min−1 kg−1) divided by the speed (m s−1). Reported to body weight, EC was expressed in ml m−1.
EC values were expressed as absolute values and percentage of pre-values in the three experiments.
Steady state cycling submaximal test
In the three experiments, a second test consisting to a steady state trial of 4 min was performed on a cycle ergometer (Chrono Bike Electronic 400 ES®, Air Machine, Cesena, Italy). The cycling mechanical efficiency (%) (MEbic) was calculated as the work performed divided by the energy expended (Moseley and Jeukendrup 2001). MEbic was calculated at a constant power corresponding to a heart rate of 130 beats min−1 steady state measured during the pre Hi–Lo tests. This steady state is an essential condition for a reliable measurement. The \(\dot{V}\hbox{O}_{2}\) values (ml min−1) were recorded breath-by-breath and averaged during the last min of the trial.
Energy expended = net \(\dot{V}\hbox{O}_{2}\)(ml min−1 kg−1) EO2(kJ l−1) 60−1, with resting \(\dot{V}\hbox{O}_{2}\) removed.
Resting \(\dot{V}\hbox{O}_{2}\) was the 1 min average values measured in a sitting position prior the beginning of the test.
EO2 was the energy equivalent and expressed in kJ l−1
RER is the respiratory exchange ratio. \( \hbox{RER} = \dot{V}\hbox{CO}_{2} /\dot{V}\hbox{O}_{2}.\)
Hematological variables
The analysis of the hematological factors was reported in two previous studies, Robach et al. (2006) for the swimmers, and Brugniaux et al. (2006) for the runners. In the current study we have reported only the analysis of the hemoglobin level. Blood samples were obtained at rest by venopuncture by a physician on three occasions: at pre- and repeated at post-1 and post-15. These samples were analyzed, following standard procedures, in the two hours following the blood samplers, by the Pentra 120 analyzer (ABX Montpellier, France), for hemoglobin (Hb; g dl−1).
Statistical analysis
All data are expressed as mean ± SD. The Student t-test was used to analyze the differences between the tests. To compare the characteristics of the two groups, the U Mann-Whitney test was used. The influence of the two training methods on the measured variables was, after analysis of the normality and the homogeneity of variance of the tested samples, analyzed using a two-way analysis of variance (ANOVA) [group (HL, LL) × time [pre-, post-1, post-15]] with repeated measures on the second factor; for example, two \((\dot{V}\hbox{O}_{\rm 2RCP}, {P}_{\rm RCP}, \hbox{ME}_{\rm bic})\) or three \((\dot{V}\hbox{O}_{\rm 2max}, \hbox{PPO}, \hbox{EC}_{\rm spe}, \hbox{Hb})\) measures. Significant effects were subsequently analyzed using the Scheffe’s post hoc test. Since post-1 tests were not conducted in the Nordic skiers, the conditions of application of the 2-way ANOVA were re-examined separately. The number of subjects selected was 13 (HL) and 13 (LL) in the comparison between pre- and post-1, and of 19 (HL) and 19 (LL) in the comparison between pre- and post-15.
The Statview version 5.0 statistical package was used for these analyses. For all statistical analyses, a P value of 0.05 was accepted as the level of statistical significance.
Results
Training
In each experiment, the training loads were usual for the athletes at this moment of the preparation period, and training loads and intensities were always similar in HL and LL (Table 3). During the training period, training loads remained constant in swimmers and runners but decreased in skiers. The training was mainly aerobic with 70% inferior or equal to VT and 93% inferior to RCP during the Hi–Lo period (Table 4). The post-training period presented a decrease in training loads in the two groups (HL and LL).
Performance
The variables measured during the tests are shown in Table 5.
\(\dot{V}\hbox{O}_{\rm 2max}\) and PPO
\(\dot{V}\hbox{O}_{\rm 2max}\) increased at post-1 in HL (P < 0.01) and in LL (P < 0.05) to the same extent (P = 0.11, F = 2.8) and returned to pre-values at post-15. PPO increased at post-1 in HL (P < 0.01), not in LL, with a tendency to a significant difference of change (P = 0.09, F = 2.2) in HL than in LL. At post-15, PPO increased in HL (P < 0.001), not in LL, with a significant difference of change (P = 0.03, F = 4.6) in HL than in LL.
\(\dot{V}\hbox{O}_{2}\) and power at respiratory compensation point
Living high–training low had a significantly greater improvement in \(\dot{V}\hbox{O}_{\rm 2RCP}\) (P = 0.05, F = 4.1) and P RCP (P = 0.01, F = 6.4) between pre- and post-15, when compared to LL.
Aerobic energy cost in specific locomotion and mechanical efficiency on bicycle ergometer
At post-1, ECspe was unchanged in HL and in LL (P = 0.81, F = 0.1). The decrease in ECspe between post-1 and post-15 was significant in both groups (HL: −7.0 ± 12.6%, P < 0.05; LL: −6.8 ± 12.0%, P < 0.05) and with no significant difference between HL and LL (P = 0.97, F = 0.1). The increase in MEbic between pre- and post-15 was not significantly different between HL and LL (P = 0.56, F = 0.35). However, at post-15, MEbic was higher than at pre-only in HL (+ 8.2 ± 12.5%, P < 0.01).
Hematological variables
The change in hemoglobin was significant between pre- and post-1 only in HL (P < 0.01), but the magnitude of the change was similar in HL and LL (P = 0.60, F = 0.2). Hematological results have been shown in two previous studies, Robach et al. (2006) for the swimmers, and Brugniaux et al. (2006) for the runners.
Discussion
The major finding of the current study was that bringing together 40 elite endurance athletes of different sports confirmed the efficacy of the Hi–Lo model: PPO increased to a greater extent in HL than in LL immediately and fifteen days after the training period. Some submaximal variables as \(\dot{V}\hbox{O}_{\rm 2RCP}, {P}_{\rm RCP},\) increased more in HL than in LL. During the post-altitude seal-level training period, the improvement in non-specific (8–5%) and specific (7–7%) economy was important in both groups but not significantly different between the groups.
Methodological limitations
The same experimental design was not exactly replicated in the three experiments. Due to the characteristics of the subjects, the second and third experiments (swimmers and runners) were slightly different from the first one (cross-country skiers) by limiting the altitude level to 3,000 m. Indeed the 6 days of hypoxic exposure to 3,500 m probably caused a condition of fatigue in the Nordic-skiers: we noted a decrease in the neuro-vegetative activity analyzed by heart rate variability (unpublished data). We thus decided to limit to a maximum altitude of 3,000 m for the two following experiments. This was in accordance with one of the aims of this study elected by the IOC on the impact of these hypoxic methods on health. Due to the nature of this study, funded by the IOC, the subjects were all elite and the training program was part of an actual training season. It is of interest that, before Hi–Lo, no difference was shown between HL and LL groups. The authors think that is a very strong point of the present study, although some differences in training were obviously observed between the three sports in term of intensity and volume. Similarly, some testing protocols were adapted to reflect the specificity of the sport practiced by the subjects. Nevertheless, a sub-maximal cycling test was common for all subjects in order to compare the economy of exercise. One might also argue that the \(\dot{V}\hbox{O}_{\rm 2max}\) values measured in so-called ‘elite athletes’ were low, around 60 ml min−1 kg−1. Several reasons may explain this fact: (1) all the exercise procedures were performed at the altitude of 1,200 m, which is known to reduce sea-level \(\dot{V}\hbox{O}_{\rm 2max}\) by 5–6% (Robergs et al. 1998); (2) even if all the subjects involved in this study were elite athletes, most of the selected sport specialities (i.e. Nordic-combined skiers, swimmers, runners) did not necessarily correspond to the highest levels of \(\dot{V}\hbox{O}_{\rm 2max}\) found in the literature, as it is the case for cycling or cross-country ski (in the present study, the two cross-country ski subjects exhibited \(\dot{V}\hbox{O}_{\rm 2max}\) of ∼70 ml min−1 kg−1 at 1,200 m); (3) the female athletes represented 20% of the subjects; (4) the studied periods were at minimum two months prior the competitive period and the \(\dot{V}\hbox{O}_{\rm 2max}\) values of the athletes were probably not the highest of the season.
Immediate post-Hi–Lo effects
In the current study, if \(\dot{V}\hbox{O}_{\rm 2max}\) and hemoglobin were increased to the same extend in HL and LL, PPO increased more in HL. These results confirm the efficacy of Hi–Lo, but not solely mediated by an increase in hemoglobin as reported by Levine and Stray-Gundersen (1992, 1997). This suggests different muscular adaptations between the two groups. Although it is clear that PPO is strongly related with \(\dot{V}\hbox{O}_{\rm 2max}\) (Hawley and Noakes 1992), it is known that \(\dot{V}\hbox{O}_{\rm 2max}\) and performance may be modified differently after sea level (Acevedo and Goldfarb 1989) or Hi–Lo (Levine and Stray-Gundersen 1997) training. In the study of Levine and Stray-Gundersen (1997), after 4 weeks of altitude training, the Hi–Lo group (training at 1,250 m of altitude and nights at 2,500 m) increased the level of \(\dot{V}\hbox{O}_{\rm 2max}\) by 5% in the same way that the Hi–Hi group (training at 2,500 m of altitude and nights at 2,500 m), whereas the performance on 5,000 m for these runners was improved only in the Hi–Lo group. The potential mechanisms for such a difference could come from muscular adaptations such as improvement in the anaerobic capacities, increase in the enzymatic pool of anaerobic glycolysis or improvement of the muscular buffer capacity (Gore et al. 2001; Gore and Hopkins 2005). The current study demonstrated that 15–18 days of Hi–Lo with 14–16 h per 24 h of hypoxic stimulation was sufficient to induce a greater improvement in several physiological variables \((\hbox{PPO}, {P}_{\rm RCP}, \dot{V}\hbox{O}_{\rm 2RCP})\) than the same training performed at low altitude. One may argue that the effects would have been even more important with a longer Hi–Lo period. Indeed, Levine and Stray-Gundersen (1997, 2005) suggested that 4 weeks was the ideal duration for Hi–Lo. However, a continuous altitude training camp of 4 weeks is unlikely in elite athletes who have a busy training and competition program. The current trend is to replicate several 2–3 weeks camp during a season, instead of a single longer one. Another major difference with most of the previous Hi–Lo studies relates to the intensity of training. It has been advocated that one of the key mechanism of Hi–Lo relies on the maintenance of an usual training intensity (Levine and Stray-Gundersen 1992, 1997). In the current study, the efficiency of Hi–Lo is confirmed despite a mainly aerobic training programme (70% of training were at an intensity lower than VT and 93% than RCP). The authors support the view that the intensity of training is paramount but believe that the current results complement the previous findings of Levine and Stray-Gundersen (1992, 1997), by showing that Hi–Lo induces a greater improvement even with a moderate intensity of training.
Delayed post-Hi–Lo effects
\(\dot{V}\hbox{O}_{\rm 2max}\) and PPO
Fifteen days after Hi–Lo, \(\dot{V}\hbox{O}_{\rm 2max}\) and hemoglobin were returned to pre-values while PPO was maintained. The similar time course of change in \(\dot{V}\hbox{O}_{\rm 2max}\) and hemoglobin is not surprising since \(\dot{V}\hbox{O}_{\rm 2max}\) is a function of the O2 transport system, itself mainly dependant on cardiac output and O2 content in trained athletes (e.g. Wagner 2000). In the same way, di Prampero (2003) proposed a model for quantifying the respective influences of the limiting factors of \(\dot{V}\hbox{O}_{\rm 2max}\) in human. He showed that in exercise with large muscle groups, like running or cycling, the main factor limiting \(\dot{V}\hbox{O}_{\rm 2max}\) is the O2 transport capacity, product of maximal cardiac output and the O2 transport coefficient for blood. This factor is responsible for 70–75% of the factors limiting \(\dot{V}\hbox{O}_{\rm 2max}\) downstream of the lung and, for example, an increase of 10% of this factor lead an increase of about 7% of \(\dot{V}\hbox{O}_{\rm 2max}.\) Following this model, since in the current study, we noted at post-1 an increase in \(\dot{V}\hbox{O}_{\rm 2max}\) of 7.8% in HL group, we can assume that the increase in O2 transport capacity was important (> 10%). As PPO remained improved 15 days after Hi–Lo while \(\dot{V}\hbox{O}_{\rm 2max}\) was returned to the pre-values, the hypothesis that PPO was improved mainly by local muscular adaptations is reinforced (Acevedo and Goldfarb 1989; Gore et al. 2001).
Respiratory compensation point
Fifteen days after Hi–Lo, the submaximal factors of aerobic performance were improved, especially \(\dot{V}\hbox{O}_{2}\) and power at respiratory compensation point. di Prampero (1986) showed that aerobic power at intensities below \(\dot{V}\hbox{O}_{\rm 2max}\) is the product of three components: \(\dot{V}\hbox{O}_{\rm 2max},\) the fraction of \(\dot{V}\hbox{O}_{\rm 2max}\) representing exercise intensity \((\dot{V}\hbox{O}_{\rm 2fracmax})\) and the amount of energy per unit of distance. In the current study, 15 days after Hi–Lo, there was no increase in \(\dot{V}\hbox{O}_{\rm 2max}.\) So, in agreement with the model of di Prampero, we speculate that the increase in \(\dot{V}\hbox{O}_{\rm 2RCP}\) induced the increase in submaximal aerobic power. Gore et al. (2001) have shown that, after 23 nights of Hi–Lo, exercise efficiency was increased, mainly because of an improved muscle buffer capacity. These authors, skeptical about the efficiency of the Hi–Lo method, suggested that this latter mechanism was among the main adaptations to Hi–Lo (Gore and Hopkins 2005). It has been also shown, that endurance performance was in strong relationship with the level of \(\dot{V}\hbox{O}_{2}\) and the power at onset of blood lactate accumulation (e.g. Tanaka 1990). OBLA and RCP are located at levels of identical power and it was shown that the lactic acidosis is in causal relation with the hyperventilation which starts with RCP (Meyer et al. 2004). Our study shows, in agreement with the literature, that the changes in endurance performance are in relation with the changes in submaximal \(\dot{V}\hbox{O}_{2}\) and power at RCP. It shows that one of the important effects of Hi–Lo can be observed 15 days after the end of the training session in hypoxia.
Energy cost
One of the major findings of the present study is the important improvement of economy of work during the post-altitude period of sea-level training (between post-1 and post-15), whereas no change in economy was observed during the altitude training period (between pre- and post-1). This question of altitude effect on economyis on heart of apassionate debate (Levine and Stray-Gundersen 2005; Gore and Hopkins 2005). As shown by di Prampero (1986) the velocity in human locomotion depends on \(\dot{V}\hbox{O}_{\rm 2max},\) fractional utilization of \(\dot{V}\hbox{O}_{\rm 2max}\) but also on EC, defined as the amount of energy spent per unit of distance. It was reported that EC is correlated with performance in most of the aerobic sports, i.e. swimming (Toussaint and Hollander 1994), running (Saunders et al. 2004), or cross-country skiing (Millet et al. 2003). EC has been reported as more predictive of aerobic performance than \(\dot{V}\hbox{O}_{\rm 2max}\) in homogeneous groups of athletes (Morgan et al. 1989). The present efficiency or economy values were in line with those reported in the literature: for example, Moseley and Jeukendrup (2001), Luhtanen et al. (1987) reported mean value of cycling net efficiency of 22% that is similar to our values ranging between 22 and 25.5%. In running, di Prampero (1986) reported EC of 3.16 kJ kg−1 km−1 at a 0%-grade slope and of 8 kJ kg−1 km−1 at a 20%-grade slope. Our values of EC ranged between 231 and 256 ml kg−1 km−1 (4.9–5.4 kJ kg−1 km−1) at a 8%-grade slope in high-level runners. Similarly, in swimming, Kjendlie et al. (2004) showed mean values of EC of 36.5 ml m−1 in adult swimmers that are comparable with the present values ranging between 30.1 and 36.6 ml m−1 in elite swimmers (Table 5).
Energy cost did not change during the training period at mild altitude possibly because of the combination of positive effects as a decrease in ventilation at a identical intensity of exercise or a greater oxidation of carbohydrate (Saunders et al. 2004) and of negative ones as the increase in \(\dot{V}\hbox{O}_{2}\) or the fatigue induced by the additional hypoxic stress, as energy cost was shown to increase under the influence of fatigue (Morgan et al. 1989). However 15 days later, the energy cost was reduced in the non-specific activity (i.e. cycling) only in the Hi–Lo group, and in their specific activity in both groups. Since the two groups trained similarly at 1,200 m and had the same recovery post-stage training period, it is not surprising to observe no difference in specific aerobic energy cost between groups and when compared to pre-values. The mild hypoxic environment (1,200 m) could have modified the specific neuromuscular pattern of these elite subjects who used to train only at sea level (swimmers and runners) (it is less true for the cross-country skiers, but the training between post-1 and post-15 was performed at sea-level).
The persistence of some physiological adaptations induced by altitude after 8 days (Beidleman et al. 1997) or 1 month (Katayama et al. 2005) has already been reported. In the present study, it may be that at post-15 the athletes in the HL group may have acquired a cumulative effect of both the altitude-induced improvements in economy of exercise and power at RCP and the sea-level decrease in exercise energy cost observed in the two groups. These results suggest that the time period of 2–3 weeks generally recommended by coaches between an altitude training sojourn and a sea-level competition is an adequate resultant of these two mechanisms.
Conclusion
The current study confirms the efficiency of the Hi–Lo method by demonstrating that elite athletes in different endurance sports improved maximal (PPO) and submaximal \((P_{\rm RCP}, \dot{V}\hbox{O}_{\rm 2RCP})\) parameters of performance in Hi–Lo to a greater extent than with a similar Lo–Lo training program. In addition, this study provides some novel findings regarding the post-altitude period by showing the important increase in the economy of work when returning to sea-level.
References
Acevedo EO, Goldfarb AH (1989) Increased training intensity effects on plasma lactate, ventilatory threshold, and endurance. Med Sci Sports Exerc 21:563–568
Ashenden MJ, Gore CJ, Dobson GP, Hahn AG (1999) “Live high, train low” does not change the total haemoglobin mass of male endurance athletes sleeping at a simulated altitude of 3000 m for 23 nights. Eur J Appl Physiol 80:479–484
Banister EW, Calvert TW, Savage MV, Bach TM (1975) A systems model of training for athletic performance. Aust J Sports Med 7:57–61
Beidleman BA, Muza SR, Rock PB, Fulco CS, Lyons TP, Hoyt RW, Cymerman A (1997) Exercise responses after altitude acclimatization are retained during reintroduction to altitude. Med Sci Sports Exerc 29:1588–1595
Brugniaux JV, Schmitt L, Robach P, Nicolet G, Fouillot JP, Moutereau S, Lasne F, Pialoux V, Saas P, Chorvot MC, Cornolo J, Olsen NV, Richalet JP (2006) Eighteen days of “Living High–Training Low” stimulate erythropoiesis and enhance aerobic performance in elite middle-distance runners. J Appl Physiol 100:203–211
Busso T, Denis D, Bonnefoy R, Geyssant A, Lacour JR (1997) Modeling of adaptations to physical training by using a recursive least squares algorithm. J Appl Physiol 82:1685–1693
di Prampero PE (1986) The energy cost of human locomotion on land and in water. Int J Sports Med 7:55–72
di Prampero PE (2003) Factors limiting maximal performance in humans. Eur J Appl Physiol 90:420–429
Dick FW (1992) Training at altitude in practice. Int J Sports Med 13:203–206
Gore CJ, Hopkins WG (2005) Counterpoint: positive effects of intermittent hypoxia (live high:train low) on exercise performance are not mediated primarily by augmented red cell volume. J Appl Physiol 99:2055–2057
Gore CJ, Hahn AG, Aughey RJ, Martin DT, Ashenden MJ, Clark SA, Garnham AP, Roberts AD, Slater GJ, McKenna MJ (2001) Live high: train low increases muscle buffer capacity and sub-maximal cycling efficiency. Acta Physiol Scand 173:275–286
Green HJ, Roy B, Grant S, Hughson R, Burnett M, Otto C, Pipe A, McKenzie D, Johnson M (2000) Increases in submaximal cycling efficiency mediated by altitude acclimatization. J Appl Physiol 89:1189–1197
Hawley JA, Noakes TD (1992) Peak power output predicts maximal oxygen uptake and performance time in trained cyclists. Eur J Appl Physiol Occup Physiol 65:79–83
Ingjer F, Myhre K (1992) Physiological effects of altitude training on elite male cross-country skiers. J Sports Sci 10:37–47
Katayama K, Fujita H, Sato K, Ishida K, Iwasaki K, Miyamura M (2005) Effect of a repeated series of intermittent hypoxic exposures on ventilatory response in humans. High Alt Med Biol 6:50–59
Keskinen KL, Rodriguez FA, Keskinen OP (2003) Respiratory snorkel and valve system for breath-by-breath gas analysis in swimming. Scand J Med Sci Sports 13:322–329
Kjendlie PL, Ingjer F, Stallman RK, Stray-Gundersen J (2004) Factors affecting swimming economy in children and adults. Eur J Appl Physiol 93:65–74
Levine BD, Stray-Gundersen J (1992) A practical approach to altitude training: where to live and train for optimal performance enhancement. Int J Sports Med 13:209–212
Levine BD, Stray-Gundersen J (1997) “Living high-training low”: effect of moderate-altitude acclimatization with low-altitude training on performance. J Appl Physiol 83:102–112
Levine BD, Stray-Gundersen J (2005) Point: positive effects of intermittent hypoxia (live high:train low) on exercise performance are mediated primarily by augmented red cell volume. J Appl Physiol 99:2053–2055
Luhtanen P, Rahkila P, Rusko H, Viitasalo JT (1987) Mechanical work and efficiency in ergometer bicycling at aerobic and anaerobic thresholds. Acta Physiol Scand 131:331–337
Melissa L, Macdougall JD, Tarnopolsky MA, Cipriano N, Green HJ (1997) Skeletal muscle adaptations to training under normobaric hypoxic versus normoxic conditions. Med Sci Sports Exerc 29:238–243
Meyer T, Faude O, Scharhag J, Urhausen A, Kindermann W (2004) Is lactic acidosis a cause of exercise induced hyperventilation at the respiratory compensation point? Br J Sports Med 38:622–625
Millet GP, Boissiere D, Candau R (2003) Energy cost of different skating techniques in cross-country skiing. J Sports Sci 21:3–11
Morgan DW, Martin PE, Krahenbuhl GS (1989) Factors affecting running economy. Sports Med 7:310–330
Moseley L, Jeukendrup AE (2001) The reliability of cycling efficiency. Med Sci Sports Exerc 33:621–627
Robach P, Schmitt L, Brugniaux JV, Roels B, Millet G, Hellard P, Nicolet G, Duvallet A, Fouillot JP, Moutereau S, Lasne F, Pialoux V, Olsen VN, Richalet JP (2006) Living high–training low: effect on erythropoiesis and aerobic performance in highly-trained swimmers. Eur J Appl Physiol 96:423–433
Robergs RA, Qunintana R, Parker DL, Frankel CC (1998) Multiple variables explain the variability in the decrement in \(\dot{V}\hbox{O}_{\rm 2max}\) during acute hypobaric hypoxia. Med Sci Sports Exerc 30:869–879
Saunders PU, Telford D, Pyne B, Cunningham RB, Gore J, Hahn G, Hawley A (2004) Improved running economy in elite runners after 20 days of simulated moderate-altitude exposure. J Appl Physiol 96:931–937
Tanaka K (1990) Lactate-related factors as a critical determinant of endurance. Ann Physiol Anthropol 9:192–202
Telford RD, Graham KS, Sutton JR, Hahn AG, Campell DA, Creighton SW, Cunningham RB, Davis PG, Gore CJ, Smith JA, McA Tumilty D (1996) Medium altitude training and sea-level performance. Med Sci Sports Exerc 28:S124
Toussaint HM, Hollander AP (1994) Energetics of competitive swimming. Implications for training programs. Sports Med 18:384–405
Wagner PD (2000) News ideas on limitations to \(\dot{V}\hbox{O}_{\rm 2max}.\) Exerc Sport Sci Rev 28:10–14
Acknowledgments
The authors acknowledge the International Olympic Committee and the French Ministry of Sports for their financial support, the swimming team of Brest and its coach, Thierry Boulonnois, the French Swimming Federation, the French Ski Federation and the national coaches of the French biathlon team, Pascal Etienne and Christophe Vassallo, the French Athletism Federation and the national coaches of middle distance running, Roger Milhau and Patrice Binelli.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Schmitt, L., Millet, G., Robach, P. et al. Influence of “living high–training low” on aerobic performance and economy of work in elite athletes. Eur J Appl Physiol 97, 627–636 (2006). https://doi.org/10.1007/s00421-006-0228-3
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-006-0228-3