
Abstract
Critical power (CP) and the second ventilatory threshold (VT2) are presumed to indicate the power corresponding to maximal lactate steady state (MLSS). The aim of this study was to investigate the use of CP and VT2 as indicators of MLSS. Eleven male trained subjects [mean (SD) age 23 (2.9) years] performed an incremental test (25 W·min−1) to determine maximal oxygen uptake (V̇O2max), maximal aerobic power (MAP) and the first and second ventilatory thresholds (VT1 and VT2) associated with break points in minute ventilation (V̇E), carbon dioxide production (V̇CO2), V̇E/V̇CO2 and V̇E/V̇O2 relationships. Exhaustion tests at 90%, 95%, 100% and 110% of V̇O2max and several 30-min constant work rates were performed in order to determine CP and MLSS, respectively. MAP and V̇O2max values were 344 (29) W and 53.4 (3.7) ml·min−1·kg−1, respectively. CP [278 (22) W; 85.4 (4.8)% V̇O2max] and VT2 power output [286 (28) W; 85.3 (5.6)% V̇O2max] were not significantly different (p=0.96) but were higher (p<0.05) than the MLSS work rate [239 (21) W; 74.3 (4.0)% V̇O2max] and VT1 power output [159 (23) W; 52.9 (6.9)% V̇O2max]. MLSS work rate was significantly correlated (p<0.05) with those noted at VT1 and VT2 (r=0.74 and r=0.93, respectively). VT2 overestimated MLSS by 10.9 (6.3)% V̇O2max which was significantly higher than VT1 [+21.4 (5.6)% V̇O2max; p<0.01]. CP calculated from a given range of exhaustion times does not correspond to MLSS.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Thanks to advances in the field of physiology, coaches can now design training programs at many specific work intensities and measure their effect on aerobic performance. For example, the intensity that demarcates the transition from moderate to heavy exercise can theoretically be maintained indefinitely, i.e. for a long period of time without fatigue (Kinderman et al. 1979). Numerous direct and indirect methods have been proposed (McLellan and Skinner 1981) to determine this balance intensity and several investigations have been conducted to assess the level of agreement between the different intensities estimated (Housh et al. 1991; McLellan and Cheung 1992; Smith and Jones 2001). Consequently, multiple discordant results have been obtained leading to confusion and controversy.
Estimated from several sub-maximal constant workload tests, the maximal lactate steady state (MLSS) has been defined as the highest work rate that can be maintained over time without continual blood lactate accumulation (Hollman 1959; Heck et al. 1985; Beneke 1995). Above MLSS, an increase of blood lactate is observed, because the anaerobic component of the body's metabolic processes becomes greater (Hill et al. 1924). As early as 1930, Owles observed a concomitant increase of ventilation, carbon dioxide (CO2) excretion, and blood lactate, and Harrison and Pilcher (1930) then proposed that the rise in CO2 excretion was due to bicarbonate buffering of the acids that were produced during such exercises. Working on this assumption, Wasserman and McIlroy (1964) were able to avoid invasive measurements of blood lactate, and based their detection of the onset of metabolic acidosis (anaerobic metabolism) purely on ventilatory, or non-invasive measurements. Accordingly, the measurement of various ventilatory and/or gas exchange indices within a single incremental load test enables the detection of two ventilatory thresholds. The first one, called the first ventilatory threshold or ventilatory adaptation to CO2 production (V̇CO2) threshold (VT1), corresponds to the first non-linear increases in V̇CO2 and ventilation due to the bicarbonate buffering of H+ in response to the systematic increase of blood lactate above resting values (Wasserman et al. 1973). On the other hand, the second threshold (VT2), also named the respiratory compensation threshold or threshold of uncoupling of ventilation from V̇CO2, can be identified as the second breakpoint in the ventilation response mainly explained by an acidosis (pH decrease) as bicarbonate is overwhelmed by the growing production of lactate (Vallier et al. 2000). Following the publication of Wasserman and McIlroy (1964), the use of ventilation/gas exchange techniques to quickly determine the onset of metabolic acidosis has become widespread in the literature and has an obvious attraction in cardio-respiratory evaluations as a non-invasive and useful index of exercise performance (Vallier et al. 2000).
Furthermore, critical power (CP) is a theoretical concept expected to provide an accurate estimation of the maximal work rate in a state of physiological aerobic balance (Moritani et al. 1981). In 1960, Scherrer and Monod presented this concept based on the linear relationship between total work (W lim) and time to exhaustion (T lim), the slope representing CP. From their investigation on local dynamic muscle work, these authors have stated that exercise at power outputs equal to or less than CP could theoretically be maintained "for a very long period of time without fatigue". Since Scherrer and Monod (1960), several studies have been conducted to apply this concept to whole-body exercises on an ergocycle (Moritani et al. 1981), a treadmill (Hughson et al. 1984) or whilst swimming (Wakayoshi et al. 1993). Nevertheless, even if CP remains a theoretical and indirect means of estimating MLSS, numerous investigations have attempted to compare this parameter with other indices of aerobic fitness (Poole et al. 1988; Housh et al. 1991; McLellan and Cheung 1992; Smith and Jones 2001), leading to discrepancies that remain to be elucidated.
Interestingly, most studies conducted to compare the direct and indirect methods of determining of MLSS on a cycle ergometer included imposed pedal rates in their protocols (Housh et al. 1991; McLellan and Cheung 1992). Nevertheless, several authors have reported that pedal rates could influence the cardiorespiratory and efficiency responses to exercise (Vandewalle et al. 1997; Weissland et al. 1997) as well as the measurement of VT2, CP or MLSS (Hill et al. 1995). Consequently, some authors have recommended using optimal pedal rates (Takano 1988), which are close to the spontaneously chosen pedal rates (Weissland et al. 1997).
No investigation has been conducted to compare, within a single population, VT2, CP and MLSS using an ergometer and allowing subjects to perform, at spontaneously chosen pedal rates, as they can do in sport activities such as cycling. Hence, the purpose of this study was to assess the relationship between MLSS, VT2 and CP with subjects exercising at their chosen cadence on a cycle ergometer.
Methods
Subjects
Eleven healthy and well-trained male students [mean aged 23 (2.9) years] volunteered for this study. Mean height and body mass were 1.8 (0.1) m, and 69.2 (5.1) kg, respectively. Before the test protocol, all the subjects were informed of the procedure, gave written consent to take part in the study according to the French National Committee for Clinical Research and underwent a complete medical examination.
Experimental design and general procedures
Each subject performed three sessions of different tests as follows:
-
1.
An incremental maximal test (Tmax) in order to measure the maximal oxygen uptake (V̇O2max) and to determine the maximal aerobic power (MAP), and the ventilatory thresholds (VT1 and VT2).
-
2.
Three to four 30-min constant load tests set at power outputs ranging from 65% to 85% V̇O2max (T30) in order to determine MLSS and the corresponding blood lactate concentration ([La]MLSS).
-
3.
Four all-out tests set at 90%, 95%, 100% and 110% V̇O2max (T90, T95, T100 and T110) in order to calculate the CP.
Before the initial test, each subject was given preliminary instructions on how to complete the different tests and was allowed to practise. They were instructed not to do any strenuous exercises the day before each test in order to avoid a possible effect on their physiological responses. Except for Tmax, all the tests were randomly assigned and performed within a 3-week period, during which time the subjects reduced their training.
All the tests were performed on an ergometrics "er800S". This ergometer allows subjects to perform with a constant power regime. Indeed, the power output of the device was controlled throughout the exercise by increasing or decreasing resistance based on the evolution of the user's pedal rate. Thus, subjects were allowed to pedal at a self-selected cadence. The seat height was set to 100% of trochanteric length and adjusted to provide a slight bend of 10° in the knee at full extension.
Determination of V̇O2max and VT
The maximal incremental test was preceded by a 3-min warm-up period at 75 W. This initial work rate was then increased by 25 W·min−1. To ensure that V̇O2max was reached, subjects were encouraged to continue as long as possible so that a levelling off in the course of oxygen uptake (V̇O2). The test ended at the point of voluntary exhaustion. During the test, the subjects breathed through a facemask. V̇O2, V̇CO2, minute ventilation (V̇E) and respiratory frequency (f) were continuously measured using an open circuit system (CPX Medical Graphics, St. Paul, Minnesota, USA). A 10-lead ECG (Quinton Q 810, Seattle, Washington, USA) was recorded continuously during tests to determine heart rate (HR). The mean respiratory exchange ratio (RER), and ventilatory equivalents in O2 (V̇E/V̇O2) and CO2 (V̇E/V̇CO2) values were calculated from the recorded measurements. The maximal values of V̇O2, HR, V̇CO2, RER and VE attained during the test were reported (V̇O2max, HRmax, V̇CO2max, RERmax, VEmax). When no plateau of V̇O2 was achieved, MAP was identified as the peak power output, i.e. the maximal exercise intensity maintained during the last stage. When a plateau of V̇O2 was achieved, MAP was defined as the minimal exercise intensity that could elicit V̇O2max. A plateau of V̇O2 in the V̇O2–exercise intensity relationship was defined as an increase in V̇O2 of less than 1.5 ml·min−1·kg−1 with the increase in exercise intensity. The single indices used individually in order to determine VT1 and VT2 for each subjects were V̇E, V̇CO2, V̇E/V̇O2 and V̇E/V̇CO2. The following criteria were employed in selecting the thresholds. According to Wasserman et al. (1973), VT1 was defined as the minimal load at which V̇E/V̇O2 exhibited a systematic increase without a concomitant increase in V̇E/V̇CO2. VT2 corresponded to the minimal work rate at which the increase in V̇E/V̇O2 was accompanied by an increase of V̇E/V̇CO2 (Wasserman and McIlroy 1964). Beaver et al. (1986) defined VT1 and VT2 as the work rates associated with a first and a second non-linear increase of VE and V̇CO2. According to the criteria outlined above, three independent investigators blindly reviewed the plots of each index and made individual determinations of VT1 and VT2.
Determination of MLSS
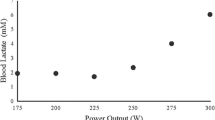
In the first 30-min constant load test (T30), the work rate was fixed at 75% V̇O2max. Another 30-min work rate test with a 5% V̇O2max higher work rate was performed on a separate day if, during the test, a steady state or a decrease of the blood lactate concentration ([La]) was observed. Subsequent 30-min constant work rate tests were increased by 5% V̇O2max each time until no steady-state [La] could be maintained. In contrast, if, during the first 30-min test set at 75% V̇O2max, an increase of [La] was observed, subsequent 30-min tests decreased by 5% V̇O2max each test until a steady state or a decrease of [La] could be maintained (Fig 1). According to the procedure published by Heck et al. (1985), MLSS work rate was defined as the highest work rate that could be maintained for 30 min without an increase of [La] by more than 1.0 mmol·l-1 within the last 20 min of the constant work rate test. [La]MLSS was calculated as the average [La] measured at the 15th, 20th, 25th and 30th min of the test (Beneke 1995).

Determination of maximal lactate steady state (MLSS) work rate in a typical subject from the change of blood lactate concentration during the 30-min sub-maximal constant load tests
Measurements of [La] were obtained from capillary blood samples. The fingertip was cleaned with an alcohol swab, dried and then punctured with an automated lancet to sample blood into capillary tubes to determine blood [La] at the beginning, at the end, and every 5th min of all the 30-min tests. Blood samples were analysed using Dr Lange's photometric method (Berlin, Germany).
Determination of CP
Prior to each test, subjects warmed up at 40% V̇O2max for a 3-min period followed by 2 min at 50% V̇O2max. Four exhaustion exercises set at 90%, 95%, 100% and 110% V̇O2max were used to calculate the CP employing the protocol described by Moritani et al. (1981). These powers were selected so exercise time to exhaustion would occur between approximately 2 and 15 min. Subjects were instructed before the rides to pedal to exhaustion and were also given verbal encouragement during the rides. The test was terminated and the time recorded to the nearest second when they simply stopped pedalling because they were so exhausted they could not pedal any more. CP was determined for each subject from a plot of accomplished work (W lim) versus exhaustion time (T lim) with the power slope of the resulting linear relationship being defined as the CP (Fig. 2).

Determination of critical power (CP) in a typical subject from the linear relationship between accomplished work (kJ) and exhaustion time (s). (T lim Time to exhaustion, W lim total work)
Analyses of the spontaneous chosen pedal cadences (SCPC)
The pedal rates were visualized only by the research worker on the screen of the computer and were noted every 15 s during the incremental test, every 30 s during the 30-min tests, and every 10 s during the exhaustion tests. These values were also verified with the values calculated from a goniometer fixed to the knee. Interpolations were realized on the SCPC–power output relationship obtained from the incremental test data in order to determine individually the mean SCPC values corresponding to 30%, 40%, 50%, 60%, 70%, 80%, 90%, and 100% MAP. Concerning the other exhaustion tests, the SCPC value was individually determined every 25% of the exhaustion time.
Statistical analysis
All the calculations were made using STATISTICA (Statsoft 2000). Standard statistical methods were used for the calculation of means and standard deviations. Normal Gaussian distribution of the data was verified by the Shapiro-Wilks test (Shapiro et al. 1968) to ensure that parametric analyses could be used. An analysis of variance with repeated measures (ANOVA) was used to test for the existence of significant differences between VT1, VT2, CP and MLSS, between the V̇O2max and HRmax measured during the all-out tests and the incremental test, and between the mean spontaneous pedal rate values reported for each test. This procedure was also used to test the changes in SCPC during each test. All data were analysed 20 s by 20 s for the exhaustion tests, minute by minute for the sub-maximal constant load tests, and very 10%MAP for the incremental test. The compound symmetry, or sphericity, was checked by the Mauchley test (Winter et al. 2001). Sphericity was defined as the assumption that, in repeated measures designs, the variance among the repeated measures is the same or nearly so (homogeneity of variance), and the relationships or correlations among all of the combinations of the repeated measures are the same or nearly so (homogeneity of covariance). When the assumption of sphericity was not met, the significance of F-ratios was adjusted according to the Greenhouse–Geisser procedure (Greenhouse and Geisser 1959). Multiple comparisons were made with the Tukey HSD post hoc test when the Greenhouse–Geisser epsilon correction factor was >0.50, or with the Bonferroni post hoc test when the epsilon factor was <0.50 (Stevens 1996). The relationships between VT1, VT2, CP and MLSS were examined by simple linear regression analyses (Bravais-Pearson coefficient). A 95% level of confidence was accepted for all comparisons.
Results
Comparisons between VT1, VT2, CP and MLSS
Values of work rates (in W, %MAP and %V̇O2max) corresponding to VT1, VT2, MLSS and CP are presented in Table 1. Mean MLSS work rate was significantly higher than mean VT1 but was significantly lower than mean CP and VT2 (p<0.05), which were not significantly different (p=0.80). Furthermore, located between VT1 and VT2 values, the mean MLSS work rate was significantly correlated with these two threshold values (r=0.68 and 0.69, respectively; p<0.05). CP was not significantly correlated with any intensity (Fig. 3). All coefficients of correlation obtained between VT1, VT2, MLSS, CP, and MAP are presented in a matrix (Table 2).

Pearson product moment correlation coefficients between CP and MLSS, MLSS and VT2 and CP and the second ventilatory threshold (VT 2 )
Tmax
MAP, V̇O2max, HRmax, and RERmax values were 345 (27) W, 50.8 (2.7) ml·min−1·kg−1, 194 (9) beats·min−1, and 1.21 (0.07), respectively. At VT1 and VT2, V̇O2, HR and RER values were 26.4 (4.4) ml·min−1·kg−1, 132 (12) beats·min−1 and 0.79 (0.06), and 42.5 (4.9) ml·min−1·kg−1, 174 (10) beats·min−1 and 1.03 (0.04), respectively.
T30
Three of the eleven subjects were not able to perform for 30 min at a work rate 5% V̇O2max higher than MLSS (23.0 min, 19.2 min and 18.2 min). For the eight other subjects, [La] values increased by more than 1 mmol·l−1 within the last 20 min of the test. [La]MLSS and HRMLSS were 5.87 (1.31) mmol·l−1 and 171 (8) beats·min−1, respectively. HRMLSS and HR measured at VT2 were not significantly different (p=0.5) and were not correlated (p=0.07).
Exhaustion times and the work–time relationship
Individual simple regression coefficients for the plot of work against time ranged from 0.99 to 1 [0.99 (0.0)]. Exhaustion times, peak V̇O2 and HR at 90%, 95%, 100% and 110% V̇O2max are presented in Table 3. V̇O2max and HRmax measured during Tmax were not significantly different from the maximal V̇O2 and maximal HR values attained during the exhaustion tests (p>0.05).
Evolutions of SCPC within and between the tests
Concerning the evolution of SCPC throughout each test, the incremental test values ranged from 82 (12) rpm at 30% MAP to 93 (11) rpm at MAP. For the 30-min tests and the exhaustion tests, the group means (±SD) and ranges of SCPC are presented in Table 4. The mean SCPC values did not change significantly (p>0.05), except for T100 and T110 during which SCPC significantly decreased during the last quarter of the tests (p<0.05). There were inter-subject differences in these SCPC, but there was no relationship between power output and SCPC. Thus, there were no significant difference (p>0.05) between the group mean SCPC values calculated for each test. The mean SCPC value was 89 (9) rpm. The variations within subjects for the various rides, namely the lowest and highest subject mean SCPC values, standard deviations, and coefficients of variation, were 77 (8) rpm and 103 (6) rpm, 4.8 and 10.4 rpm, and 5.8 and 12.9%, respectively.
Discussion
The main finding of this investigation indicates that CP and VT2 are significantly higher than MLSS. These two parameters do not represent the maximal intensity that can be maintained for a long period of time without a continuous increase of blood [La].
Concerning CP, the speed slope of the distance–time relationship corresponds to the speed asymptote of the speed–time relationship, which is thought to represent an intensity that can theoretically be sustained indefinitely. However, it is well known that because of factors such as substrate depletion, temperature regulation, and fluid and electrolyte balance, which may contribute to fatigue during prolonged exercises, this mathematical expression is unlikely to provide a precise representation of an infinitely sustainable speed (Poole et al. 1988). Authors then proposed to think of this parameter in terms of an intensity that can be maintained for a prolonged period of time and for which V̇O2 and [La] can be stabilized (Gaesser and Wilson 1988; Poole et al. 1988). Unfortunately, in agreement with the present results, some authors reported exhaustion times at CP ranging from 18 min to 60 min (Housh et al. 1989; Jenkins and Quigley 1990; McLellan and Cheung 1992; Brickley et al. 2002), and accumulation of blood lactate during exercises performed at CP (McLellan and Cheung, 1992; Brickley et al. 2002). Moreover, in most of these investigations, the exhaustion tests were performed using constant cadence (Poole et al. 1988; Jenkins and Quigley 1990; Housh et al. 1989). Nevertheless, the methodology enabling subjects to choose their preferred cadence, as in the study of McLellan and Cheung (1992), reflects the actual sporting activity more accurately. The spontaneously chosen cadences are also related to the optimal cadence (Brisswalter et al. 1999) and react to a decrease in cardiorespiratory parameters (Weissland et al. 1997, 1999) or neuromuscular fatigue (Takaishi et al. 1994; Vercruyssen et al. 2001); indeed, the use of SCPC should improve the determination not only of CP, but also of MLSS, VT1 and VT2. Accordingly, several authors have reported that pedal rates can influence the measurement of CP (Carnevale and Gaesser 1991; Hill et al. 1995) and VT1 and VT2 (Hughes et al. 1982; Takano 1988). The results of the present study highlighted that even if the mean SCPC value did not change significantly between and throughout tests (except for T100 and T110 during which SCPC significantly decreased during the last quarter of the test), there were inter-subject differences in these SCPC, with the lowest and highest subject mean SCPC values equal to 77 (8) rpm and 103 (6) rpm. Each subject had the opportunity to choose the pedal cadence that he preferred and felt as optimal. This is all the more important for well-trained subjects. Hill et al. (1995) investigated the effect of pedal rate on parameters of the hyperbolic power–time relationship, and advised researchers to be careful when comparing results among studies using different pedal rate methodologies. Thus, according to Brickley et al. (2002) and Pringle and Jones (2002), CP overestimates the maximal intensity that can be maintained for a long period of time without a continuous increase of blood [La], whatever the pedal rate methodology chosen by the investigators. Conversely, an overestimation of CP engendered by the employed methodology (Hill 1993) can explain the subjects' inability to perform at this particular work rate for a long period of time. Indeed, the slope of the work–exhaustion time relationship is considerably dependent on the duration of predictive trials because of the strict non-linearity of the work–exhaustion times relationship (Hill 1993; Vandewalle et al. 1997). The lower the exhaustion time, the greater the slope, and the lower the y-intercept (Bishop et al. 1998; McLellan and Cheung 1992). In this study, the exhaustion times averaged between 3.7 (2.3) min and 13.6 (4.4) min and are within the range (3–20 min) recommended by Bishop et al. (1998) to avoid an overestimation of CP.
To date, only two investigations (Smith and Jones 2001; Pringle and Jones 2002) have been conducted to compare CP with the directly and independently determined MLSS. Smith and Jones (2001) did not find any significant difference between these variables during treadmill running and explained the lack of correlation (r=0.62) by the heterogeneous fitness of the subjects. Conversely, Pringle and Jones (2002) obtained a higher mean CP value but it was strongly correlated with mean MLSS (r=0.95; p<0.01). The first authors concluded that CP and MLSS probably represent the same physiological phenomenon whereas the second study accounted for their results by saying that MLSS, and not CP, represents the upper limit of the heavy exercise domain in cycling. In the present study, no correlation has been obtained between CP and MLSS (r=−0.11; p=0.76) despite the homogeneous fitness of the subjects (low standard deviations of all mean values). The results of this study highlighted that CP determination leads to an overestimation of the metabolic rate associated with MLSS, and that CP and MLSS are assumed to be different physiological parameters during cycling on an ergocycle. The CP concept should not be generalized to whole-body exercises on an ergocycle, especially in light of original work looking at local dynamic muscle work (Scherrer and Monod 1960). Using this parameter to estimate aerobic endurance seems to be irrelevant.
Contrary to CP, VT2 is significantly correlated with MLSS (r=0.67; p=0.02), indicating that VT2 overestimates MLSS by 10.9 (6.3)% V̇O2max which is significantly higher than VT1 [+21.4 (5.6)% V̇O2max; p<0.01]. Such results have to be related to those of Beneke (1995), who found that, in rowing, the onset of blood lactate accumulation (OBLA) and the individual anaerobic threshold (IAT) are significantly higher than MLSS and correlated with this parameter (r=0.79 and r=0.76, respectively). These results are consistent with the physiological mechanisms underlying the emergence of VT2. Indeed, the respiratory compensation threshold is identified as a second ventilatory breakpoint following a short period of isocapnic buffering, in which V̇E increases in direct proportion to V̇CO2 during fast incremental tests. Above this point, V̇E increases at a faster rate than V̇CO2 to achieve respiratory compensation for the metabolic acidosis. Since the adaptation of V̇E is regulated by peripheral chemoreceptors that are thought to detect the liberation of large amounts of "non-metabolic" CO2 due to the buffering of protons liberated by the lactic acid dissociation, V̇E drifts and blood lactate production should be linked (Harrison and Pilcher 1930). This could explain the significant correlation obtained between VT2 and MLSS (r=0.67; p=0.02) when definitions of these parameters are considered. Thus, VT2 and MLSS seem physiologically related. Even if the difference between VT2 and MLSS cannot be explained by the present study, several explanations can be postulated. First, the two different types of exercises used to determine MLSS (sub-maximal constant load exercises) and the transition threshold (incremental exercises) should not require similar physiological and psychological adaptations to the task constraint, and thus should not create a similar stress. Second, determinations of MLSS and VT2 depend on the responses of different physiological variables requiring blood lactate and ventilatory measurements, respectively, and the mechanisms by which their responses may be linked are still being debated (Brooks 1985; Davis 1985; Loat and Rhodes 1993). Consequently, even if estimating MLSS from VT2 (MLSS equal to VT2 minus 10% V̇O2max) in the laboratory has its attractions, any such results would have to be prudently applied. The controversies concerning the concept of ventilatory thresholds are supported by the discordances between the present results and those of Yamamoto et al. (1991). Accordingly, these authors observed a lactate steady state and an increase of the blood [La] between the 15th and 30th min of two 30-min exercises set at VT2 [61.3 (6.0)% V̇O2max] and at 4.9% above VT2, respectively. The mean VT2 obtained by Yamamoto et al. (1991) is noticeably lower than that reported in the present study [85.4 (4.8)% V̇O2max] whereas the methods used to determine VT2 and the characteristics of the two populations seem to be similar.
No significant difference and correlation (r=0.07; p=0.84) between CP [85.4 (4.8)% V̇O2max] and VT2 [85.3 (5.6)% V̇O2max] were obtained. These results support those obtained previously, showing that CP and MLSS do not represent the same physiological phenomenon and that VT2 and MLSS are physiologically related. In contrast, Moritani et al. (1981) reported that CP [67.8 (9.2)% V̇O2max for male subjects and 60.1 (5.4)% V̇O2max for female subjects] was not significantly different to VT1 [62.8 (8.8)% V̇O2max for male subjects and 65.0 (5.7)% V̇O2max for female subjects] and that these two work rates were related. No explanation has been put forward to elucidate these disparities of results, notably the lower CP values obtained by these authors.
Conclusion
In conclusion, since CP and VT2 were significantly higher than the directly determined MLSS, these values do not represent the maximal work rate that can be maintained for a long period of time without a continuous rise of blood [La]. Thus, an accurate and reliable evaluation of aerobic endurance can only be realized from a direct, but long and tedious method of MLSS determination. Indeed, even if the performance of a unique incremental exercise leads to an overestimation of MLSS [VT2 equal to MLSS minus 10.9 (6.3)% V̇O2max], this method has frequently been questioned because of its unexplained underlying mechanisms and its difficult employment. Moreover, the indirect method based on the performance of several all-out tests is simple, rapid and non-invasive and, consequently, could appear particularly attractive for coaches. Regrettably, this method leads to a calculated intensity whose physiological significance remains unknown. More extensive and elaborate studies are needed to define the accurate physiological meaning of CP and the underlying mechanisms of VT2.
References
Beaver WL, Wasserman K, Whipp BJ (1986) A new method for detecting anaerobic threshold by gas exchange. J Appl Physiol 60:2020–2027
Beneke R (1995) Anaerobic threshold, individual anaerobic threshold, and maximal lactate steady state in rowing. Med Sci Sports Exerc 27:863–867
Bishop D, Jenkins DG, Howard A (1998) The critical power function is dependent on the duration of the predictive exercise tests chosen. Int J sports Med 19:125–129
Brickley G, Doust J, Williams CA (2002) Physiological responses during exercise to exhaustion at critical power. Eur J Appl Physiol 10.1007/s00421-002-0706-1
Brisswalter J, Hausswirth C, Smith D, Vercruyssen F, Vallier JM (1999) Energetically optimal cadence vs. freely-chosen cadence during cycling: effect of exercise duration. Int J Sports Med 20:60–64
Brooks GA (1985) Anaerobic threshold: review of the concept and directions for future research. Med Sci Sports Exerc 17:22–31
Carnevale TJ, Gaesser GA (1991) Effects of pedalling speed on the power-duration relationship for high-intensity exercise. Med Sci Sports Exerc 23:242–246
Davis JA (1985) Anaerobic threshold: review of the concept and directions for future research. Med Sci Sports Exerc 17:6–21
Gaesser GA, Wilson LA (1988) Effects of continuous and interval training on the parameters of the power–endurance relationship for high intensity exercise. Int J Sports Med 9:417–421
Greenhouse SW, Geisser S (1959) On methods in the analysis of profile data. Psychometrika 24:95–112
Harrison TR, Pilcher C (1930) Studies in congestive heart failure: II. The respiratory exchange during and after exercise. J Clin Invest 8:291
Heck H, Mader A, Hesse G, Mucke S, Muller R, Hollmann W (1985) Justification of the 4- mmol·l−1 lactate threshold. Int J Sports Med 6:117–130
Hill AV, Long CNH, Lupton H (1924) Muscular exercise lactic acid and the supply and utilization of oxygen: Part VII. Muscular exercise and oxygen intake. Proc R Soc Lond [B] 97:155–167
Hill DW (1993) The critical power concept. Sports Med 16:237–254
Hill DW, Smith JC, Leuschel JL, Chasteen SD, Miller SA (1995) Effect of pedal cadence on parameters of the hyperbolic power–time relationship. Int J Sports Med 16:82–87
Hollman W (1959) The relationship between pH, lactic acid, potassium in arterial and venous blood, the ventilation (PoW) and pulse frequency during increasing spirometric work in endurance-trained and untrained persons. Pan-American Congress for Sports Medicine, Chicago, Ill.
Housh DJ, Housh TJ, Bauge SM (1989) The accuracy of the critical power test for predicting time to exhaustion during cycle ergometer. Ergonomics 32:997–1004
Housh TJ, Devries HA, Housh DJ, Tichy MW, Smyth KD, Tichy A (1991) The relationship between critical power and the onset of blood lactate accumulation. J Sports Med Phys Fitness 31:31–36
Hughes E, Turner S, Brooks G (1982) Effects of glycogen depletion and pedaling speed on "anaerobic threshold". J Appl Physiol 52:1598–1607
Hughson RL, Orok CJ, Staudt LE (1984) A high velocity treadmill running test to assess endurance running potential. Int J Sports Med 5:23–25
Jenkins DG, Quigley BM (1990) Blood lactate in trained cyclists during cycle ergometry at critical power. Eur J Appl Physiol 61:278–283
Kinderman W, Simon G, Keul J (1979) The significance of the aerobic-anaerobic transition for determination of workload intensities during endurance training. Eur J Appl Physiol 42:25–34
Loat CER, Rhodes EC (1993) Relationship between lactate and ventilatory thresholds during prolonged exercise. Sports Med 15:104–115
McLellan TM, Cheung KS (1992) A comparative evaluation of the individual anaerobic threshold and the critical power. Med Sci Sports Exerc 24:543–550
McLellan TM, Skinner JS (1981) The use of the aerobic threshold as basis of training. Can J Appl Sports Sci 6:197–201
Moritani T, Nagata A, De Vries HA, Muro M (1981) Critical power as a measure of physical work capacity and anaerobic threshold. Ergonomics 24:339–350
Owles WH (1930) Alterations in the lactic acid content of the blood as a result of light exercise, and associated changes in CO2 combining power of the blood and into alveolar CO2 pressure. J Physiol (Lond) 69 214–237
Poole DC, Ward SA, Gardner GW, Whipp BJ (1988) A metabolic and respiratory profile of the upper limit for prolonged exercise in man. Ergonomics 31:1265–1279
Pringle JSM, Jones AM (2002) Maximal lactate steady state, critical power and EMG during cycling. Eur J Appl Physiol 10.1007/s00421-002-0703-4
Scherrer J, Monod H (1960) Le travail musculaire local et la fatigue chez l'homme. Masson et Compagnie, Paris
Shapiro SS, Wilk MB, Chen HJ (1968) A comparative study of various tests of normality. J Am Stat Assoc 63:1343–1372
Smith CGM, Jones AM (2001) The relationship between critical velocity, maximal lactate steady-state velocity and lactate turnpoint velocity in runners. Eur J Appl Physiol 85:19–26
Stevens J (1996) Applied multivariate statistics for the social sciences. Erlbaum, Mahwah, NJ, USA
Takaishi T, Yasuda Y, Moritani T (1994) Neuromuscular fatigue during prolonged pedaling exercise at different pedaling rates. Eur J Appl Physiol 69:154–158
Takano N (1988) Effects of pedal rates on respiratory responses to incremental bicycle work. J Physiol (Lond) 396:389–397
Vallier JM, Bigard AX, Carré F, Eclache JP, Mercier J (2000) Détermination des seuils lactiques et ventilatoires. Position de la société française de médecine du sport. Sci Sports 15:133–140
Vandewalle H, Vautier JF, Kachouri M, Lechevalier JM, Monod H (1997) Work–exhaustion time relationships and the critical power concept. J Sports Med Phys Fitness 37:89–102
Vercruyssen F, Hausswirth C, Smith D, Brisswalter J (2001) Effect of exercise duration on optimal pedalling rate choice in triathletes. Can J Appl Physiol 26:44–54
Wakayoshi K, Yoshida T, Udo M, Haradaa T, Moritani T, Mutoh Y, Miyashita M (1993) Does critical swimming velocity represent exercise intensity at maximal lactate steady state? Eur J Appl Physiol 66:90–95
Wasserman K, McIlroy MB (1964) Detecting the threshold of anaerobic metabolism in cardiac patients during exercise. Am J Cardiol 14:844–852
Wasserman K, Whipp BJ, Koyl SN, Beaver WL (1973) Anaerobic threshold and respiratory gas exchange during exercise. J Appl Physiol 35:236–243
Weissland T, Pelayo P, Vanvelcenaeher J, Marais G, Lavoie JM, Robin H (1997) Physiological effects of variations of spontaneously chosen crank rates in incremental upper body exercise. Eur J Appl Physiol 76:428–433
Weissland T, Marais G, Robin H, Vanvelcenaeher J, Pelayo P (1999) Relationship in humans between spontaneously chosen crank rate and power output during upper body exercise at different levels of intensity. Eur J Appl Physiol 79:230–236
Winter EM, Eston RG, Lamb K (2001) Statistical analysis in the physiology of exercise and kinanthropometry. J Sports Sci 19:761–775
Yamamoto Y, Mizashita M, Hughson RL, Tamura S, Shinohara M, Mutoh Y (1991) The ventilatory threshold gives maximal lactate steady state. Eur J Appl Physiol 63:55–59
Acknowledgements
The authors would like to thank all the participants and more particularly, D. Boy and Dr. P. Camblin for their enthusiastic co-operation
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Dekerle, J., Baron, B., Dupont, L. et al. Maximal lactate steady state, respiratory compensation threshold and critical power. Eur J Appl Physiol 89, 281–288 (2003). https://doi.org/10.1007/s00421-002-0786-y
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-002-0786-y