
Abstract
To examine factors that regulate ceramide production during keratinization of the human stratum corneum (SC), we developed a reconstructed human epidermal keratinization model in which a fresh layer of SC is newly formed within 1 week. Addition of the UDP-glucose: ceramide glucosyltransferase inhibitor 1-phenyl-2-decanoylamino-3-morpholino-1-propanol significantly diminished SC ceramide levels (expressed as µg/mg protein) with decreased glucosylceramide levels. Desipramine hydrochloride, an inhibitor of sphingomyelinase, also significantly reduced SC ceramide levels. Similarly, conduritol B epoxide, an inhibitor of β-glucocerebrosidase, significantly down-regulated SC ceramide levels and significantly increased glucosylceramide levels. These results indicate the reliability of this model to elucidate ceramide synthesis regulating factors. Using this model, we assessed the effects of the inflammatory cytokine interleukin-1α (IL-1α), several bioactive sphingolipids and all-trans retinoic acid (RA) on ceramide levels in the SC. Whereas treatment with IL-1α (at 10 nM) significantly down-regulated ceramide levels, treatment with sphingosylphosphorylcholine (at 50 µM) or sphingosine-1-phosphate (at 10 or 20 µM) distinctly up-regulated ceramide levels. Interestingly, RA (at low as 10 nM) significantly up-regulated ceramide levels without affecting the formation of the SC or levels of keratinization-related proteins in the epidermis. The increased levels of ceramide were accompanied by a significantly increased secretion of granulocyte–macrophage colony-stimulating factor as well as by a significantly down-regulated expression of acid-ceramidase at both the gene and protein levels. Taken together, our results underscore the superiority of this reconstructed human epidermal keratinization model to analyze factors that regulate ceramide synthesis, especially in human SC.
Similar content being viewed by others
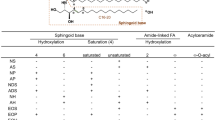


Avoid common mistakes on your manuscript.
Introduction
Ceramides in the stratum corneum (SC) play a pivotal role in the water-holding property [13–15] and in the cutaneous permeability barrier of the skin [3, 8, 18]. Dysfunction of either or both of those parameters occurs in the skin of patients with atopic dermatitis (AD) [12] or in aged skin [12] due to a ceramide deficiency. As for the mechanism involved in the ceramide deficiency in AD, we have already reported that a novel sphingolipid metabolic enzyme (termed sphingomyelin/glucosylceramide deacylase) is highly expressed in the epidermis of patients with AD. The expression of that enzyme results in the generation of lysosphingolipid, sphingosylphosphorylcholine (SPC) and glucosylsphingosine, instead of ceramides produced by the normally expressed ceramide producing enzymes, acid-sphingomyelinase (SMase) and β-glucocerebrosidase (GCase), which leads in turn to the ceramide deficiency [4, 16]. Recently, sphingomyelin/glucosylceramide deacylase has been purified from rat skin and was found to be identical to the β-subunit of acid-ceramidase (CDase) which consists of a complex of an α-subunit and a β-subunit linked by two S–S bonds (unpublished data). This strongly suggested that the induction of sphingomyelin deacylase in AD is mediated by an abnormality of the S–S cross-linking enzyme responsible for the formation of the α-subunit and β-subunit complex of CDase during its processing in the endoplasmic reticulum or is due to the cleavage of those S–S bonds by unknown enzymes. In aged skin with decreased water content, CDase activity is remarkably up-regulated in the SC [17]. However, little is known about the mechanism(s) underlying the high expression levels of sphingomyelin/glucosylceramide deacylase or CDase in several pathological conditions. To determine which biological factors elicit the expression of those abnormal enzymes, an animal model is needed in which the topical application of chemicals and/or genetically mutated mice can be used to test whether a similar ceramide deficiency is evoked in the SC. However, not all data obtained using animal models can be applied to humans because there is no proof of the similar sphingolipid metabolism between humans and other animals. It seems reasonable to assume that a human epidermal model consisting of human keratinocytes alone would be the most suitable model by which bioactive factors capable of modulating ceramide levels in the SC could be evaluated to analyze the mechanisms involved in the ceramide deficiency.
Although human epidermal models consisting of human keratinocytes alone are commercially available and have been used in several studies to analyze sphingolipid metabolism in the epidermis, almost all of those studies focused on ceramide levels in the entire epidermis [25, 29]. In this study, we developed a new reconstructed human epidermal keratinization culture model in which epidermis without the SC (thus prior to keratinization) is cultured for 1 week to form a complete SC layer after which ceramides can be extracted from the whole SC and quantified as per SC protein. Characterization of the new model revealed that several specific sphingolipid metabolic inhibitors induce a ceramide deficiency in the SC in a fashion similar to the in vivo situation in human skin. Further, the results show that several biologically active factors, such as interleukin-1α (IL-1α), SPC, sphingosine-1-phosphate (S1P) or all-trans retinoic acid (RA) serve as suppressors or stimulators of ceramide synthesis in the SC. These findings underscore the superiority of this new reconstructed human epidermal keratinization model to analyze factors that regulate ceramide synthesis, especially in the SC of human skin.
Materials and methods
Reconstructed human epidermal keratinization culture model
The reconstructed human epidermal keratinization model was established by Green’s method (LabCyte EPI-MODEL, Japan Tissue Engineering Co., Ltd.) [2, 24] In this model, foreskin-derived human keratinocytes are incubated in an air–liquid condition for a total of 14 days during which the SC formation occurs in the latter 7 days. The model was shipped to us after the first 7 days and then we incubated it at 37 °C in a 5 % CO2 atmosphere for the latter 7 days in medium containing the following chemical samples: 1-phenyl-2-decanoylamino-3-morpholino-1-propanol (PDMP, Sigma-Aldrich), conduritol B epoxide (CBE, Matreya), desipramine hydrochloride (DSP, Sigma-Aldrich), SPC (Sigma-Aldrich), S1P (Sigma-Aldrich), IL-1α (R&D Systems), RA (Wako) or solvents [dimethylsulfoxide or phosphate-buffered saline (PBS)] as a control. The basic medium for the latter 7 days also contains ascorbic acid (concentration was not available due to manufacturer’s confidential information) for enhancement of SC formation. All media were replaced every day.
Lipid extraction
After incubation, the whole skin was peeled from the membrane and the epidermal cells were separated from the SC by incubation in a trypsin (2.5 mg/mL)/EDTA (0.2 mg/mL) aqueous solution at 37 °C for 1 h. The separation was confirmed by microscopic observation. Both the SC and the epidermal cell samples were washed with PBS and were stored at −80 °C until used. Lipids were extracted using a modification of the Bligh/Dyer method [1] as follows: Samples were homogenized in Bligh/Dyer solution (chloroform:methanol:PBS = 1:2:0.8 by vol) and were mixed for 20 min. Following centrifugation at 3,000 rpm for 15 min, the supernatants were collected in glass test tubes and then 1 mL of chloroform and 1 mL of PBS were added. After 20 min of mixing, and centrifugation for 15 min at 3,000 rpm, the bottom layer was collected and dried at 30 °C under nitrogen gas. The remaining precipitates were collected for quantification of total protein amount, using BCA protein assay kit (Thermo Scientific).
Thin layer chromatography
The amounts of ceramides and glucosylceramides were determined by high-performance thin-layer chromatography (HPTLC) as described by Imokawa et al. [12]. The dried samples were dissolved in chloroform/methanol = 2/1 solvent and were fractionated by TLC on 20 × 10 cm silica gel plates (Merck). Ceramides were developed twice over 9 cm with chloroform/methanol/acetic acid (190:9:1 by vol) followed by separation over 1 cm with chloroform/methanol/acetone (76:20:4 by vol) to develop glucosylceramides. The plates were scanned and analyzed using Quantity One 1-D analysis software (Bio-Rad). Species of the ceramides were assigned according to Motta nomenclature as well as Masukawa ceramide characterization [19–22].
Real-time RT-PCR
The expression of mRNAs for several sphingolipid enzymes were examined by real-time RT-PCR as described elsewhere [26]. The primers used for real-time PCR were synthesized by Invitrogen and the sequences are as follows: 5′-TGGCTCTATGAAGCGATGG-3′ as the forward and 5′-AGGCCGATGTAGGTAGTTGC-3′ as the reverse primers for SMase; 5′-CCTTCTTCCTTGATGATCGC-3′ as the forward and 5′-GTGGAGTTCACCATGGTTCG-3′ as the reverse primers for CDase; 5′-TGGCATTGCTGTACATTGG-3′ as the forward and 5′-CGTTCTTCTGACTGGCAACC-3′ as the reverse primers for GCase; 5′-GCGCGCTACTTGGAGAAAGA-3′ as the forward and 5′-TGTTCCACCGTGACCACAAC-3′ as the reverse primers for serine palmitoyltransferase (SPT1); 5′-AGCCGCCAAAGTCCTTGAG-3′ as the forward and 5′-CTTGTCCAGGTTTCCAATTTCC-3′ as the reverse primers for SPT2; 5′-GAAGGTGAAGGTCGGAGTCAACG-3′ as the forward and 5′-AGTCCTTCCACGATAACCAAAGTTG-3′ as the reverse primers for GAPDH.
Western blotting
Reconstructed human epidermal keratinization model tissues were lysed and subjected to western blotting as described elsewhere [26]. The following primary antibodies were used: rabbit anti-human ASAH1 polyclonal antibody (Proteintech group) for CDase, anti-SMase rabbit polyclonal antibody (Exalpha Biologicals) for SMase, and monoclonal anti-beta-actin clone ac-74 mouse ascites fluid (Sigma) for β-actin.
ELISA
The amounts of cytokines or chemokines after 1 or 2 days of culture were determined with an ELISA kit (Thermo Scientific) according to the manufacturer’s protocol.
Statistical analysis
All data are expressed as mean ± SD (n = 3) unless noted otherwise. For pairwise comparisons, either the Student’s or Welch’s t test was applied. For multiple comparisons, data were tested by one-way ANOVA, subsequently using the Tukey multiple comparison test. p values <0.05 are considered statistically significant.
Results
H&E staining of the separated SC and the whole epidermis
After the epidermal model was incubated for 1 h with the trypsin/EDTA solution, the SC layers were separated from the whole epidermis. H&E staining of the separated SC and the whole epidermis indicates that the SC layers were completely separated without contamination by epidermal cells.
Effect of the inhibitor for UDP-glucose: ceramide glucosyltransferase, PDMP, on ceramide levels and composition in the SC
The addition of PDMP at a concentration of 50 µM during 7 days of culture induced a slight thinning of the epidermis (Fig. 1a) and resulted in a marked decrease in all species of ceramides and glucosylceramide as revealed by TLC analysis (Fig. 1b). Quantitative analysis of ceramides revealed that ceramide levels expressed per well or per mg protein were significantly down-regulated by PDMP (Fig. 1c, e) despite the fact that the SC protein levels expressed per well were significantly decreased (Fig. 1d). The composition of ceramide species (Fig. 1f) demonstrated that almost all ceramide species (except for Cer[AP]) were significantly reduced by PDMP treatment. Quantitative analysis of glucosylceramide levels also demonstrated a significant reduction of glucosylceramide (expressed per well or per mg protein) in the SC (Fig. 1g, h).

Effects of PDMP on ceramide levels in the SC. a H&E staining on days 0 or 7, b TLC analysis, c effects on ceramide level per well, d effects on protein level of the SC per well, e effects on total ceramide per mg SC protein, f effects on ceramide species per mg SC protein, g effects on glucosylceramide per well, h effects on glucosylceramide per mg protein. n = 3
Effect of the inhibitor for GCase, CBE, on ceramide levels and composition in the SC and within epidermal cells
The addition of CBE at a concentration of 3 mM during 7 days of culture induced no structural changes in the epidermis (Fig. 2a) but resulted in a marked decrease in all species of ceramides and in a remarkable up-regulation of glucosylceramide as revealed by TLC analysis (Fig. 2b). Quantitative analysis of ceramides revealed that ceramide levels expressed per well or per mg protein were significantly down-regulated by CBE (Fig. 2c, e) while SC protein levels expressed per well remained unchanged (Fig. 2d). The composition of ceramide species (Fig. 2f) demonstrated that all ceramide species were significantly reduced by the CBE treatment. In contrast, quantitative analysis of glucosylceramide demonstrated a significant and remarkable up-regulation of glucosylceramide (expressed per well or per mg protein) in the SC (Fig. 2g, h).

Effects of CBE on ceramide levels in the SC. a H&E staining on days 0 or 7, b TLC analysis, c effects on ceramide level per well, d effects on protein level per well, e effects on total ceramide per mg SC protein, f effects on ceramide species per mg SC protein, g effects on glucosylceramide per well, h effects on glucosylceramide per mg protein. n = 3
Effect of the inhibitor for SMase, DSP, on ceramide levels and composition in the SC
The addition of DSP at a concentration of 25 µM during 7 days of culture induced a slight thinning of the epidermis (Fig. 3a) and resulted in a distinct decrease in all species of ceramides as revealed by TLC analysis (Fig. 3b). Quantitative analysis of ceramides revealed that ceramide levels expressed per well or per mg protein were significantly down-regulated by DSP (Fig. 3c, e) despite the fact that the SC protein levels expressed per well were slightly but not significantly decreased (Fig. 3d). The composition of ceramide species (Fig. 3f) demonstrated that ceramide species, such as Cer[EOS], [NDS/NS], [NP] and [AH], but not [EOH], [AS], [NH] and [AP], were significantly reduced by DSP treatment although Cer[EOS] decreased to a lesser extent than did Cer[NDS/NS] and [NP].

Effects of DSP on ceramide levels in the SC. a H&E staining at days 0 or 7, b TLC analysis, c effects on ceramide level per well, d effects on protein level of the SC per well, e effects on total ceramide per mg protein, f effects on ceramide species per mg protein
Comparison of ceramide levels between the SC and the epidermal cells
Comparison of ceramide levels expressed per mg protein between the SC and the epidermal cells in inhibitor-treated reconstructed epidermal keratinization models demonstrated that while treatment with CBE at relatively high concentrations significantly reduced SC ceramide levels (as a result of decreased ceramide and no change in protein), the same treatment slightly diminished epidermal ceramides (as a result of decreased ceramide and no change in protein) (Table 1). On the other hand, treatment with CBE at relatively low concentrations elicited no change in ceramide levels in the SC (as a result of decreased ceramide and protein), but slightly up-regulated ceramide levels (as a result of an increase in ceramide and no change in protein) within epidermal cells (Table 2). Following treatment with DSP, whereas the ceramide level was significantly down-regulated in the SC (as a result of decreased ceramide and no change in protein), there was a significantly up-regulated ceramide level (as a result of increased ceramide and protein) in the epidermal cells (Table 3). These comparisons indicate that there are distinctly different responses in ceramide metabolism between the SC and epidermal cells and that ceramide levels vary with a balance between ceramide and protein levels. This strongly suggests the importance for measuring ceramide levels only in the SC.
Effects of several biologically active factors on ceramide levels and composition in the SC
To highlight the influence of inflammatory cytokines, we used IL-1α to assess its effect on SC ceramide levels. The addition of IL-1α at a concentration of 10 nM during 7 days of culture induced no distinct change in the keratinization process of the epidermal equivalents (Fig. 4a). Quantitative analysis of ceramides revealed that ceramide levels expressed per mg protein were significantly down-regulated by IL-1α treatment at 10 nM (Fig. 4d) while ceramide levels (Fig. 4b) or SC protein (Fig. 4c) expressed per well were significantly decreased or not changed, respectively. The composition of ceramide species (Fig. 4e) demonstrated that Cer[EOS], [NP], [EOH], [NH] and [AH] species expressed per mg protein were significantly decreased by treatment with IL-1α, whereas only Cer[NDS/NS] was significantly increased. Second, we examined the effects of biologically active sphingolipids such as SPC and S1P. The addition of SPC or S1P at concentrations of 5–50 or 5–20 µM, respectively, during 7 days of culture induced no distinct change in the keratinization process of the epidermal equivalents (Figs. 5a, 6a). Quantitative analysis of ceramides revealed that ceramide levels expressed per mg protein were significantly up-regulated by SPC or S1P at 50 or 10–20 µM, respectively (Figs. 5d, 6d), while ceramide levels (Figs. 5b, 6b) or SC protein (Figs. 5c, 6c) expressed per well were significantly increased at 50 or 10–20 µM, or were not changed, respectively. The composition of ceramide species (Figs. 5e, 6e) demonstrated that Cer[NDS/NS], [NP], [EOH] and [AH]expressed per mg protein were significantly increased by SPC at 50, 50, 5–50 and 5–50 µM, respectively, and Cer[EOS], [NDS/NS], [NP], [EOH] and [AH] expressed per mg protein were significantly increased by S1P at 20, 5–20, 10–20, 10–20 and 20 µM, respectively. The addition of RA at concentrations of 5 or 10 nM during 7 days of culture induced no structural changes in the epidermis (Fig. 7a), whereas RA at more than 0.1 µM markedly attenuated the SC formation (data not shown); thus, we did not examine ceramide levels at higher concentrations of RA.

Effects of IL-1α on ceramide levels in the SC. a H&E staining at day 7, b effects on ceramide level per well, c effects on protein level of the SC per well, d effects on total ceramide per mg protein, e effects on ceramide species per mg protein. n = 3 or 4

Effects of SPC on ceramide levels in the SC. a H&E staining at day 7, b effects on ceramide level per well, c effects on protein level of the SC per well, d effects on total ceramide per mg protein, e effects on ceramide species per mg protein. n = 3

Effects of S1P on ceramide levels in the SC. a H&E staining on day 7, b effects on ceramide level per well, c effects on protein level of the SC per well, d effects on total ceramide per mg protein, e effects on ceramide species per mg protein

Effects of RA at low doses on SC ceramide. a H&E staining on day 7, b effects on ceramide levels per well, c effects on protein levels of the SC, d effects on ceramide per mg protein, e effects on ceramide species per mg protein. **p < 0.01, *p < 0.05
Quantitative analysis of ceramides revealed that ceramide levels expressed per well or per mg protein were significantly up-regulated by RA treatment at 10 nM (Fig. 7b, d) despite the fact that SC protein levels expressed per well remained unchanged (Fig. 7c). The composition of ceramide species (Fig. 7e) demonstrated that Cer[NP] and [AH] at 5 nM RA and Cer[NDS/NS], [NP] and [AH] at 10 nM RA were significantly increased per mg protein.
Effect of RA at lower concentrations on the expression of sphingolipid metabolic enzymes
Because RA can exert its stimulatory effect on SC ceramide production at concentrations as low as 10 nM (1,000-fold lower than other biologically active substances tested) it was of interest to elucidate its influence on the sphingolipid metabolism responsible for ceramide synthesis. Real-time RT-PCR analysis revealed that treatment with RA at concentrations of 5 or 10 nM significantly down-regulated the expression of genes encoding SMase (Fig. 8a) and CDase (Fig. 8b) on day 5 of culture, whereas they elicited no changes in the gene expression for SPT1 (Fig. 8c) or SPT2 (Fig. 8d) or in GCase (Fig. 8e) at day 5 of culture. Western blotting analysis revealed that treatment with RA significantly down-regulated protein levels of CDase (at RA concentrations of 5 and 10 nM) and SMase (at RA concentrations of 10 nM) (Fig. 8f, g).

Effects of RA at low doses on ceramide related enzymes. a–e Effects on mRNA levels of SMase, CDase, SPT1, SPT2, GCase, respectively. n = 4 or 5. f, g Effects on protein expression of CDase and SMase, respectively. Immunoblots from three independent experiments are shown. n = 3. **p < 0.01, *p < 0.05
Effect of RA on the release of cytokines or a chemokine
When levels of cytokines or chemokines, including IL-1α, TNFα, IL-4, IL-6, IL-8 and GM-CSF, were measured in the conditioned medium of the reconstructed epidermal keratinization model on days 1 and 2 of culture, treatment with RA at a concentration of 10 nM significantly stimulated the secretion of GM-CSF alone among the cytokines tested on day 1 of culture (Table 4). The same treatment on day 2 (fresh medium containing 10 nM RA was exchanged each day) significantly increased the secretion of GM-CSF to a lesser extent than on day 1 (Table 4).
Effect of RA at a low concentration on the protein expression of keratinization-related proteins
Immunohistochemistry with antibodies to filaggrin, involucrin, loricrin, transglutaminase I, transglutaminase III, cytokeratin 10 and cytokeratin 19, revealed no distinct changes in staining patterns or intensities in the RA (10 nM)-treated reconstructed epidermal keratinization model (data not shown), which suggests that 10 nM RA does not affect keratinization processing to a detectable extent.
Discussion
In this study, we developed a reconstructed human epidermal keratinization culture model, in which a newly layered SC is generated within 1 week during which factors can be added to examine their effects on ceramide levels, especially in the SC. In other studies which used similar epidermal equivalents [25, 30] as well as epidermal sheets in culture, it is difficult to assess the effects of chemicals or cytokines on ceramide levels in the SC since the entire SC is already formed before the start of culture and normal desquamation of the SC layers never occurs in a pattern similar normal epidermis in vivo due to the wet conditions. Because of these constraints, all evaluations conducted on ceramide changes had been limited not to the SC, but to the whole epidermis [5, 29]. However, although ceramide levels in the whole epidermis (which includes the SC) might roughly reflect levels in the SC since ceramides are predominantly located in abundance in the SC, one could not rule out the possibility that intracellular ceramide levels within living epidermal cells affect the total ceramide levels expressed per mg protein (due to a unbalanced change in ceramide and/or protein). This could result in errors in estimating ceramide levels in the SC which are essentially and precisely responsible for the barrier and water holding functions of the skin [3, 8, 13–15, 18]. In fact, in this study, the comparison of ceramide levels between the SC and the epidermal cells after treatment with ceramide metabolic inhibitors revealed that there is no mutual correlation in ceramide levels between the SC and the living epidermal cells. This is quite reasonable when one considers the ceramide synthetic pathway in the SC where lamellar granules are secreted into intercellular spaces between SC cells during which the ceramide precursors, glucosylceramide and sphingomyelin, are hydrolyzed by GCase and SMase, respectively, indicating a major contribution of the SC ceramide levels to such ceramide metabolic enzyme activity.
For those reasons, in this study we examined the effects of inhibitors of major sphingolipid metabolic enzymes on the formation of ceramide, especially in the SC. The addition of the UDP-glucose: ceramide glucosyltransferase inhibitor, PDMP [28, 31] at a concentration of 50 µM during the 7-day keratinization phase significantly decreased ceramide levels expressed per mg SC protein, with almost all ceramide species being deficient and this was concomitant with significantly decreased levels of glucosylceramide. This is in good agreement with the findings of our previous study [27], which used cultured murine epidermal sheets, and supports the notion that all ceramide species including acylceramide can be synthesized via glucosylceramide, as an intermediate of ceramide synthesis. This notion was also substantiated by our related study using an inhibitor of GCase [27], CBE, in which all ceramide species were significantly down-regulated in concert with a significant increase in glucosylceramide. This was also in a good agreement with our previous study [27] that used murine skin. Similarly, we show for the first time that an inhibitor of SMase, DSP [11], at a concentration of 25 µM significantly reduced the levels of ceramides, including the acylceramide Cer[EOS], without any change in the level of another acylceramide Cer[EOH] although it remains unclear why Cer[EOS] was significantly reduced by the inhibition of SMase. Among the inhibitors tested, while DSP has the lowest potential to down-regulate SC ceramide levels, CBE is the most effective in reducing SC ceramide levels, which indicates a consistency for the participation ratio [9, 30] of SMase (approximately 30 %) and GCase (approximately 70 %) for ceramide synthesis in the SC. The sum of these results indicates that the major metabolic pathways responsible for ceramide synthesis through the interface between the SC and the granular layer occur in the reconstructed human epidermal keratinization model such as occurs in vivo. That evidence allows us to set a unique keratinization model against which to evaluate the influence of several bioactive substances on the SC ceramide production during the initial keratinization process instead of using animal or human skin in vivo.
We used SPC, S1P, IL-1α and RA as biologically active substances to measure their effects on SC ceramide production because SPC [7], S1P [32], IL-1α [34] and RA [23] are known to alter the keratinization process. Of substances capable of stimulating transglutaminase expression, we found that SPC and S1P significantly increase SC ceramide levels, whereas IL-1α exerts an inhibitory effect on SC ceramide production. This suggests that the up-regulated expression of transglutaminase is not directly linked to sphingolipid metabolism which is responsible for SC ceramide levels. Interestingly, SPC up-regulated mainly Cer[NDS/NS] and [NP] which are synthesized via both major enzymes, SMase and GCase, whereas S1P up-regulated not only Cer[NDS/NS] and [NP], but also Cer[EOS], which is synthesized only via GCase pathway. Since SPC can be generated instead of ceramides from sphingomyelin via sphingomyelin deacylase pathway [6], the accumulation of SPC may lead to suppression of sphingomyelin deacylase, resulting in the increase of ceramides, although it is too speculative because it remains unclear whether sphingomyelin deacylase activity is involved in our reconstructed epidermal keratinization model. On the other hand, as S1P is one of the metabolites of ceramide degradation, the S1P accumulation may result in the increase of ceramides via the interruption of the CDase pathway [33] although we have no available data to backup these assumptions at this point.
Intriguingly, we found that RA at concentrations as low as 10 nM significantly up-regulates ceramide levels accompanied by increases in Cer[NDS/NS] and [NP] without affecting the formation of the SC (namely no changes in SC protein), whereas RA at concentrations higher than 0.1 µM remarkably attenuated formation of the SC (data not shown); thus we did not examine ceramide levels at higher RA concentrations. In contrast to the other factors tested, such a remarkably low concentration of RA that is sufficient to stimulate SC ceramide production prompted us to analyze its biological mechanism of action in more detail. The up-regulation of ceramide synthesis in the SC by low levels of RA was not associated with any marked alteration in keratinization markers, but was accompanied by the enhanced release of the Th1 type of cytokine GM-CSF. It was also accompanied by the significantly down-regulated gene expression of both SMase and CDase, whereas the gene expression levels of SPT 1/2 and GCase were not affected. Consistently, in the low RA-treated reconstructed human epidermal keratinization model, the protein expression levels of both SMase and CDase were distinctly down-regulated compared with the control non-treated epidermal model. In a related study using the same reconstructed human epidermal keratinization model, we found that the addition of GM-CSF at a concentration of 10 nM significantly stimulated ceramide levels in the SC (unpublished data). This supports the possibility that GM-CSF is one of the stimulators for the low RA-induced up-regulation of ceramide synthesis in the SC. Although the reduced expression of SMase may lead to a controversial explanation for the increased ceramide levels in the low RA-treated epidermal model, we hypothesize that the significant down-regulation of CDase expression and the minor increase in GCase expression as well as a weaker contribution of the SMase pathway to produce all ceramides could compensate for the decrease in SMase expression, resulting in the slight increase of ceramide levels in the SC. Further research into the mechanism underlying the changes in expression of sphingolipid metabolic enzymes is beyond the scope of this study which focused on establishing the feasibility of the epidermal keratinization model using human epidermal equivalents.
In conclusion, we observed that several specific sphingolipid metabolic inhibitors induce a ceramide deficiency in the SC in a fashion similar to the in vivo situation [10] and that several biologically active factors, such as IL-1α, SPC, S1P and RA, serve as suppressors or stimulators of ceramide synthesis in the SC. We also found that the increased production of SC ceramide by RA is mainly mediated by the down-regulated expression of CDase. These results underscore the superiority of the reconstructed human epidermal keratinization model to examine factors that regulate ceramide synthesis, especially in the human SC as well as for analyzing their modes of action.
Abbreviations
- AD:
-
Atopic dermatitis
- CBE:
-
Conduritol B epoxide
- CDase:
-
Acid-ceramidase
- DSP:
-
Desipramine hydrochloride
- GCase:
-
β-Glucocerebrosidase
- GM-CSF:
-
Granulocyte–macrophage colony-stimulating factor
- HPTLC:
-
High-performance thin layer chromatography
- IL-1α:
-
Interleukin-1α
- PDMP:
-
1-Phenyl-2-decanoylamino-3-morpholino-1-propanol
- RA:
-
all-trans retinoic acid
- SC:
-
Stratum corneum
- S1P:
-
Sphingosine-1-phosphate
- SMase:
-
Acid-sphingomyelinase
- SPC:
-
Sphingosylphosphorylcholine
- SPT:
-
Serine palmitoyltransferase
References
Bligh EG, Dyer WJ (1959) A rapid method of total lipid extraction and purification. Can J Biochem Physiol 37:911–917
Green H (1978) Cyclic AMP in relation to proliferation of the epidermal cell: new view. Cell 15:801–811
Grubauer G, Feingold KR, Harris RM, Elias PM (1989) Lipid content and lipid type as determinants of the epidermal permeability barrier. J Lipid Res 30:89–96
Hara J, Higuchi K, Okamoto R, Kawashima M, Imokawa G (2000) High-expression of sphingomyelin deacylase is an important determinant of ceramide deficiency leading to barrier disruption in atopic dermatitis. J Invest Dermatol 115:406–413
Hatano Y, Terashi H, Arakawa S, Katagiri K (2005) Interleukin-4 suppresses the enhancement of ceramide synthesis and cutaneous permeability barrier functions induced by tumor necrosis factor-alpha and interferon-gamma in human epidermis. J Invest Dermatol 124:786–792
Higuchi K, Hara J, Okamoto R, Kawashima M, Imokawa G (2000) The skin of atopic dermatitis patients contains a novel enzyme, glucosylceramide sphingomyelin deacylase, which cleaves the N-acyl linkage of sphingomyelin and glucosylceramide. Biochem J 350:747–756
Higuchi K, Kawashima M, Takagi Y, Ichikawa Y, Imokawa G (2001) Sphingosylphosphorylcholine is an activator of transglutaminase activity in human keratinocytes. J Lipid Res 42:1562–1570
Holleran WM, Feingold KR, Man MQ, Gao WN, Lee JM, Elias PM (1991) Regulation of epidermal sphingolipid synthesis by permeability barrier function. J Lipid Res 32:1151–1158
Holleran WM, Ginns EI, Menon GK, Grundmann JU, Fartasch M, McKinney CE, Elias PM, Sidransky E (1994) Consequences of beta-glucocerebrosidase deficiency in epidermis. Ultrastructure and permeability barrier alterations in Gaucher disease. J Clin Invest 93:1756–1764
Holleran WM, Takagi Y, Menon GK, Legler G, Feingold KR, Elias PM (1993) Processing of epidermal glucosylceramides is required for optimal mammalian cutaneous permeability barrier function. J Clin Invest 91(4):1656–1664
Hurwitz R, Ferlinz K, Sandhoff K (1994) The tricyclic antidepressant desipramine causes proteolytic degradation of lysosomal sphingomyelinase in human fibroblasts. Biol Chem Hoppe Seyler 375:447–450
Imokawa G, Abe A, Jin K, Higaki Y, Kawashima M, Hidano A (1991) Decreased level of ceramides in stratum corneum of atopic dermatitis: an etiologic factor in atopic dry skin? J Invest Dermatol 96:523–526
Imokawa G, Akasaki S, Hattori M, Yoshizuka N (1986) Selective recovery of deranged water-holding properties by stratum corneum lipids. J Invest Dermatol 87:758–761
Imokawa G, Akasaki S, Minematsu Y, Kawai M (1989) Importance of intercellular lipids in water-retention properties of the stratum corneum: induction and recovery study of surfactant dry skin. Arch Dermatol Res 281:45–51
Imokawa G, Hattori M (1985) A possible function of structural lipids in the water-holding properties of the stratum corneum. J Invest Dermatol 84:282–284
Ishibashi M, Arikawa J, Okamoto R, Kawashima M, Takagi Y, Ohguchi K, Imokawa G (2003) Abnormal expression of the novel epidermal enzyme, glucosylceramide deacylase, and the accumulation of its enzymatic reaction product, glucosylsphingosine, in the skin of patients with atopic dermatitis. Lab Invest 83:397–408
Jin K, Higaki Y, Takagi Y, Higuchi K, Yada Y, Kawashima M, Imokawa G (1994) Analysis of beta-glucocerebrosidase and ceramidase activities in atopic and aged dry skin. Acta Derm Venereol 74:337–340
Lampe MA, Burlingame AL, Whitney J, Williams ML, Brown BE, Roitman E, Elias PM (1983) Human stratum corneum lipids: characterization and regional variations. J Lipid Res 24:120–130
Madison KC (2003) Barrier function of the skin: “la raison d’être” of the epidermis. J Invest Dermatol 121:231–241
Masukawa Y, Narita H, Sato H, Naoe A, Kondo N, Sugai Y, Oba T, Homma R, Ishikawa J, Takagi Y, Kitahara T (2009) Comprehensive quantification of ceramide species in human stratum corneum. J Lipid Res 50:1708–1719
Masukawa Y, Narita H, Shimizu E, Kondo N, Sugai Y, Oba T, Homma R, Ishikawa J, Takagi Y, Kitahara T, Takema Y, Kita K (2008) Characterization of overall ceramide species in human stratum corneum. J Lipid Res 49:1466–1476
Motta S, Monti M, Sesana S, Caputo R, Carelli S, Ghidoni R (1993) Ceramide composition of the psoriatic scale. Biochim Biophys Acta 1182(2):147–151
Nagpal S, Thacher SM, Patel S, Friant S, Malhotra M, Shafer J, Krasinski G, Asano AT, Teng M, Duvic M, Chandraratna RA (1996) Negative regulation of two hyperproliferative keratinocyte differentiation markers by a retinoic acid receptor-specific retinoid: insight into the mechanism of retinoid action in psoriasis. Cell Growth Differ 7:1783–1791
Rheinwald JG, Green H (1975) Serial cultivation of strains of human epidermal keratinocytes: the formation of keratinizing colonies from single cells. Cell 6:331–344
Rivier M, Castiel I, Safonova I, Ailhaud G, Michel S (2000) Peroxisome proliferator-activated receptor-alpha enhances lipid metabolism in a skin equivalent model. J Invest Dermatol 114:681–687
Suganuma K, Nakajima H, Otsuki M, Imokawa G (2010) Astaxanthin attenuates the UVA-induced up-regulation of matrix-metalloproteinase-1 and skin fibroblast elastase in human dermal fibroblasts. J Dermatol Sci 58:136–142
Takagi Y, Nakagawa H, Matsuo N, Nomura T, Takizawa M, Imokawa G (2004) Biosynthesis of acylceramide in murine epidermis: characterization by inhibition of glucosylation and deglucosylation, and by substrate specificity. J Invest Dermatol 122:722–729
Takami Y, Abe A, Matsuda T, Shayman JA, Radin NS, Walter RJ (1998) Effect of an inhibitor of glucosylceramide synthesis on cultured human keratinocytes. J Dermatol 25:73–77
Tokudome Y, Uchida R, Yokote T, Todo H, Hada N, Kon T, Yasuda J, Hayashi H, Hashimoto F, Sugibayashi K (2010) Effect of topically applied sphingomyelin-based liposomes on the ceramide level in a three-dimensional cultured human skin model. J Liposome Res 20:49–54
Uchida Y, Hara M, Nishio H, Sidransky E, Inoue S, Otsuka F, Suzuki A, Elias PM, Holleran WM, Hamanaka S (2000) Epidermal sphingomyelins are precursors for selected stratum corneum ceramides. J Lipid Res 41:2071–2082
Uemura K, Sugiyama E, Tamai C, Hara A, Taketomi T, Radin NS (1990) Effect of an inhibitor of glucosylceramide synthesis on cultured rabbit skin fibroblasts. J Biochem 108:525–530
Vogler R, Sauer B, Kim DS, Schäfer-Korting M, Kleuser B (2003) Sphingosine-1-phosphate and its potentially paradoxical effects on critical parameters of cutaneous wound healing. J Invest Dermatol 120:693–700
Yada Y, Higuchi K, Imokawa G (1995) Purification and biochemical characterization of membrane-bound epidermal ceramidase from guinea pig skin. J Biol Chem 270:12677–12684
Yano S, Banno T, Walsh R, Blumenberg M (2008) Transcriptional responses of human epidermal keratinocytes to cytokine interleukin-1. J Cell Physiol 214:1–13
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Yoshida, N., Sawada, E. & Imokawa, G. A reconstructed human epidermal keratinization culture model to characterize ceramide metabolism in the stratum corneum. Arch Dermatol Res 304, 563–577 (2012). https://doi.org/10.1007/s00403-012-1232-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00403-012-1232-6