
Abstract
We reclassified the pathological subtypes of dementia with Lewy bodies (DLB), based on both Lewy pathology and Alzheimer pathology, to clarify the pathological entity of DLB and the boundary between DLB and Alzheimer’s disease (AD) in autopsied cases, using both pathological and immunohistochemical methods. DLB was classified as either limbic type or neocortical type according to the degree of Lewy pathology including Lewy bodies (LB) and LB-related neurites by our staging, and was classified as pure form, common form or AD form according to the degree of Alzheimer pathology including neurofibrillary tangles (NFT) and amyloid deposits by Braak staging. These combined subtypes were lined up on a spectrum, not only with Lewy pathology but also with other DLB-related pathologies including Alzheimer pathology, neuronal loss in the substantia nigra, spongiform change in the transentorhinal cortex and LB-related neurites in the CA2–3 region. In contrast, the Lewy pathology of AD did not meet the stages of Lewy pathology in DLB, and there were scarcely any similarities in other DLB-related pathologies between AD and DLB. In addition, the Lewy pathology of AD had characteristics different from that of DLB, including the coexistence rate of LB with NFT, and the immunohistochemical and immunoelectron microscopic findings of LB and LB-related neurites. These findings suggest that DLB is a distinctive pathological entity that can be differentiated from AD, although it shows some pathological subtypes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Dementia with Lewy bodies (DLB) is the second most frequent neurodegenerative dementing disorder after Alzheimer’s disease (AD) [24]. DLB is clinically characterized by progressive dementia, which is frequently accompanied by parkinsonism and psychiatric symptoms, and is pathologically characterized by the occurrence of Lewy bodies (LB) in the brain [18, 24]. The LB are morphologically divided into brainstem type and cortical type. Brainstem-type LB are found in the brainstem nuclei and diencephalons, while cortical-type LB are preferentially found in the cerebral limbic cortices and amygdala [17]. Although the occurrence of LB is the only feature essential for the pathological diagnosis of DLB, LB-related neurites in the CA2–3 region of the hippocampus, Alzheimer pathology in the cerebral cortex, neuronal loss in the brainstem nuclei, and spongiform change in the transentorhinal cortex are also frequently seen in DLB [24].
In the Consensus guidelines for pathological diagnosis of DLB, DLB has been classified into three pathological subtypes including the brainstem-predominant type, limbic type and neocortical type, based on the distribution of LB [24]. The pathological entity of DLB is, however, not sufficiently established, because the criteria for the classification of these subtypes are not practical for Lewy pathology, the boundary between DLB and Parkinson’s disease (PD) is unclear, and DLB is accompanied by Alzheimer pathology in various degrees. It has been recently reported that AD cases, including familial and sporadic cases, occasionally show LB in the limbic region, especially in the amygdala [1, 8, 21, 22]. These findings also make the pathological boundary between DLB and AD indistinct.
It is sometimes difficult to differentiate cortical-type LB from other neuronal structures, such as neurofibrillary tangles (NFT) and ballooned neurons, using hematoxylin-eosin (HE) staining. Anti-ubiquitin antibodies sensitively detect LB, but are not specific for LB [20]. Recently, α-synuclein has been recognized as a major component of LB, and anti-α-synuclein antibodies exclusively detect LB in PD and DLB brains [3, 28].
We have previously proposed new pathological subtypes for DLB, using autopsied cases [14]. Furthermore, we prepared the staging to evaluate the degree of Lewy pathology including LB and LB-related neurites in the cerebrum [23]. LB and LB-related neurites were defined as α-synuclein-positive structures in the neuronal cell body, and α-synuclein-positive structures in the dendrites and axons, respectively.
In the present study, we reclassified the pathological subtypes of DLB, according to our staging of Lewy pathology and Braak staging of Alzheimer pathology [4] to clarify the pathological entity of DLB, and showed that DLB could be pathologically differentiated from AD.
Materials and methods
We examined 51 autopsied cases (mean age 76.7 years; mean brain weight 1,102 g) preserved in our laboratory. All the cases had shown progressive dementia without a family history of dementia, and 31 out of 51 cases had shown parkinsonism after or before dementia. All the cases pathologically demonstrated some Lewy pathology at least in the amygdala, and also some Alzheimer pathology at least in the limbic region.
The brains were fixed in 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). Two coronal slices of the cerebral hemispheres through the levels of the mamillary body and lateral geniculate body, and two horizontal slices of the midbrain through the red nucleus and the pons through the locus ceruleus were embedded in paraffin. The 6-μm-thick sections were stained using the HE, Klüver-Barrera (KB), methenamine silver (MS) and Gallyas-Braak (GB) methods for pathological examination. The serial sections were also immunostained with anti-α-synuclein (SyαC: polyclonal, rabbit, 1:200) [23], anti-PHF tau (AT8: monoclonal, mouse, 1:2,000, Innogenetics, Belgium) and anti-Aβ (polyclonal, rabbit, 1:5,000, donated by Dr. T. Ishii) antibodies. Immunolabeling was detected using the avidin-biotin-horseradish peroxidase complex (ABC) method (Elite Kit: Vector, USA), and visualized with diaminobenzidine (DAB: Wako, Japan). In addition, the sections were double-immunostained with SyαC and AT8. Immunolabeling was detected using the ABC method and the ABC-alkaline phosphatase method (ABC-AP Kit: Vector), and visualized with DAB and fast blue, respectively. Before immunostaining, these sections were pretreated with 70% formic acid for 10 min for SyαC and anti-Aβ antibody.
Lewy pathology including LB and LB-related neurites was evaluated with SyαC immunostaining, and Alzheimer pathology including NFT and amyloid deposits was evaluated with AT8 and Aβ immunostaining, respectively. The number of SyαC-positive LB was counted on microscopic fields (field size: 0.66 mm2) at ×200 magnification in the amygdala, the limbic cortex (the transentorhinal or insular cortex) and the neocortex (the middle temporal or superior frontal cortex). The average number of the largest three values obtained for each region was calculated and scored from (0) to (4): (0) absent; (1) >0–1; (2) >1–3; (3) >3–9; (4) >9. The degree of SyαC-positive LB-related neurites was graded from (−) to (+++): (−) absent; (±) rare; (+) some; (++) many; (+++) numerous. The progression of Lewy pathology in the cerebrum was classified from stages I to IV, based on the number of LB and the degree of LB-related neurites, by our staging [23] (Table 1). The progression of Alzheimer pathology was classified from stages I to VI and from stages A to C by Braak staging [4]. The degree of neuronal loss in the substantia nigra and locus ceruleus was evaluated with HE staining. In the substantia nigra, the total number of pigmented neurons in each quarter was counted and scored from (0) to (4): (0) ≥500; (1) >300–500; (2) >150–300; (3) >50–150; (4) ≤50, whereas in the locus ceruleus, the number of pigmented neurons was counted and scored from (0) to (4): (0) ≥50; (1) >30–50; (2) >15–30; (3) >5–15; (4) ≤5. In addition, Lewy pathology in the substantia nigra and locus ceruleus was evaluated with SyαC immunostaining. The number of SyαC-positive LB was counted and scored from (0) to (4): in the substantia nigra, (0) absent; (1) 1–10; (2) 11–25; (3) 26–50; (4) >50, and in the locus ceruleus, (0) absent, (1) 1–5; (2) 6–10; (3) 11–20; (4) >20. The degree of SyαC-positive LB-related neurites was graded from (−) to (+++): (−) absent; (±) rare; (+) some; (++) many; (+++) numerous. The degree of spongiform change in the transentorhinal cortex was evaluated with HE staining, and was scored from (0) to (4): (0) absent; (1) very mild; (2) mild; (3) moderate; (4) severe. The degree of LB-related neurites in the CA2–3 region of the hippocampus was evaluated with SyαC-immunostaining, and was scored from (0) to (4): (0) absent; (1) rare; (2) some; (3) many; (4) numerous. The rate of coexistence of LB with NFT in the amygdala was evaluated with double-immunostaining of SyαC and AT8, and was represented as the percentage of double-positive neurons in SyαC-positive and double-positive neurons.
All statistical analyses were performed using SPSS software (version 10.0) for Windows. Analysis of variance (ANOVA) was performed for continuous variables such as the scores between subtypes. If there was a significant difference, Tukey tests were performed to establish where the difference lay. P values under 5% were accepted as indicating significance.
For immunoelectron microscopy, small blocks including the amygdala in the DLB and AD cases were cut into 40-μm-thick vibratome sections and immunostained with SyαC (1:100) by the ABC procedure. The sections were postfixed with 2.5% glutaraldehyde and 2% OsO4, embedded in Epon, cut into ultrathin sections, and stained with lead citrate.
Results
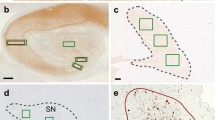
DLB was pathologically and immunohistochemically defined as having Lewy pathology of stages I–IV by our staging, and was classified into some pathological subtypes according to either Lewy pathology or Alzheimer pathology. The limbic type and neocortical type corresponded to stages I–II and stages III–IV of Lewy pathology, respectively, whereas the pure form, common form and AD form corresponded to stages I–II/stages 0–A, stages II–III/stages B–C, and stages IV–VI/stage C of Alzheimer pathology, respectively. Consequently, 34 out of 51 cases fulfilled the definition of DLB, and were divided into the combined subtypes of Lewy pathology and Alzheimer pathology (Table 2). The common form was the most frequent in both the limbic and neocortical types. The pure form was more frequently found in the limbic type, whereas the AD form was found only in the neocortical type. Immunohistochemically, many LB with LB-related neurites were found in the amygdala, and subsequently in the limbic cortex in both the limbic and neocortical types, whereas many LB and LB-related neurites were found in the neocortex in the neocortical type, with some found in the limbic type (Fig. 1A, B).

Some LB with many LB-related neurites in the amygdala of DLB. B Some LB with LB-related neurites in the transentorhinal cortex of DLB. C Some LB with only a few LB-related neurites in the amygdala of AD. D An LB without LB-related neurites in the transentorhinal cortex of AD. E Loss of pigmented neurons in the substantia nigra of DLB. F A few LB with LB-related neurites in the substantia nigra of DLB. G Spongiform change in the transentorhinal cortex of DLB. H Many LB-related neurites in the CA2–3 region of DLB. I Coexisting neurons (arrows) with some LB in the amygdala of DLB. J Coexisting neurons (arrows) with some NFT in the amygdala of AD. K LB-related neurites surrounding amyloid deposits in the amygdala of DLB. L LB-related neurites surrounding amyloid deposits in the amygdala of AD (LB Lewy body, DLB dementia with LB, SyαC anti-α-synuclein, AD Alzheimer’s disease, HE hematoxylin and eosin, NFT neurofibrillary tangle). A–D, F, H SyαC immunostaining; E, G HE staining I–L double immunostaining with SyαC (brown) and AT8 (blue)
Out of 51 cases, 17 were diagnosed as AD, because Lewy pathology did not meet stages I–IV, while Alzheimer pathology showed stages IV–VI/stage C corresponding to definite AD [25]. These cases showed LB corresponding to stages I–III in the amygdala, although the LB were accompanied by only a few LB-related neurites (Fig. 1C). LB were rare in the limbic cortex, and absent in the neocortex (Fig. 1D). Of the 17 cases, 4 also showed some LB in the CA1 and subiculum of the hippocampus, in addition to the amygdala.
ANOVA demonstrated that there were significant differences between combined subtypes of DLB and AD with respect to all factors (P<0.05).
In the 34 DLB cases, the substantia nigra and locus ceruleus showed a loss of pigmented neurons in varying degrees (Fig. 1E). The degree of neuronal loss in the substantia nigra and locus ceruleus is shown in each combined subtype of DLB, as well as in the AD cases (Table 3). The differences among the subtypes were more clearly found in the substantia nigra than in the locus ceruleus. Neuronal loss in the substantia nigra was most severe in the neocortical type/pure form, while it varied from mild to severe in the neocortical type/common and AD forms. The 17 AD cases showed only mild neuronal loss in the substantia nigra, although one case revealed moderate neuronal loss.
In the 34 DLB cases, the substantia nigra and locus ceruleus showed Lewy pathology in various degrees (Fig. 1F). The number of SyαC-positive LB and degree of SyαC-positive LB-related neurites in the substantia nigra and locus ceruleus are shown in each combined subtype of DLB, as well as in the AD cases (Table 4). The number of LB in the substantia nigra was largest in the neocortical type/common form, whereas it was smallest in the neocortical type/pure form, although the degree of LB-related neurites was not always proportional to the number of LB. Of the 17 AD cases, 10 showed no Lewy pathology in the substantia nigra; 6 out of the remaining 7 cases revealed only a few LB, with LB-related neurites rare, although 4 out of the 6 cases were the same cases as those with some LB in the hippocampus. Only one case showed moderate Lewy pathology with neuronal loss.
In the DLB cases, the transentorhinal cortex showed spongiform change in various degrees (Fig. 1G). The degree of spongiform change in the transentorhinal cortex is shown in each combined subtype of DLB, and in the AD cases (Table 5). Spongiform change was most severe in the neocortical type/AD form, whereas it was mildest in the neocortical type/pure form. A few AD cases showed moderate spongiform change, although in most cases it was only mild.
In the DLB cases, the CA2–3 region of the hippocampus showed LB-related neurites in various degrees (Fig. 1H). The degree of LB-related neurites in the CA2–3 region is shown in each combined subtype of DLB, and in the AD cases (Table 6). There were many LB-related neurites in the neocortical type/pure, common and AD forms, especially in the pure form. In contrast, they were absent in the AD cases, except for one case with some LB in the hippocampus.
Most of the DLB cases showed coexistence of LB with NFT to various degrees. These DLB cases showed many LB, in addition to coexisting neurons. The coexisting neurons demonstrated intermingled SyαC- and AT8-positive substances (Fig. 1I). In contrast, most of the AD cases had no LB other than coexisting neurons and many NFT. The coexisting neurons demonstrated a superimposed aggregation of SyαC-positive substances on AT8-positive NFT (Fig. 1J). SyαC- and AT8-positive neurites were found separately surrounding amyloid deposits in the DLB cases (Fig. 1K), whereas SyαC was double-immunostained in some of the AT8-positive neurites in the AD cases (Fig. 1L).
The rate of coexistence of LB with NFT in the amygdala is shown in each combined subtype of DLB, and in the AD cases (Table 7). The coexistence rate was highest in the neocortical type/AD form, reasonably high in the neocortical type/common form, and lowest in the neocortical type/pure form. In contrast, the coexistence rate was almost 100% in the AD cases, although 4 cases with some LB in the hippocampus revealed only a few LB.
Clinical data of all 51 cases are shown in each combined subtype of DLB, and in the AD cases (Table 8). In the DLB cases, the onset of disease was the oldest in the limbic type/pure and common forms, subsequently old in the neocortical type/common and AD forms, and middle-aged in the neocortical type/pure form. The initial symptom was dementia in the neocortical type/common and AD forms, whereas it was parkinsonism in the neocortical type/pure form. In the limbic type, the initial symptom was more frequently parkinsonism in the pure form, whereas it was dementia in the common form.
SyαC immunoelectron microscopy revealed compact bundles of SyαC-positive granulo-filamentous components measuring 10–20 nm in diameter in the LB and LB-related neurites of the DLB cases (Fig. 2A–C), whereas SyαC-positive filamentous components measuring 20–30 nm in diameter were joined to the loosened portion of bundles of paired helical filaments (PHF) in the LB and LB-related neurites of the AD cases (Fig. 2D–F).

An LB in the amygdala of DLB. B Higher magnification of A. Note SyαC-positive granulo-filamentous components (arrows). C An LB-related neurite in the amygdala of DLB. Note SyαC-positive granulo-filamentous components (arrows). D An LB in the amygdala of AD. Note SyαC-positive areas (arrows) E Higher magnification of D; SyαC-positive filamentous components (arrows) are joined to PHF. F An LB-related neurite in the amygdala of AD. SyαC-positive filamentous components (arrows) are joined to PHF (PHF paired helical filaments). A–F SyαC immunoelectron microscopy; A ×4,300; B, C ×19,000; D ×5,400; E ×18,000; F ×25,000
Discussion
Consensus guidelines for the pathological diagnosis of DLB classified DLB into three pathological subtypes: the brainstem-predominant, limbic and neocortical [24]. However, we previously examined 23 autopsied cases of DLB using ubiquitin or α-synuclein immunohistochemistry, and showed that these cases could not be practically classified into three subtypes, because most of them were of the neocortical type, not the brainstem-predominant type [14]. Therefore, we classified DLB into more detailed pathological subtypes, based on the regional incidence of LB, degree of Alzheimer pathology and degree of neuronal loss in the substantia nigra [14]. These subtypes were correlated to some extent with clinical features.
However, these previous subtypes of DLB were not sufficiently evaluated on the progress of Lewy pathology including LB and LB-related neurites in the cerebrum. Thereafter, we demonstrated that α-synuclein accumulates first in the axonal terminal, subsequently in the neuronal cell body, and finally in the dendrites, showing six developmental stages of LB [16], and that Lewy pathology in the cerebrum progresses first in the amygdala, subsequently in the limbic cortex, and finally in the neocortex [23]. Based on this progression of Lewy pathology, we quantitatively evaluated the incidence of LB and LB-related neurites in the amygdala, limbic cortex and neocortex of DLB cases, and graded Lewy pathology from stage I to stage IV [23]. Very recently, staging of Lewy pathology was performed using PD cases, and Lewy pathology was graded from stage I to stage VI [5]. This study suggested that Lewy pathology progresses from the lower brainstem to the cerebrum, which is in agreement with the results of other studies [7, 15]. However, Lewy pathology in the cerebrum in stages IV–VI overlaps with that of our stages in DLB cases, suggesting that these PD cases include DLB cases, or that PD has the pathological continuity with DLB [15].
In the present study, we reclassified DLB into combined subtypes of Lewy pathology and Alzheimer pathology. We classified DLB into limbic type or neocortical type by our staging of Lewy pathology. Lewy pathology in the brainstem-predominant type was defined to be below stage I. The cerebral type [19] was included either in the limbic type or the neocortical type. Consequently, all 34 DLB cases were divided into the limbic type or the neocortical type, but not the brainstem-predominant type. Some of the limbic type DLB and the neocortical type/pure form DLB cases may be clinically PD with dementia [24], because parkinsonism preceded dementia in these cases. However, these cases showed the direct pathological continuity with other DLB cases, probably PD cases [5], suggesting that PD with dementia lies among DLB and PD.
DLB is accompanied by Alzheimer pathology in varying degrees, so that entities such as senile dementia of Lewy body type [27] and Lewy body variant of AD [9] have been proposed. In addition, AD occasionally exhibits LB in the amygdala [1, 8, 21, 22]. These findings make the pathological boundary between DLB and AD indistinct.
In the present study, we classified DLB into pure form, common form or AD form by Braak staging for Alzheimer pathology. Alzheimer pathology of the AD form corresponded to that of AD. Consequently, the common form was most frequently found in both the limbic and neocortical types. The pure form was more frequently found in the limbic type, whereas the AD form was found only in the neocortical type. These findings show that DLB cases with severe Alzheimer pathology tend to show severe Lewy pathology, suggesting that Alzheimer pathology exacerbates Lewy pathology in DLB [6, 23].
Lewy pathology in all 17 AD cases did not meet the stages of Lewy pathology. Lewy pathology was found to be limited almost solely to the amygdala, was rare in the limbic cortex, and absent in the neocortex, although 4 out of the 17 cases also showed some Lewy pathology in the hippocampus. In addition, the LB were accompanied by only a few LB-related neurites even in the amygdala, unlike those in DLB. There are almost no differences in the distribution of Lewy pathology in AD cases between the present study and previous studies [1, 8, 21, 22]. Some studies showed Lewy pathology in the neocortex and substantia nigra, in addition to the amygdala, in AD cases [6, 15, 26]. These cases are likely to include the AD or common form DLB cases.
In the present study, the degree of Lewy pathology in the brainstem was evaluated by the degree of loss of pigmented neurons and the number of LB and LB-related neurites. Neither finding was proportional, and frequently showed a reciprocal relationship, in that the cases with more severe neuronal loss demonstrated a smaller number of LB, suggesting that the number of remaining LB-bearing neurons did not reflect the degree of Lewy pathology in the brainstem. In addition, the neocortical type/common and AD forms with severe Lewy pathology in the cerebrum showed various degrees of Lewy pathology from mild to severe in the brainstem, suggesting the possibility that Lewy pathology occasionally progresses from the cerebrum to the brainstem in DLB, unlike PD [5]. Most of the AD cases showed only mild neuronal loss without Lewy pathology in the substantia nigra. However, the 4 AD cases with some Lewy pathology in the hippocampus showed only a few LB with rare LB-related neurites, suggesting that Lewy pathology is exacerbated in both the cerebrum and brainstem in some of AD cases, although its extent and degree are much less than in those with DLB. Only one case showed moderate Lewy pathology with neuronal loss in the substantia nigra. This case should be regarded as a combination case of AD and PD.
The degree of spongiform change in the transentorhinal cortex was most severe in the neocortical type/AD form, whereas it was mildest in the neocortical/pure form. A few AD cases showed moderate spongiform change, although in most cases it was only mild. These findings suggest that both Lewy pathology and Alzheimer pathology are implicated in the development of spongiform change.
The degree of LB-related neurites in the CA2–3 region was severe in the neocortical type/pure, common and AD forms. In contrast, they were absent in all the AD cases except for one. These findings suggest that Lewy pathology in the axonal terminal is almost compatible with that in the neuronal cell body in DLB [12], although AD shows no Lewy pathology in the axonal terminal.
It has been reported that LB and NFT coexist in the same neurons of the limbic region in DLB [2, 13]. LB in AD also coexist with NFT, although they are morphologically different from those in DLB [1, 22]. In the present study, the coexistence rate of LB with NFT in DLB was highest in the neocortical type/AD form, reasonably high in the neocortical type/common form, and lowest in the neocortical type/pure form, suggesting that the rate depends on the number of NFT. In contrast, the coexistence rate was almost 100% in AD cases, regardless of the number of NFT. LB in DLB and AD immunohistochemically showed different morphology on the coexistence with NFT. Immunoelectron microscopy demonstrated that the LB and LB-related neurites in DLB were composed of α-synuclein-positive Lewy filaments, while the LB and LB-related neurites in AD were formed by the aggregation of α-synuclein on the PHF. These findings suggest that Lewy pathology in AD may develop only in the most favorable regions of Lewy pathology in PD and DLB [10, 11], that it may develop mainly in the neuronal cell body, not in the axonal terminal, and that it may develop secondary to the development of NFT, with the pathomechanism different from that in DLB.
In the present study, we reclassified DLB into combined subtypes of Lewy pathology and Alzheimer pathology. These subtypes were lined up on a spectrum, not only with Lewy pathology but also with other DLB-related pathologies. DLB could be pathologically differentiated, even from AD showing some Lewy pathology. The present study clarified the pathological entity of DLB, compared with AD.
References
Arai Y, Yamazaki M, Mori O, Muramatsu H, Asano G, Katayama Y (2001) α-Synuclein-positive structures in cases with sporadic Alzheimer’s disease: morphology and its relationship to tau aggregation. Brain Res 888:287–296
Arima K, Mizutani T, Alim MA, Tonozuka-Uehara H, Izumiyama Y, Hirai S, Ueda K (2000) NACP/α-synuclein and tau constutute two distinctive subsets of filaments in the same neuronal inclusions in brains from a family of parkinsonism and dementia with Lewy bodies: double-immunolabeling fluorescence and electron microscopic studies. Acta Neuropathol 100:115–121
Baba M, Nakajo S, Tu PH, Tomita T, Nakaya K, Lee VM, Trojanowski JQ, Iwatsubo T (1998) Aggregation of α-synuclein in Lewy bodies of sporadic Parkinson’s disease and dementia with Lewy bodies. Am J Pathol 152:879–884
Braak H, Braak E (1991) Neuropathological staging of Alzheimer-related changes. Acta Neuropathol 82:239–259
Braak H, Del Tredici K, Rüb U, Vos RAI de, Jansen Steur ENH, Braak E (2003) Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol Aging 24:197–211
Brown DF, Dababo MA, Bigio EH, Risser RC, Eagan KP, Hladik CL, White CL (1998) Neuropathologic evidence that the Lewy body variant of Alzheimer’s disease represents coexistence of Alzheimer disease and idiopathic Parkinson’s disease. J Neuropathol Exp Neurol 57:39–46
Del Tredici, Rüb U, Vos RAI de, Boel JRE, Braak H (2002) Where does Parkinson’s disease pathology begin in the brain? J Neuropathol Exp Neurol 61:413–426
Hamilton RL (2000) Lewy bodies in Alzheimer’s disease: a neuropathological review of 145 cases using α-synuclein immunohistochemistry. Brain Pathol 10:378–384
Hansen LA, Salmon D, Galasko D, Masliah E, Katzman R, De Teresa R, Thal L, Pay MM, Hofstetter R, Klauber M, Rice V, Butters N, Alford M (1990) The Lewy body variant of Alzheimer’s disease: a clinical and pathologic entity. Neurology 40:1–8
Harding AJ, Stimson E, Henderson JM, Halliday GM (2002) Clinical correlates of selective pathology in the amygdala of patients with Parkinson’s disease. Brain 125:2431–2445
Iseki E, Odawara T, Suzuki K, Kosaka K, Akiyama H, Ikeda K (1995) A pathological study of Lewy bodies and senile changes in the amygdala in diffuse Lewy body disease. Neuropathology 15:112–116
Iseki E, Marui W, Akiyama H, Ueda K, Iwatsubo T (1998) Degenerative terminals of the perforant pathway are human α-synuclein-immunoreactive in the hippocampus of patients with diffuse Lewy body disease. Neurosci Lett 258:81–84
Iseki E, Marui W, Kosaka K, Ueda K (1999) Frequent coexistence of Lewy bodies and neurofibrillary tangles in the same neurons of patients with diffuse Lewy body disease. Neurosci Lett 265:9–12
Iseki E, Marui W, Kosaka K, Kato M, Yamamoto T, Ueda K (1999) Clinicopathological multiplicity of dementia with Lewy bodies. Neuropathology 19:386–394
Jellinger KA (2003) α-Synuclein pathology in Parkinson’s disease and Alzheimer’s disease brain: incidence and topographic distribution-a pilot study. Acta Neuropathol 106:191–201
Katsuse O, Iseki E, Marui W, Kosaka K (2003) Developmental stages of cortical Lewy bodies and their relation to axonal transport blockage in brains of patients with dementia with Lewy bodies. J Neurol Sci 211:29–35
Kosaka K (1978) Lewy bodies in cerebral cortex. Report of three cases. Acta Neuropathol (Berl) 42:127–134
Kosaka K (1990) Diffuse Lewy body disease in Japan. J Neurol 237:197–204
Kosaka K, Iseki E, Odawara T, Yamamoto T (1996) Cerebral type of Lewy body disease. Neuropathology 16:32–35
Leigh PN, Probst A, Dale GE, Power DP, Brion JP, Dodson A, Anderson BH (1989) New aspect of the pathology of neurodegenerative disorders as revealed by ubiquitin antibodies. Acta Neuropathol 79:61–72
Lippa CF, Fujiwara H, Mann DM, Giasson B, Baba M, Schmidt ML, Nee LE, O’Connel B, Pollen DA, St George-Hyslop P, Ghetti B, Nochlin D, Bird TD, Cairns NJ, Lee VM, Iwatsubo T, Trojanowski JQ (1998) Lewy bodies contain alteredα-synuclein in brains of many familial Alzheimer’s disease patients with mutations in presenilin and amyloid precursor protein genes. Am J Pathol 153:1365–1370
Marui W, Iseki E, Ueda K, Kosaka K (2000) Occurrence of human α-synuclein-immunoreactive neurons with neurofibrillary tangle formation in the limbic areas of patients with Alzheimer’s disease. J Neurol Sci 174:81–84
Marui W, Iseki E, Nakai T, Miura S, Kato M, Ueda K, Kosaka K (2002) Progression and staging of Lewy pathology in brains from patients with dementia with Lewy bodies. J Neurol Sci 195:153–159
McKeith IG, Glasko D, Kosaka K, Perry EK, Dickson DW, Hansen LA, Salmon DP, Lowe J, Mirra SS, Byrne EJ, Lennox G, Quinn NP, Edwardson JA, Ince PG, Bergeron C, Burns A, Millar BL, Lovestone S, Collerton D, Jansen ENH, Ballard C, Vos RAI de, Willcock GK, Jellinger KA, Perry RH (1996) Consensus guidelines for the clinical and pathologic diagnosis of dementia with Lewy bodies (DLB): report of the consortium on DLB workshop. Neurology 47:1113–1124
Mirra SS, Heyman A, McKeel D, Sumi SM, Crain BJ, Brownlee LM, Vogel FS, Hughes JP, Belle G van, Berg L (1991) The Consortium to Establish a Registry for Alzheimer’s Disease (CERAD). Part II. Standarization of the neuropathologic assessment of Alzheimer’s disease. Neurology 41:479–486
Parkkinen L, Soininen H, Alafuzoff I (2003) Regional distribution of α-synuclein pathology in unimpaired aging and Alzheimer’s disease. J Neuropathol Exp Neurol 62:363–367
Perry RH, Irving D, Blessed G, Fairbairn A, Perry EK (1990) Senile dementia of Lewy body type: a clinically and neuropathologically distinct form of Lewy body dementia in the elderly. J Neurol Sci 95:119–139
Spillantini MG, Schmidt ML, Lee VM, Trojanowski JQ, Jakes R, Goedert M (1997) α-Synuclein in Lewy bodies. Nature 388:839–840
Acknowledgements
This research was supported by Grants-in-aid from the Japan Ministry of Education, Science, Sports and Culture, and from the Japan Ministry of Health, Labor and Welfare. We are grateful to Dr. T. Ishii for the generous supply of antibody.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Marui, W., Iseki, E., Kato, M. et al. Pathological entity of dementia with Lewy bodies and its differentiation from Alzheimer’s disease. Acta Neuropathol 108, 121–128 (2004). https://doi.org/10.1007/s00401-004-0869-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00401-004-0869-4