
Abstract
The pure-tone thresholds of four domestic female chickens were determined from 2 Hz to 9 kHz using the method of conditioned suppression/avoidance. At a level of 60 dB sound pressure level (re 20 μN/m2), their hearing range extends from 9.1 Hz to 7.2 kHz, with a best sensitivity of 2.6 dB at 2 kHz. Chickens have better sensitivity than humans for frequencies below 64 Hz; indeed, their sensitivity to infrasound exceeds that of the homing pigeon. However, when threshold testing moved to the lower frequencies, the animals required additional training before their final thresholds were obtained, suggesting that they may perceive frequencies below 64 Hz differently than higher frequencies.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Although audiograms for over 40 species of birds have been published, almost all of them have confined their testing to the animals’ mid- and high-frequency hearing range with the result that little is known about the ability of birds to hear low-frequency sounds (cf. Dooling 2002). The one exception is the homing pigeon which was shown many years ago to hear lower frequency sounds than humans, i.e., they can hear infrasound (Kreithen and Quine 1979). However, the acoustical conditions in which the pigeon audiogram was obtained differed from those of other audiograms, making direct comparisons difficult. Specifically, the pigeons were tested in a small airtight pressure chamber originally designed to test altimeters, whereas the audiograms of mammals, as well as those of other birds, have been obtained in free-field acoustic conditions. Because the pressure chamber was small (30 × 44.5 × 30 cm), it was not possible to use it to determine the low-frequency hearing ability of humans for comparison and thresholds were not attempted on another small animal. Thus, although it was undeniable that pigeons were sensitive to low frequencies, there was some question as to exactly how they would compare with other animals when tested in the same acoustic conditions (Heffner and Heffner 2007).
Recently, we determined the audiogram of the pigeon using the same acoustic environment and equipment used to test humans and other mammals (Heffner et al. 2013). Although the resulting low-frequency thresholds were not quite as low as those found by Kreithen and Quine (1979), we confirmed their finding that pigeons are more sensitive to low-frequency sounds than any mammal yet tested, including humans. Given these results, the next question was whether the ability to hear infrasound is a general characteristic of birds or whether it is limited to certain lifestyles, such as long distance navigation as suggested by Kreithen and Quine (1979). To address this question, we determined the audiogram of the domestic chicken, a species that does not navigate long distances. The results showed that not only do domestic chickens hear infrasound, but they are even more sensitive to it than pigeons. In addition, as testing progressed to the lower frequencies, we noticed an unexpected change in performance at 32 Hz in that the chickens required additional training at this frequency before their final thresholds emerged. This observation suggested that their perception of infrasound may have differed qualitatively from that of higher frequencies.
Methods
The method of conditioned suppression/avoidance was used to obtain absolute thresholds for chickens for pure tones ranging from 2 to 9,000 Hz. Four hens were trained to peck a key to obtain access to food and to stop pecking in the presence of a tone to obtain access to food and avoid electric shock that was delivered through bead chains around the base of their wings. The tympanic membranes of one hen were perforated and the animal retested to determine if it was using its ears or some other modality to detect low frequencies. A detailed description of the equipment and procedure can be found elsewhere (Heffner et al. 2013).
Animals
Four female chickens (Gallus gallus domesticus) were obtained from a local breeder. Two of them were Black sex-linked (chickens A and B) and the other two were Golden Comet (chickens C and D). The chickens were 21 weeks old when testing began, and 36 weeks old upon its completion. They were group housed in a room with free access to water. Chicken food (Purina Layena crumbles) was used as a reward and the animals were weighed daily to monitor their health and deprivation level.
Behavioral apparatus
Testing was conducted in a double-walled sound chamber (IAC model 1204; Industrial Acoustics Co., Bronx, NY, USA; 2.55 × 2.75 × 2.05 m), the walls and ceiling of which were lined with eggcrate foam and the floor carpeted to reduce sound reflections. The chickens were tested in a cage (50 × 30 × 42 cm) constructed of one-inch (2.54 cm) wire mesh that was mounted 92 cm above the floor on a tripod. A response key was constructed using a plastic disk (2.2 cm diameter, 1.5 cm thick) with a red LED embedded in it. The response key was mounted vertically 34 cm above the floor of the cage and the LED was momentarily turned off when the key was pecked. Access to chicken food was provided by a solenoid-operated food tray that, when operated, would come up underneath the bottom of the cage (below the response key) to allow the chicken to eat from it for 3.5 s; the entire feeder mechanism was below the level of the cage floor so that it would not interfere with the sound field.
Electric shock was produced by a Coulbourn constant current shock generator that was connected via alligator clips hanging from the top of the cage to bead chains around the base of the chickens’ wings [for a description of the bead chain procedure, see Hoffman ( 1960) and Stein et al. (1971)]. The animals were trained and tested using shock levels of 0.2–0.5 mA, 1.5-s duration, with the level adjusted for each animal to the lowest level that produced a consistent avoidance response to an obviously audible signal. The shock was defined as mild because the chickens never developed a permanent fear of pecking the key and returned to pecking the key after the shock had been delivered. A 25-watt light bulb, placed above the loudspeaker, was turned on coincident with the shock.
Acoustical procedures
Pure tones were generated (Agilent 33220A function generator), attenuated (Tucker-Davis Technologies model PA4 attenuator) and gated on and off (Coulbourn S84-04 rise-fall gate) at zero crossing as follows: Frequencies of 250 Hz and higher were pulsed 400 ms on, 100 ms off (5 tone pulses) with a rise-decay of 40 ms for 250 Hz, 20 ms for 500 Hz, and 10 ms for 1–9 kHz; frequencies from 2 to 125 Hz were gated on with a 100-ms rise-decay time for 2 s—they were not pulsed because the longer rise-decay time would have significantly reduced the on-full duration of the signal and possibly raised the threshold. Frequencies above 4 Hz were filtered with a band-pass filter (Krohn-Hite 3550) set 1/3 octave above and below the tone’s frequency; for 2 and 4 Hz, the filter was set on double low-pass at 1/3 octave above the frequency of the tone. Finally, the signal was amplified (Crown D-75 amplifier), monitored on an oscilloscope, and sent to a loudspeaker (with the same frequency often tested with different speakers). The loudspeakers used were a TC Sounds Axis 15-in (38.1 cm) subwoofer in an unported enclosure (65 × 65 × 120 cm) for 2–16 Hz, a Paradigm Servo 15 subwoofer for 16–32 Hz, a 12-in (30.5 cm) woofer for 32–64 Hz, an Infinity RS 2000 for 64 Hz to 1 kHz, an enclosed Tang Band W3-319SF 3-in (7.2 cm) full-range driver for 1–4 kHz, and a Foster Ribbon tweeter or Motorola piezoelectric tweeter for frequencies from 2.8 to 9 kHz. All speakers were placed at least 1 m in front of the test cage.
The sound pressure level (SPL re 20 μN/m2) of the stimulus was measured and checked for overtones using a 1-inch (2.54-cm) microphone (Brüel & Kjaer 4145) or a ¼-inch (0.635 cm) microphone (Brüel & Kjaer 4939, calibrated down to 2 Hz), measuring amplifier (Brüel & Kjaer 2610), and a spectrum analyzer (Zonic A&D 3525 FFT Analyzer). Sound measurements were taken by placing the microphone in the position occupied by a chicken’s head when it was pecking the response key and, with the exception of the two subwoofers, pointing it directly ahead toward the loudspeaker (0º incidence). The Paradigm subwoofer (46 × 55 × 51 cm) was placed on the floor of the chamber in front of the test cage; the TC Sounds Axis 15 subwoofer was placed in front of the cage and turned 180° to prevent the chicken from seeing the movement of the speaker diaphragm (the microphones used are omidirectional at low frequencies so no correction for orientation needed to be applied). No overtones were present at the sound pressure levels used to obtain thresholds and no spectral splattering was observed during the onset of the tones. The background noise level in the sound chamber was relatively low; although modern buildings often have significant low-frequency noise produced by the heating-ventilating-air conditioning system (HVAC), our laboratory is located in a building constructed in 1929 in which the HVAC equipment for our area is located in a room external to the building.
Behavioral procedure
The chickens were first trained to peck the response key on a random ratio schedule to obtain access to food for 3.5 s. Next they were trained to peck the key during trials in which no sound was presented (“silent” trials), but not during trials in which a tone was present (“tone” trials). This was done by rewarding an animal at the end of a trial with food if it (1) pecked during a silent trial or (2) did not peck during a tone trial. In addition, an animal received a mild shock if it had pecked during a tone trial.
A session consisted of a series of 2.4-s trials, each with an intertrial interval of no less than 10 s. Because each trial was initiated by a key peck, the length of the intertrial interval exceeded 10 s if the chicken stopped to eat a reward or had just received a shock, but was typically less than 10 s. The response of a chicken was defined by whether or not it pecked during the last 300 ms of the trial, giving the animal sufficient time to react to the tones. If the chicken did not peck during this 300-ms period, an avoidance response was recorded. The avoidance response (withholding key pecks) was classified as a “hit” if a tone had been presented and as a “false alarm” if there had been no tone. Each trial had a 22 % probability of containing a tone. An animal received access to food at the end of a trial if it had made a correct response, that is, if it pecked during a silent trial (correct rejection) or if it stopped key pecking during a tone trial (hit). A “miss” (pecking during the last 300 ms of a tone trial) was followed by a 1.5-s shock.
To reduce the effect of spurious pauses, a trial did not begin until the chicken pecked the key, which also meant that a tone was only presented when an animal’s head was in position in front of the response key. Test sessions typically consisted of 50–100 tone trials (and associated silent trials) and lasted from 30 to 90 min depending on the individual chicken and how much food it wished to eat.
Hit and false alarm rates were determined for each block of trials (5–10 tone trials interspersed among 18–36 silent trials) for each frequency. The hit rate was corrected for the false alarm rate to produce a performance measure according to the following formula: Performance = Hit rate−(False alarm rate × Hit rate), which can also be expressed as Performance = Hit rate × Correct rejection rate, where Correct rejection rate = 1−False alarm rate (Heffner and Heffner 1995). This measure proportionally reduces the hit rate by the false alarm rate and varies from 0 (no hits) to 1 (100 % hit rate with no false alarms).
Absolute thresholds were determined by presenting blocks of 5–10 tone trials (all at the same amplitude) and then reducing the amplitude of successive blocks in 10-dB steps until the chicken no longer responded to the tone above the 0.01 chance level (binomial distribution). Once a preliminary threshold had been obtained, final threshold determination was conducted by presenting blocks of 10 tone trials in which the amplitude of the tone of each block was varied in 5-dB steps extending from 10 dB above to 10 dB below the estimated threshold. Trial blocks of higher intensities were given after a threshold had been obtained to ensure that an animal was still attending to the tone. Threshold was defined as the amplitude corresponding to a performance of 0.50, which was usually determined by interpolation. Threshold testing for a particular frequency was considered complete when the thresholds obtained in at least three different sessions were stable and within 3 dB of each other. After an audiogram had been completed, each threshold was rechecked to ensure reliability.
Tympanic membrane perforation
Chicken B was anesthetized with isoflurane and, with the aid of a dissecting microscope, multiple perforations were made in the tympanic membranes of both ears with a 20 ga. hypodermic needle, leaving the columellae intact. The animal was then tested daily at 32, 250, and 2,000 Hz for 2 weeks.
Results
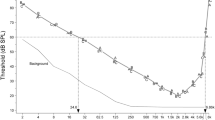
Acclimating the chickens to the test cage and training them to peck the key to obtain food and to withhold pecking in the presence of a 1-kHz tone took 17–19 sessions. Threshold testing was begun at 1 kHz, progressing to the higher frequencies, and then systematically moving to the lower frequencies, beginning with 500 Hz. As shown in Fig. 1, the absolute thresholds of the four chickens are in close agreement, with their 60-dB hearing range extending from 9.1 Hz to 7.2 kHz, a range of 9.6 octaves. Unlike most mammals, but like most other birds, chickens are unable to hear above 10 kHz. On the other hand, their low-frequency sensitivity extends lower than that of any mammal yet tested, including the elephant (Heffner and Heffner 1982). The low-frequency thresholds were probably not affected by background noise as the noise levels were at least 10 dB below the chickens’ thresholds for 64 Hz and lower frequencies.

Audiograms of four female chickens (A, B, C, and D). The audiogram line is the mean threshold of the four animals. SPL is the sound pressure level re 20 μN/m2
Perforating the tympanic membrane in mammals is known to cause a hearing loss that is greatest at low frequencies (e.g., Voss et al. 2001), and that is also the case in chickens. Six hours after making large perforations in both tympanic membranes of chicken B, the animal’s thresholds had increased by 22 dB at 2 kHz, 43 dB at 250 Hz, and by more than 47 dB at 32 Hz (the animal could not detect 32 Hz at 95 dB, the maximum undistorted amplitude of the speaker). These results demonstrate that the chicken was detecting low-frequency sound with its ears and not through substrate vibration or other non-auditory route. Thresholds gradually improved as the membranes healed and were within 3 dB of the preoperative thresholds after 14 days.
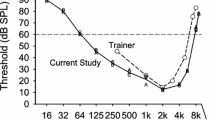
In the process of determining the audiogram, we noticed anomalous behavior while testing 32 Hz. When training animals to response to sound, the first threshold is often a “false threshold” in that an animal needs to “learn to listen” before it gives its lowest threshold. This can be seen in Fig. 2 for the first threshold that was determined, 1-kHz, where the average difference between the thresholds obtained in an animal’s initial and final session for that frequency was relatively large (4.5 dB). As testing moved to higher frequencies, and later on to the lower frequencies, the animals were now experienced observers and the average difference between the initial and final test sessions was less than 3 dB, with one exception. The exception occurred at 32 Hz where the difference between the initial and final threshold sessions ranged from 5.2 to 13.5 dB, for an average improvement of 8.6 dB.

Improvement in threshold with practice. Threshold testing began at 1 kHz and progressed to 9 kHz, following which the lower frequencies were tested. The greatest improvement in threshold with practice typically occurs at the first frequency on which an animal is trained, in this case, 1 kHz. However, all four chickens showed the most improvement with practice at 32 Hz, but only when 32 Hz was retested after testing was completed at 16 Hz. One interpretation of this observation is that the perception of 32 Hz and lower frequencies is mediated by a different modality than higher frequencies. The four chickens are indicated by the letters A, B, C, and D. The line through the thresholds is the mean of the four animals
In addition to the unexpected differences between the initial and final 32-Hz test sessions, the duration of this frequency’s “false threshold” was also atypical. Whereas improvements in thresholds at a particular frequency typically occurred by the second session in which it was tested, the initial thresholds at 32 Hz were stable for three consecutive sessions and so testing progressed to the next lower frequency, 16 Hz. Expecting that the chickens would be less sensitive at 16 Hz than at 32 Hz, we were surprised when their 16-Hz thresholds were either equal to or lower than the 32-Hz thresholds. After completing the 16-Hz testing, the 32-Hz thresholds were retested, at which time they improved to their final levels. A similar recheck of thresholds at higher frequencies found no significant change in thresholds.
Several possible explanations of the 32-Hz threshold shift were considered. First, we considered that it might have been due to changing from one loudspeaker to another. However, the same speaker, the 12-in woofer, had been used both before and after the threshold shift. Moreover, rechecking the 32-Hz thresholds using the Paradigm Servo 15 subwoofer gave the same results as the 12-in woofer. Therefore, the threshold shift could not be accounted for by loudspeaker differences. A second possibility was that the background noise level at 32 Hz decreased as that frequency was tested. However, no change in the background noise level was noted on the spectrographic records that were routinely taken whenever the loudspeaker or frequency was changed. In addition, subsequent thresholds for 32 Hz obtained over the next several months for three of the chickens did not vary implying that the background noise level was stable. Thus, we propose a third possibility, namely that while being trained to detect 16 Hz, the chickens changed their strategy for detecting low frequencies, suggesting that frequencies of 32 Hz and lower are in some way perceived or interpreted as different from higher frequencies.
Discussion
Chicken audiograms
There have been two previous studies of absolute thresholds of chickens. The first used an unconditioned procedure in which the regular peeping of chicks was momentarily suppressed by presenting tones in octave steps from 125 Hz to 4 kHz (Gray and Rubel 1985). The thresholds obtained on chicks at 0 and 4 days after hatching are not as low as those found here. This difference is most likely due to the use of an unconditioned procedure with the chicks, but may also be due to the immaturity of the animals. The second study used a go/no-go procedure in which chickens were trained to peck an “observing” key to begin a trial and to peck a “reporting” key when they detected a tone (Saunders and Salvi 1993). The animals were rewarded with food for pecking the reporting key when a tone was presented (Hit) and for not pecking the key when no tone was presented (Correct Rejection). Pecking the key when no tone was presented (False Alarm) was punished with a 10 s time out whereas failing to peck the key when a tone was presented (Miss) resulted in a missed reward. Test frequencies ranged from 250 Hz to 5.5 kHz in 0.1-octave steps. The absolute thresholds obtained by Saunders and Salvi were much lower than those obtained by Gray and Rubel. However, they were, on average, 11 dB less sensitive than the thresholds reported here, possibly because their procedure punished False Alarms more severely than Misses, thus making the animals less likely to respond to tones near threshold.
Ability of birds to detect infrasound
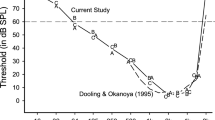
Infrasound is anthropocentrically defined as sound below the 20-Hz nominal low-frequency hearing limit of humans, although this is not an absolute limit as we can hear lower frequencies if the amplitude is sufficiently high (e.g., Jackson et al. 1999). Among mammals, humans have relatively good low-frequency hearing and only the elephant is known to hear lower (Heffner and Heffner 1982). However, of the two species of birds whose low-frequency hearing has been behaviorally tested, both can detect infrasound. Figure 3 compares the chicken audiogram with that of humans and pigeons. At high frequencies, humans have better hearing than either chickens or pigeons and they are also more sensitive at their best frequency. But at low frequencies both chickens and pigeons are decidedly more sensitive than humans. The observation that at least some birds can hear much lower frequencies than larger animals (e.g., humans) supports the view that a small size is not necessarily a limiting factor in detecting low frequency sounds (cf., Heffner et al. 2001).

The first discovery of a bird that could detect infrasound was made by Kreithen and his colleagues who were studying pigeons (Kreithen and Quine 1979; Yodlowski et al. 1977). Their interest in this issue stemmed from the hypothesis that homing pigeons might use infrasound for long distance navigation, an idea that is still being explored (Hagstrum 2013). Although our results do not bear directly on the use of infrasound for homing, they do indicate that infrasound detection is not limited to birds that fly long distances as flight in the domestic chicken is limited to flying up into trees or other perches, either to escape predators or to roost at sunset. Moreover, although the chicken’s wild ancestor, the red Jungle Fowl (Gallus gallus), is considered by comparison to be a strong flyer, it prefers to walk away from an approaching human rather than to fly (Collias and Collias 1967). Just how far jungle fowl may fly is illustrated by the following observation on the migration of jungle fowl (Giles 1932): “In the Chiengsen region the birds are apparently trying to cross the [Mekong] river eastwards, where it is about 700 yards [640 m] across. In crossing, the birds fly up as high as they can go, and then attempt to glide across. A large number, however, fall in the river and are drowned.” Thus, we conclude that the ability to detect infrasound serves other functions in chickens and possibly in birds in general.
There is physiological evidence for infrasonic hearing in other birds, especially galliformes, which are an avian Order comprising large bodied, ground feeding birds such as chickens, turkeys, grouse, quail and pheasant. Evidence for this comes from electrophysiological thresholds in the auditory midbrain nucleus of the Guinea fowl in which responses could be recorded down to 5 Hz. In addition, a quantitative morphological analysis of the inner ear of seven galliform species found over half of the basilar papilla tuned to frequencies below 1 kHz (Corfield et al. 2013; Theurich et al. 1984). Thus, the ability to hear infrasound may be widespread among birds.
Does infrasound detection involve a different modality?
The unusual improvement in the chickens’ 32-Hz thresholds after being tested at 16 Hz raises the possibility that the perception of very low frequencies involves a different sensory modality, perhaps one in which infrasound is perceived, not as sound, but as tactile pressure change. This could explain why the chickens required extra training as they had to shift their attention from audition to the other modality. If the detection of infrasound relies on a different modality, then one might expect that a different receptor would be involved. The avian ear contains several organs that might serve to detect infrasound. One such structure is the paratympanic organ which Kreithen and his colleagues suggested might serve as a barometer or altimeter (Kreithen and Quine 1979; von Bartheld and Giannessi 2011). Another is the macula lagenae, which is thought to have a vestibular function (Kaiser and Manley 1996; Khorevin 2008). However, physiological evidence indicates that infrasound is detected by the basilar papilla, the avian counterpart to the Organ of Corti in mammals.
Physiological recordings in the chick cochlear nucleus have found units that respond to frequencies as low as 10 Hz (Warchol and Dallos 1989b). Unlike the responses to higher frequencies that show typical tuning curves with an identifiable characteristic frequency, the very low-frequency units have tuning curves that either resemble a low-pass filter or else have very broad band-pass responses. Moreover, these low-frequency units receive input from the apex of the basilar papilla (Warchol and Dallos 1989a), the tip of which contains hair cells that are thought to be suitable for the perception of very low frequencies because they have the morphological characteristics of both vestibular and auditory organs (Lavigne-Rebillard et al. 1985).
A similar situation appears to exist for the pigeon in which units sensitive to frequencies below 20 Hz respond differently from ordinary auditory units. Specifically, fibers innervating the apex of the basilar papilla have a relatively high spontaneous activity that is modulated by infrasound and do not appear to be tuned to a particular frequency (Schermuly and Klinke 1990a, b). These infrasound units are so different from ordinary auditory units that it has been suggested that they may form a distinct class (Schermuly and Klinke 1990a). The pigeon inner ear also contains atypical hair cells that resemble those described in the chicken by Lavigne-Rebillard et al. (1985), but they are not located on the basilar papilla and their function is unknown (Schermuly et al. 1991).
In this context, it might be noted that we did not see a similar improvement in thresholds when obtaining the pigeon audiogram (Heffner et al. 2013). However, the pigeons had been used in auditory studies involving low-frequency as well as high-frequency sounds for several years before their audiogram was obtained and thus may have already been experienced in detecting infrasound in the test situation. Thus, although we have no data to indicate whether they would have shown the same practice effect as the chickens, we cannot rule it out.
In conclusion, the observation that units in the chicken and pigeon basilar papillae sensitive to infrasound respond differently from ordinary auditory units lends support to the idea that infrasound may give rise to a different sensation than higher-frequency sounds. This in turn suggests that the chicken audiogram presented here may have two components, one based on the perception of sound, the other based on a different modality, perhaps the perception of pressure change. The results shown in Fig. 2 suggest that the perception of sound goes from 9 kHz down to about 32 Hz, with the initial threshold at 32 Hz representing the threshold for the perception of sound. The portion of the audiogram representing the other modality may be the final thresholds for 32 Hz and lower frequencies. Thus, there may be two overlapping sensitivity curves, with the sensitivity of the second being an average of 9 dB more sensitive at 32 Hz than the sensitivity to sound.
References
Collias NE, Collias EC (1967) A field study of the red jungle fowl in North-Central India. Condor 69(4):360–386
Corfield JR, Krilow JM, Vande Ligt MN, Iwaniuk AN (2013) A quantitative morphological analysis of the inner ear of galliform birds. Hear Res 304:111–127
Dooling R (2002) Avian hearing and the avoidance of wind turbines. National Renewal Energy Laboratory, US Department of Energy. NREL/TP-500-30844. doi:10.2172/15000693
Giles FH (1932) Migration of jungle-fowl. Nat Hist Bull Siam Soc 8(4):333–334
Gray L, Rubel EW (1985) Development of absolute threshold in chickens. J Acoust Soc Am 77(3):1162–1172
Hagstrum JT (2013) Atmospheric propagation modeling indicates homing pigeons use loft-specific infrasonic ‘map’ cues. J Exp Biol 216:687–699
Heffner RS, Heffner HE (1982) Hearing in the elephant: absolute sensitivity, frequency discrimination, and sound localization. J Comp Physiol Psychol 96:926–944
Heffner HE, Heffner RS (2007) Hearing ranges of laboratory animals. J Am Assoc Lab Anim 46:11–13
Heffner RS, Koay G, Heffner HE (2001) Audiograms of five species of rodents: implications for the evolution of hearing and the encoding of pitch. Hear Res 157:138–152
Heffner HE, Koay G, Hill EM, Heffner RS (2013) Conditioned suppression/avoidance as a method for testing hearing in birds: the domestic pigeon (Columba livia). Behav Res Methods 45:383–392
Hoffman HS (1960) A flexible connector for delivering shock to pigeons. J Exp Anal Behav 3:330
Jackson LL, Heffner RS, Heffner HE (1999) Free-field audiogram of the Japanese macaque (Macaca fuscata). J Acoust Soc Am 106:3017–3023
Kaiser A, Manley GA (1996) Brainstem connections of the macula lagenae in the chicken. J Comp Neurol 374:108–117
Khorevin VI (2008) The lagena (the third otolith endorgan in vertebrates). Neurophysiology 40:142–159
Kreithen ML, Quine DB (1979) Infrasound detection by the homing pigeon: a behavioral audiogram. J Comp Physiol A 129:1–4
Lavigne-Rebillard M, Cousillas H, Pujol R (1985) The very distal part of the basilar papilla in the chicken: a morphological approach. J Comp Neurol 238:340–370
Saunders SS, Salvi RJ (1993) Psychoacoustics of normal adult chickens: thresholds and temporal integration. J Acoust Soc Am 94(1):83–90
Schermuly L, Klinke R (1990a) Infrasound sensitive neurons in the pigeon cochlear ganglion. J Comp Physiol A 166:355–363
Schermuly L, Klinke R (1990b) Origin of infrasound sensitive neurons in the papilla basilaris of the pigeon: an HRP study. Hear Res 48:69–78
Schermuly L, Topp G, Klinke R (1991) A previously unknown hair cell epithelium in the pigeon cochlea: the papilla chaotic. Hear Res 53:49–56
Stein N, Hoffman HS, Stitt C (1971) Collateral behavior of the pigeon during conditioned suppression of key pecking. J Exp Anal Behav 15:83–93
Theurich M, Langner G, Scheich H (1984) Infrasound responses in the midbrain of the guinea fowl. Neurosci Lett 49:81–86
von Bartheld CS, Giannessi F (2011) The paratympanic organ: a barometer and altimeter in the middle ear of birds? J Exp Zool (Mol Dev Evol) 316:402–408
Voss SE, Rosowski JJ, Merchant SN, Peake WT (2001) How do tympanic-membrane perforations affect human middle-ear sound transmission? Acta Otolaryngol 121:169–173
Warchol ME, Dallos (1989a) Localization of responsiveness to very low frequency sound on the avian basilar papilla. ARO Abst 12:125
Warchol ME, Dallos P (1989b) Neural response to very low-frequency sound in the avian cochlear nucleus. J Comp Physiol A 166:83–95
Yodlowski ML, Kreithen ML, Keeton WT (1977) Detection of atmospheric infrasound by homing pigeons. Nature 265:725–726
Acknowledgments
We thank Drs. Geoffrey Manley and Christopher Von Bartheld for their helpful comments on an earlier draft of this manuscript and K. Marchetto for her help with the animals. The use of animals in this study was approved by the University of Toledo Animal Care and Use Committee.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Hill, E.M., Koay, G., Heffner, R.S. et al. Audiogram of the chicken (Gallus gallus domesticus) from 2 Hz to 9 kHz. J Comp Physiol A 200, 863–870 (2014). https://doi.org/10.1007/s00359-014-0929-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00359-014-0929-8