
Abstract
The purpose of this study was to determine if the Japanese quail, a domesticated, gallinaceous bird, could detect infrasound. Behavioral thresholds were determined for three birds, two males and one female, ranging from 16 Hz to 8 kHz. The animals’ hearing range, at a cutoff of 60 dB SPL (re 20 μN/m2), covers 6.88 octaves, ranging from 59.5 Hz to 7 kHz. All animals had the greatest sensitivity to 2 kHz, with an average threshold of 4.4 dB SPL. Although the birds’ threshold at 16 Hz was equivalent to that of humans, at no frequency did the birds’ sensitivity ever exceed that of humans. Therefore, the Japanese quail does not hear infrasound.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
It has long been known that pigeons (Columba livia) are sensitive to lower frequencies than humans can detect, i.e. they detect infrasound (Kreithen and Quine 1979). Although nonbehavioral assessments of the avian auditory system have suggested that several different species possess good low-frequency hearing, recent behavioral research has revealed that at least one other bird species also detects infrasound: the domestic chicken (Gallus gallus domesticus) (Hill et al. 2014). The original explanation for the pigeon’s infrasonic sensitivity was that they might use this signal as navigational cue, which subsequent studies showed to be plausible (Kreithen and Quine 1979; Quine and Kreithen 1981; Quine 1982). However, this cannot be the only explanation for avian infrasonic sensitivity, because the chicken does not home or engage in long-distance flight (Hill et al. 2014). Unfortunately, the development of a comprehensive explanation for infrasound sensitivity in birds is not currently possible because the low-frequency range and sensitivity of most bird species have not been behaviorally determined. Indeed, to date only five species have had their sensitivity below 250 Hz assessed: the brown-headed cow bird (Molothrus ater) (Heinz et al. 1977), budgerigar (Melopsittacus undulatus) (Heffner et al. 2016), bullfinch (Pyrrhula pyrrhula) (Schwartzkopff 1949), mallard duck (Anas platyrhynchos) (Hill 2017), and redwing black bird (Agelaius phoeniceus) (Heinz et al. 1977). Furthermore, only the budgerigar and mallard duck were tested below 125 Hz, and neither species could detect infrasound. Thus, there are currently only four species whose full audiograms have been determined on which to base a theory of why some birds hear infrasound while others do not.
Although psychoacoustic data are lacking, other lines of research support the possibility of sensitivity to infrasonic signals in several other avian species, especially those of the order Galliformes. First, spectral analyses of a variety of non-vocal, territorial and mating displays have identified four species whose displays produce strong low-frequency components (e.g. < 500 Hz): the peacock (Pavo cristatus) (Freeman and Hare 2015), ruffed grouse (Bonasa umbellus) (Garcia et al. 2012), ring-necked pheasant (Phasianus colchicus) (Heinz and Gysel 1970), and capercaillie (Tetrao urogallus) (Lieser et al. 2005). Additionally, research by Kane et al. (2018) has identified vibrotactile receptors coupled to the crest feathers of the peacock that are sensitive to mechanical displacement at resonant frequencies produced by peacock social and mating displays. This provides an alternative, nonacoustic mechanism by which peacocks may detect low-frequency signals and indicates that detection of these signals may occur in multiple sensory modalities. Finally, electrophysiological studies of the guinea fowl (Numida meleagris) midbrain (Theurich et al. 1984), found single units able to phase-lock responses to signals as low as 2 Hz. Therefore, a plausible hypothesis is that all gallinaceous birds potentially share a heightened sensitivity to low-frequency sounds. Indeed, a morphological analysis of the basilar papilla of seven species of gallinaceous bird (ruffed grouse, gray partridge (Perdix perdix), Japanese quail (Coturnix japonica), ring-necked pheasant, turkey (Meleagris gallopavo), spruce grouse (Falcipennis Canadensis), and peacock) found that in all birds tested, greater than 50% of the basilar papilla was tuned to frequencies lower than 1 kHz (Corfield et al. 2013).
Based on the previous observations of infrasound production in numerous gallinaceous species, a common, domesticated bird of this order was selected for investigation in the present study: the Japanese quail. Previous research indicated that this species has good hearing sensitivity in the 500 Hz to 5.6 kHz range, though no frequencies below 500 Hz were tested (Dooling and Okahonya 1995). We tested the hearing range and sensitivity of Japanese quail using a conditioned suppression/avoidance paradigm, with an emphasis on those frequencies below 500 Hz.
Methods
Absolute, pure-tone thresholds, from 16 Hz to 8 kHz, were determined for the Japanese quail using standard behavioral procedures (Heffner and Heffner 1995; Heffner et al. 2013). Using the conditioned suppression/avoidance procedure, three adult quail were trained to peck a key to obtain access to food and withhold this response when a tone was presented to avoid a mild, electric shock. Food reinforcement was available to the animal in the following two circumstances: on tone trials if they did not peck the key and on no-tone trials if they did peck the key.
Animals
Three Japanese quail were obtained from an online vendor (Stromberg’s, Pine River, MN), one female (animal A) and two males (animals B and C). Testing began when the birds were 19 weeks old and was completed at 51 weeks for animal A, 76 weeks for animal B, and 70 weeks for animal C. The animals were housed in individual cages (61 × 46 × 61 cm) and provided free access to water and poultry feed (Country Lane: 16% All Flock). During training and testing, food was withheld in the animals’ home cages, and provided as a reinforcer during sessions. To monitor the animals’ deprivation levels and ensure maintenance of a healthy body weight, animals were weighed prior to daily testing sessions.
Behavioral apparatus
Testing was conducted in an acoustically controlled environment (WhisperRoom Inc., model 8484E; 2.03 × 1.97 × 1.97 m), with sound-attenuating, acoustic foam lining the walls. Animals were tested in a cage (31 × 31 × 31 cm) constructed of ½-inch (1.27 cm) mesh hardware cloth, mounted 71 cm above the floor on four wooden legs. A red LED and transparent 2 cm diameter plastic key were attached to the arm of a microswitch, allowing for automated recording of responses. This apparatus was then mounted 16 cm above the cage floor and illuminated when a response was required from the animal. The initial key peck started a 2 s trial, after which the LED briefly turned off to signal the end of the trial. Correct responses resulted in access to poultry feed (Country Lane: 16% All Flock), provided by a specially-constructed food dispenser that was operated by a high-torque, metal-gear servo (Tower Pro, model MG995R). When operated, the food dispenser removed an opaque, lightweight plastic cover from a food reservoir directly under the front of the cage. Animals were permitted 2 s access to food before the dispenser slowly moved the cover back over the reservoir over the course of 0.5 s.
Incorrect responses resulted in a brief, mild shock, produced by a constant current shock generator (Coulbourn, model H-13–15) connected via bead chain connectors hanging from the top of the cage to bead chains around the base of the quails’ wings (Heffner et al. 2013). The animals were trained and tested using shock levels of 0.5 to 1.0 mA (250 to 290 V), 1.0 s duration, with levels adjusted on an individual basis to the minimum intensity that produced a consistent avoidance response when the signal was clearly audible.
Acoustic procedures
Acoustic frequencies ranged from 16 Hz to 8 kHz in one-octave steps, with additional testing at 2.8 kHz and 5.6 kHz. All signals were produced (Agilent function generator, model 33220A), attenuated (Sound Performance Lab, model 2602) and gated on and off (Coulbourn rise-fall gate, model S84-04) for all frequencies. All acoustic signals were pulsed 900 ms on, 100 ms off for 2 s, producing two presentations of the test signal each trial. Because the wavelength of a signal increases as the frequency is decreased, different rise-decay times were required prevent the speaker from producing a click due to abrupt signal onsets. Rise-fall times for signals from 16 to 125 Hz were 100 ms, 250 Hz was 40 ms, 500 Hz was 20 ms, and all signals from 1 to 8 kHz were 10 ms. The acoustic signal was routed through a band-pass filter (Krohn-Hite, model 3322) with cutoffs set to 1/3-octave above and below the test frequency. The signal was then amplified (Gemini, model X-01) and monitored visually on an oscilloscope (Tektronix, model TBS 1022) before being presented via loudspeaker to the animal. Because the frequency response of speakers vary, three different speakers were necessary to produce all test signals: Signals from 16 to 500 Hz were presented using a 15″ subwoofer (Dayton Audio, model RSS390HO-4) in an unported enclosure (50 × 50 × 50 cm), from 500 Hz to 2 kHz using a 3″ aluminum driver (HiVi, model B3N), and from 4 to 9 kHz using a piezoelectric speaker (GRS, model PZ1005).
All signal intensities were measured daily prior to testing. The average starting intensities of each acoustic signal are presented in Table 1. Starting intensities were varied by approximately ± 3 dB from session to session to more precisely identify each animal’s minimum audible sensitivity. Sound pressure levels (re 20 μN/m2) were measured using a 1/4 in (6.35 mm) microphone (Brüel & Kjaer, model 4954-A-011, calibrated down to 5 Hz) and measuring amplifier (Brüel & Kjaer, model 2610). During sound measurement, the microphone was placed in the same location as the animal’s head during signal presentations, and oriented toward the speaker (0° sound incidence). The measuring amplifier and microphone were calibrated prior to taking any measurements using a sound level calibrator (Brüel & Kjaer, model 4230). The spectrum of the signal was checked for the presence of overtones and distortion using the FFT function of the oscilloscope. Overtones and distortion were not present in any signal at threshold-level intensities.
Behavioral procedures
Training was begun by shaping each animal to peck the response key to obtain access to food. Once an animal consistently pecked the key, the reinforcement rate was incrementally decreased until pecking occurred at a continuous, consistent rate for at least 2 s, at which point the 2 s trial duration was implemented. The final phase of training consisted of training the animal to only respond during silence and withhold their responses during sound presentations. This was accomplished by providing the animal access to reinforcement in only two instances: (1) when the animal responded throughout a 2 s trial in the absence of an acoustic signal, and (2) when the animal withheld its response during the 2 s presentation of an acoustic signal.
An animal’s performance was determined by comparing the proportion of correct responses to sound presentations, hits, to the proportion of incorrect responses during silence, false alarms, through a series of 5–8 sound trials at the same acoustic stimulus intensity. Performance was calculated by correcting the animal’s hit rate based on their false alarm rate during each block of trials based on the following formula: Performance = Hit Rate − (Hit Rate * False Alarm Rate) (Heffner and Heffner 1995). This calculation corrected for the animal’s false alarm rate, resulting in scores ranging from 0 (no hits) to 1.0 (100% hit rate and 0% false alarm rate).
Thresholds were determined by starting each session with the acoustic signal at a clearly audible level, typically 30–35 dB above their suspected threshold, and reducing the signal intensity in 5 dB steps until the animal no longer reliably responded to the signal. Threshold was defined as the intensity corresponding to a corrected performance score of 0.50, which typically needed to be determined by interpolation. Testing was repeated daily at each frequency until thresholds obtained from each animal were stable and at least three sessions were within 3 dB of each other.
Results
Testing began at 1 kHz, progressing in octave steps until the high-frequency limit of the animal’s hearing range was determined, and then systematically assessed the lower frequencies in octave steps starting at 500 Hz until their low-frequency limit was determined. After the full hearing range had initially been determined, retests of multiple frequencies were conducted to ensure the reliability of the original scores. At this time, additional thresholds were determined at intermediate frequencies, 2.8 kHz and 5.6 kHz, to verify the slope of the animal’s high-frequency cutoff. As is shown in Fig. 1 and Table 1, the absolute thresholds of the three quail show close agreement. At an intensity of 60 dB SPL, the mean hearing range extends from 59.5 Hz to 7 kHz, a range of 6.88 octaves. Each of the three animals were observed to have a best sensitivity of 2 kHz, with thresholds ranging from 3.1 to 5.7 dB SPL. As is typical of most bird species, beginning at 5.6 kHz the sensitivity of all three of the animals decreased rapidly to a mean threshold of 84.1 dB SPL at 8 kHz.

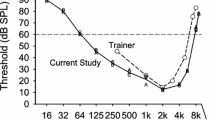
Comparison of Japanese quail thresholds obtained in the current study to those reported by Dooling and Okanoya (1995). SPL is the sound pressure level re 20 μN/m2. Horizontal dashed line denotes the 60 dB sound pressure level cutoff. Note: This figure was generated using SigmaPlot 13
Behavioral thresholds were not able to be determined for animal B at 32 and 16 Hz, or for animal A at 2.8 kHz. Animal B proved to be a difficult test subject from the outset, requiring six additional weeks of training before meeting the criteria to establish its first stable threshold (i.e. a minimum of three sessions within 3 dB of one another). Beginning at 32 Hz, this animal never reliably responded to the acoustic signal above the chance level (i.e. a corrected performance score of 0.5). Retraining using previously tested stimuli did not yield responses above the chance level at 32 or 16 Hz, despite the animal replicating previous thresholds on the retrain stimuli. This may suggest that this animal had a hearing loss at the very low end of his hearing range, though no appreciable differences were observed between this animal’s thresholds and the other birds at any other frequency. Animal A was unable to be tested at 2.8 kHz because this animal perished of coccidiosis before the study concluded.
Discussion
Comparison to previous Japanese quail audiogram
Dooling and Okaonya (1995) previously assessed the auditory sensitivity of the Japanese quail in half-octave steps from 500 Hz to 8 kHz using a two-choice procedure. This two-choice procedure consisted of training birds to peck an observing key to initiate trials and peck a separate reporting key in the presence of a tone. Like the current study, animals were reinforced with brief access to food for correctly reporting signals. However, they were not reinforced for correctly reporting the absence of tones, and a false positive resulted in a brief time out where the chamber lights were turned off. Additionally, the signal intensity for any given presentation was varied randomly between one of seven different intensities in a range of up to 60 dB. The results of the current study are compared to this previous work in Fig. 1. As is evident from Fig. 1, the behavioral thresholds obtained using two different methodologies are quite similar. Both studies found a high-frequency cutoff at 8 kHz and a range of maximal sensitivity in the 2–4 kHz range. These results indicate that the high-frequency hearing and greatest sensitivity of the Japanese quail do not deviate from the typical hearing of most bird species (Dooling 2002).
Comparison to bird low-frequency sensitivity
Prior to this study, the full auditory range and sensitivity have only been determined using psychoacoustic methods for four avian species: the budgerigar (Heffner et al. 2016), domestic chicken (Hill et al. 2014), homing pigeon (Kreithen and Quine 1979; Heffner et al. 2013), and mallard duck (Hill 2017). Figure 2 compares the results of the current study to these four species. The current results closely resemble the typical bird audiogram in two ways. First, it is well known that birds typically do not hear much above 10 kHz (Dooling et al. 2000; Heffner and Heffner 2016). The Japanese quail is no exception to this trend, with a high frequency, upper limit to its hearing range occurring at 8 kHz. Second, the Japanese quail’s frequency of maximal sensitivity occurs at 2 kHz, with all frequencies from 1 to 5.6 kHz having thresholds lower than 20 dB SPL. Again, this closely resembles what has been observed in previous studies, though the peak sensitivity varies from 1 to 5 kHz depending on species (Dooling 2002).

Behavioral thresholds for three animals in the current study compared with the four bird species whose low-frequency sensitivity and range is known. To reference the definition of infrasound (i.e. more sensitive than humans to signals 16 Hz and below) the human audiogram (Jackson et al. 1999) is also included in gray. The Japanese quail’s low-frequency sensitivity does not meet the criteria for infrasound detection. SPL is the sound pressure level re 20 μN/m2. Note: This figure was generated using SigmaPlot 13
The more important outcome of this study is the low-frequency limit of the quail hearing range. Beginning at 64 Hz, and extending to their lower limit of hearing, the sensitivity of those species that detect infrasound begins to diverge from those that do not. While this divergence is not as pronounced as was observed in the chicken and pigeon, the quail do have better sensitivity in this range than the budgerigar and mallard duck. However, the average quail threshold at 16 Hz was 82.7 dB SPL, which is not lower than the human threshold of 82.4 reported by Jackson et al. (1999). Therefore, the Japanese quail does not detect infrasound and these results do not support the hypothesis that infrasound sensitivity is characteristic of all gallinaceous species.
Implications for future research
The purpose of infrasonic sensitivity is not currently understood. To use infrasound as a navigational cue, an animal would need to be able to locate the source of the acoustic signal. However, localizing these signals using binaural cues is unlikely due to the long wavelengths of infrasonic signals, which would result in negligible differences in the acoustic signal arriving at each ear (Quine and Kreithen 1981). The addition of the current study brings the number of birds whose low-frequency hearing range and sensitivity is known to a total of five, of which two detect infrasound. Evaluating these five species there are two identifiable trends emerging from the data that can help guide the selection of future species to further clarify the functional role of this ability.
First, it appears that there is considerable variability in the low-frequency sensitivity of birds. The lowest frequency at which data are available from more than two species is 16 Hz. Among the available data, there is a 35 dB range from the least to the most sensitive species. Although there are only five species represented at this point, the trend resembles that which has been observed in mammals, where some species have good low-frequency sensitivity and others, even those within the same phylogenic group, have poor sensitivity (Heffner and Heffner 2003). This variation suggests that sensitivity to low-frequency sounds may have considerably greater survival valuable for some species than it does for others. Comparative research evaluating similarities and differences between birds and mammals with good low-frequency hearing may yield clues as to why such varation exists in this ability.
Second, the ecological niches occupied by the two species capable of infrasound detection are dramatically different. The homing pigeon, which descended from the rock dove (Columba livia), is thought to use infrasound as a mapping mechanism in navigation (Hagstrum and Manley 2015; Kreithen and Quine 1979). Rocky coastal areas, where the rock dove nests, are known to produce an abundance of infrasonic noise due to water interacting with land (Bedard 1978). Therefore, this signal could be a reliable navigation cue for this species. Chickens, on the other hand, are believed to have descended from several subspecies of the red jungle fowl (Gallus gallus) (Liu et al. 2006), which primarily inhabit areas where densely vegetated forests meet open grasslands. It would not be expected for infrasound to be as readily produced in these areas and coupled with the red jungle fowl’s lack of long-distance flight; this species would not likely use this cue for navigation. This suggests that there may be multiple purposes for sensitivity to very low-frequency sounds, including infrasound detection via other sensory modalities, similar to what has been observed in the peacock (Kane et al 2018). Further research is needed to determine if this ability is the result of convergent evolution, or if it originated from a common ancestor and was repurposed for multiple functions.
Another important consideration for future investigations of low-frequency hearing is whether it is appropriate to classify species dichotomously based on anthropomorphically defined labels (i.e. those that detect infrasound and those that do not). Infrasound is regarded as those frequencies below the lower limit of human hearing, which is defined by the American National Standards Institute (2013) as occurring at 20 Hz. A lower limit of 16 Hz is also frequently reported, as this frequency is in line with the preferred frequencies typically used by acousticians (ISO 1997). However, evolution did not drive the development of this ability based on human hearing. Rather, it is likely more accurate to consider low-frequency hearing as occurring along a continuum. Indeed, it is well established that high-frequency hearing in mammals is driven by head size and the ability to use the interaural intensity cue for localization (Heffner and Heffner 2003), with many species hearing acoustic signals that extend well into the anthropomorphically defined acoustic space of ultrasound. Until such time as the mechanism, or mechanisms, that drive the need for infrasonic sensitivity are determined, we should use caution in categorizing species based on the presence of this ability.
The results of the current study reveal that the Japanese quail does not detect infrasound. While these results do not clarify the functional role of infrasound sensitivity, they do highlight the need for a larger survey of bird low-frequency hearing. Although more than 40 avian species have had their high-frequency hearing assessed, less than 10% of these have been tested below 125 Hz. Some, but not all, of those who have been tested are capable of detecting infrasonic sound. Until such time that a sufficiently large sample of birds have had their low-frequency hearing assessed, the origins and purpose of this ability remain unclear.
References
American National Standard (2013) Bioacoustical terminology. Melville, New York, pp 1–1994
Bedard AJ (1978) Infrasound originating near mountainous regions in Colorado. J Appl Meteorol 17:1014–1022. https://doi.org/10.1175/1520-0450
Corfield JR, Krilow JM, Vande Ligt MN, Iwaniuk AN (2013) A quantitative morphological analysis of the inner ear of galliform birds. Hear Res 304:111–127. https://doi.org/10.1016/j.heares.2013.07.004
Dooling RJ (2002) Avian hearing and the avoidance of wind turbines. National Renewal Energy Laboratory, U.S. Department of Energy. NREL/TP-500–30844. https://doi.org/10.2172/15000693
Dooling RJ, Lohr B, Dent ML (2000) Hearing in birds and reptiles. In: Dooling RJ, Fay RR, Popper AN (eds) Comparative hearing: birds and reptiles. Springer, New York, pp 308–359. https://doi.org/10.1007/978-1-4612-1182-2
Dooling RJ, Okanoya K (1995) The method of constant stimuli in testing auditory sensitivity in small birds. In: Klump GM, Dooling RJ, Fay RR, Stebbins WC (eds) Methods in comparative psychoacoustics. Birkhäuser, Basel, pp 161–169. https://doi.org/10.1007/978-3-0348-7463-2
Freeman AR, Hare JF (2015) Infrasound in mating displays: a peacock’s tale. Anim Behav 102:241–250. https://doi.org/10.1016/j.anbehav.2015.01.029
Garcia M, Charrier I, Rendall D, Iwaniuk AN (2012) Temporal and spectral analyses reveal individual variation in a non-vocal acoustic display: the drumming display of the ruffed grouse (Bonasa umbellus, L.). Ethology 118:292–301. https://doi.org/10.1111/j.1439-0310.2011.02011.x
Hagstrum JT, Manley GA (2015) Release of surgically deafened homing pigeons indicate that aural cues play a significant role in their navigational system. J Comp Physiol A 201:983–1001. https://doi.org/10.1007/s00359-015-1026-3
Heffner HE, Heffner RS (1995) Conditioned avoidance. In: Klump GM, Dooling RJ, Fay RR, Stebbins WC (eds) Methods in comparative psychoacoustics. Birkhäuser, Basel, pp 73–87. https://doi.org/10.1007/978-3-0348-7463-2
Heffner HE, Heffner RS (2003) Audition. In: Davis SF (ed) Handbook of research methods in experimental psychology. Blackwell, Oxford, pp 413–440. https://doi.org/10.1002/9780470756973
Heffner HE, Heffner RS (2016) The evolution of mammalian sound localization. Acoust Today 12:20–35. https://doi.org/10.1063/1.5038516
Heffner HE, Koay G, Hill EM, Heffner RS (2013) Conditioned suppression/avoidance as a method for testing hearing in birds: the domestic pigeon (Columba livia). Behav Res Methods 45:383–392. https://doi.org/10.3758/s13428-012-0269-y
Heffner HE, Koay G, Heffner RS (2016) Budgerigars (Melopsittacus undulatus) do not hear infrasound: the audiogram from 8 Hz to 10 kHz. J Comp Physiol A 202:853–857. https://doi.org/10.1007/s00359-016-1125-9
Heinz GH, Gysel LW (1970) Vocalization behavior of the ring-necked pheasant. Auk 87:279–295. https://doi.org/10.2307/4083920
Hienz RD, Sinnott JM, Sachs MB (1977) Auditory sensitivity of the redwing blackbird and the brown-headed cowbird. J Comp Physiol Psych 91:1365–1376. https://doi.org/10.1037/h0077403
Hill EM (2017) Audiogram of the mallard duck (Anas platyrhynchos) from 16 Hz to 9 kHz. J Comp Physiol A 203:929–934. https://doi.org/10.1007/s00359-017-1204-6
Hill EM, Koay G, Heffner RS, Heffner HE (2014) Audiogram of the chicken (Gallus gallus domesticus) from 2 Hz to 9 kHz. J Comp Physiol A 200:863–870. https://doi.org/10.1007/s00359-014-0929-8
International Organization for Standardization (1997) Acoustics: preferred frequencies for measurements. Switzerland, Geneva
Jackson LL, Heffner RS, Heffner HE (1999) Free-field audiogram of the Japanese macaque (Macaca fuscata). J Acoust Soc Am 106:3017–3023. https://doi.org/10.1121/1.428121
Kane SA, Van Beveren D, Dakin R (2018) Biomechanics of the peafowl’s crest reveals frequencies tuned to social displays. PLoS ONE 13:e0207247. https://doi.org/10.1371/journal.pone.0207247
Kreithen ML, Quine DB (1979) Infrasound detection by the homing pigeon: a behavioral audiogram. J Comp Physiol A 129:1–4. https://doi.org/10.1007/BF00679906
Lieser M, Berthold P, Manley GA (2005) Infrasound in the capercaillie (Tetrao urogallus). J Ornithol 146:395–398. https://doi.org/10.1007/s10336-005-0003-y
Liu YP, Wu GS, Yao YG, Miao YW, Luikart G, Baig M, Beja-Pereira A, Ding ZL, Palanichamy MG, Zhang YP (2006) Multiple maternal origins of chickens: out of the Asian jungles. Mol Phylogenet Evol 38:12–19. https://doi.org/10.1016/j.ympev.2005.09.014
Quine DB (1982) Infrasounds: a potential navigational cue for homing pigeons. In: Papi F, Wallraff HG (eds) Avian navigation. Springer, Berlin, pp 373–376. https://doi.org/10.1007/978-3-642-68616-0
Quine DB, Kreithen ML (1981) Frequency shift discrimination: can homing pigeons locate infrasounds by Doppler shifts? J Comp Physiol A 141:153–155. https://doi.org/10.1007/BF01342661
Schwartzkopff J (1949) Über Sitz und Leistung von Gehör und Vibrationssinn bei Vögeln. Z Vergl Physiol 31:527–603. https://doi.org/10.1007/BF00348361
Theurich M, Langner G, Scheich H (1984) Infrasound responses in the midbrain of the guinea fowl. Neurosci Lett 49:81–86. https://doi.org/10.1016/0304-3940(84)90140-X
Acknowledgements
Funding for the animals, equipment, and student workers were provided by the University of Nebraska-Kearney Undergraduate Research Fellows Program. Additionally, we thank Henry Heffner for his comments and suggestions on an early draft of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Animal welfare
All animals were cared for following the Guide for the Care and Use of Animals. All procedures were conducted in accordance with Animal Care Protocol #041116, which was approved by the by the University of Nebraska-Kearney Animal Care and Use Committee.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Strawn, S.N., Hill, E.M. Japanese quail (Coturnix japonica) audiogram from 16 Hz to 8 kHz. J Comp Physiol A 206, 665–670 (2020). https://doi.org/10.1007/s00359-020-01428-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00359-020-01428-4