
Abstract
Stomach contents were identified from 206 Antarctic starry skate (Amblyraja georgiana) that were collected during three groundfish surveys (September 2007, April 2008 and January 2009) at South Georgia, Southern Ocean. The diet of A. georgiana varied with skate size and between years. Preferred prey included fish (particularly for larger individuals) and Antarctic krill, Euphausia superba, as well as amphipods, polychaetes and other benthic fauna. The skate A. georgiana appears to be an opportunistic predator, and the clear presence of Antarctic krill in this demersal predator’s diet may indicate a benthic habit of this euphausiid species, which has hitherto mainly been considered as occupying a purely pelagic niche.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Studying fish feeding ecology reveals interactions between predators and prey and hence helps to build an understanding of larger scale relationships and ecosystem functioning. In the sub-Antarctic, where climate variability can have profound effects on interannual abundance of key species such as Antarctic krill, Euphausia superba, Dana (Murphy et al. 1998; Atkinson et al. 2004), this causes the diet of certain fish predators to vary considerably between years (Main et al. 2009).
The importance of krill in the pelagic ecosystem of the Southern Ocean is well known. For example, fur seals Arctocephalus gazella Peters, gentoo penguins, Pygoscelis papua Forster and mackerel icefish, Champsocephalus gunnari Lönnberg all feed predominantly on krill. In “krill poor” years, when advection of this species to the waters surrounding South Georgia is reduced (see Brierley et al. 2002), these pelagic, krill-dependent predators may suffer from poor condition, reduced breeding success or mortality (Kock et al. 1994; Murphy et al. 1998, 2007; Everson et al. 1999, 2000; Main et al. 2009). Studies on diet in the sub-Antarctic have been mainly on these pelagic predators, and data on fish predators with a more benthic habit (see Collins et al. 2007) are less frequent.
Skates (Rajidae) are a phylogenetically diverse group of chondrichthyan fish (McEachran and Dunn 1998) that are distributed worldwide in many marine benthic habitats. As important predators, skates may exert considerable influence on marine demersal trophic dynamics (Ebert and Bizzarro 2007), by preying on a wide range of organisms including benthic polychaetes and crustacea, cephalopods and other fish. Skate diet may vary considerably with sex (Gray et al. 1997; San Martin et al. 2007); body size (Gray et al. 1997; Koen Alonso et al. 2001); foraging depth (Bizzarro et al. 2007; Robinson et al. 2007); habitat (Treloar et al. 2007); maturity or season (Rinewalt et al. 2007).
The Antarctic starry skate Amblyraja georgiana (Norman) is distributed widely on the shelves around the sub-Antarctic island of South Georgia, SW Atlantic and also on the shelves of Patagonia, New Zealand and of Scott Island in the Ross Sea. With an approximate size range of 15–115 cm total length (TL), A. georgiana is found over a relatively large depth range on the shelf around South Georgia. This brings them into contact with a commercial longline fishery for toothfish (Dissostichus eleginoides Smitt), which operates around South Georgia in depths >500 m and in which A. georgiana is caught as bycatch. The South Georgia toothfish fishery is managed under the auspices of the Commission for the Conservation of Antarctic Marine Living Resources (CCAMLR), which advocates an ecosystem approach to fisheries management. The ecosystem approach requires a better understanding of the role of bycatch species such as A. georgiana in the South Georgia ecosystem and, furthermore, a requirement to establish how vulnerable this species is to mortality potentially caused by the toothfish fishery. Recognising the need for improving the knowledge of the biology and ecology of rajids, CCAMLR designated the 2008–2009 season as “Year of the Skate” to encourage research on this important component of bycatch. The biology and ecology of A. georgiana is poorly understood, and a detailed analysis of this predator’s diet is therefore timely.
Here, we will examine the diet of Antarctic starry skate caught at South Georgia during two winter surveys (2007, 2008) and one summer survey (2009). Our aims are to describe the diet of A. georgiana and to investigate ontogenetic shifts in diet and differences between the different years from which data were collected. The role of A. georgiana as a benthic predator in the South Georgia ecosystem will be discussed in relation to its preferred prey.
Methods
Sampling locations and collection
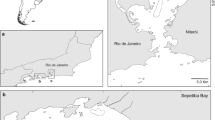
Animals were collected from winter and summer bottom trawl surveys on the South Georgia shelf. The surveys were undertaken on the FPV Dorada (September 2007) and the FV Sil (April 2008 and January 2009) in the area of South Georgia and Shag Rocks (Fig. 1). The area sampled is situated between the Polar Front and the Southern Antarctic Circumpolar Current Front, downstream of the Antarctic Peninsula in the Antarctic Circumpolar Current (Murphy et al. 2007).

Map of sampling locations from the three surveys showing the number of skate caught at each location. Contour lines represent depths of 200 m (dotted) and 500 m (solid)
All survey samples were collected using a commercial-sized otter trawl net (FP-120; see Everson et al. 1999) that was fished for 30 min on the sea floor at a speed of 3–4 knots with a headline height of 4–6 m, a wingspread of approximately 18 m and with a cod end mesh size of 40 mm. With the exception of four trawls in April 2008 and two in January 2009, all trawling took place in daylight.
Stomachs were removed from the majority of captured skate and immediately frozen at −20°C. At the time of removal, stomach fullness was recorded on a scale of 0–4, with 0 being empty, 1: <25% full; 2: 25–50% full; 3: 51–75% full and 4: >75% full (Table 1). Stomach fullness was judged subjectively in relation to the respective specimen’s body size. Biological data including total length (nearest cm below), disc width, total wet weight (nearest gram), sex and maturity stage were recorded for each animal.
Laboratory processing
In the laboratory, stomachs were thawed, total contents were weighed and were then sorted into species or species groups. Prey items were identified to the lowest taxonomic level possible using published guides (Gon and Heemstra 1990; Boltovskoy 1999) and reference collections. Partially digested fish were identified where possible from sagittal otoliths, using reference material and published guides (Hecht 1987; Reid 1996). Sorted contents were counted and weighed (to the nearest 0.01 g). The amount of digestion that prey items had undergone, the “digestive state,” was recorded on a scale from 1 to 4, judged subjectively. Total length (TL) of individual prey items was measured where digestion permitted i.e. where the body was complete and was not more digested than level 2 on the aforementioned scale. Lengths of krill were taken from the front of the eye to the tip of the telson. Any fresh prey items that had visibly undergone no digestion were thought to constitute net feeding and were excluded from the data.
Diet analysis
Precision of diet description
Cumulative prey curves were plotted to indicate whether there were enough stomachs in each sample to describe the diet of A. georgiana adequately and therefore precisely (Ferry and Cailliet 1996).
Diet composition
The relative importance of different dietary components was expressed using the measures of per cent numbers (%N), per cent mass (%M) and per cent frequency of occurrence (%F). A combined index of relative importance (IRI) of prey (Pinkas et al. 1971; Cortes 1997) was calculated using the equation given below.
Per cent index of relative importance:
where N is numbers, M is mass based on the wet weight of the prey found in the stomach and not on reconstituted mass and F is frequency of occurrence. The % IRI of each individual component of the diet was calculated from the formula above (Table 2, provided as Electronic Supplementary Material, see Online Resource 1).
Several comparisons of sample groups using diet categories were carried out, using bootstrapping (see Efron and Tibshirani 1993) to calculate 95% confidence intervals for the % IRI based on pre-defined diet categories (%IRIDC) (San Martin et al. 2007; Main et al. 2009). The allocation of species and species groups to diet categories for this analysis was as follows. Antarctic krill and other euphausiids (EUP); fish (FSH); mysids (MYS); polychaetes (POL); amphipods (AMP); crustacea “not elsewhere included” (CNEI); others (OTH includes cephalopods, salps and unidentified diet components). Differences between values of mean %IRIDC were considered to be significant when there was no overlap in 95% confidence intervals.
Other analyses included a brief investigation into prey size versus predator size and a closer look at aspects of the more important dietary components.
Results
Size, maturity and distribution of Amblyraja georgiana
Amblyraja georgiana were caught on the shelf around South Georgia and Shag Rocks, in depths ranging from approximately 115 to 346 m (Fig. 1). Larger catches tended to be taken around the northern end of South Georgia and at Shag Rocks to the west. In all 3 years, very few skate were caught in the south-east. The largest sample of skate was taken in the 2007 survey, to the north of South Georgia in ~280 m of water. The smallest overall catch of skate (less than half in number of individuals) was taken in 2009, despite a similar amount of effort being used in this survey relative to the two others (the same vessel was used for the 2008 and 2009 surveys, although a different vessel was used for the 2007 survey. The same net was used for all trawling).
Specimens of A. georgiana ranged in size from 16 to 101 cm TL (Fig. 2). The length frequency of all skate caught was used to define four size groups of “recently hatched,” “small,” “medium” and “large” skate, for which diet was compared (using %IRIDC) to investigate possible ontogenetic shifts in diet. These size groups were defined as: <28 cm; 28–50 cm; 51–74 cm and >74 cm TL.

Length frequency of skate caught in the three surveys. Divisions indicate the size classes used in subsequent diet comparisons (see Fig. 5 and text for details)
The number of males and females collected during each survey was approximately equal. In each survey, more than 60% of the skate were immature as measured on a six-point subjective scale used by CCAMLR observers deployed on toothfish fishing vessels in the region and also in use in the Falkland Islands (Arkhipkin et al. 2008) (Table 1).
Diet composition
The proportion of empty skate stomachs was low (<15% in each survey, Table 1). The number of different types of prey present in a stomach ranged from 0 to 9 with a mean of 3 types in 2007 and 2008 and 2 types in 2009.
Asymptotes approached by cumulative prey curves plotted for the 2007 and 2008 surveys (Fig. 3a, b) indicated that these sample sizes of 84 and 89 stomachs were adequate to describe the diet of A. georgiana. This evidence was not seen as clearly for the 2009 survey, where the sample size was about half the number of stomachs (Fig 3c). However, the diet generally seemed much less diverse in that year and was dominated by Antarctic krill.

Cumulative prey curves showing the average number of new species found with increasing number of stomach samples. Each analysis was randomised ten times. Error bars show the standard deviation of the mean result
Diet varied between the 3 years. The predominant prey category taken by A. georgiana was fish in 2007 and euphausiids (mainly Antarctic krill, Euphausia superba) in both 2008 and 2009 (see Fig. 4 and Table 2: Online Resource 1). Amphipods, mysids and other crustacea were taken in smaller proportions. Several cephalopod beaks were found in the stomachs, including the octopods, Thaumeledone gunteri Robson and Pareledone turqueti (Joubin). Beaks from the squid, Kondakovia longimana Filippova and Slosarczykovia circumantarctica Lipinski were also found. There were no clear patterns in stomach fullness, either between years or with time of day.

Diet comparisons (%IRIDC) between years. AMP Amphipod, CNEI Crustacea not elsewhere included, EUP Euphausiid (mainly Antarctic krill), FSH Fish, MYS Mysid, POL Polychaete and OTH Others
Ontogenetic shifts in diet were evident, with smaller animals taking more amphipods and polychaetes and larger animals switching to prey more on fish (Fig. 5). Animals of all sizes had preyed on euphausiids, although the largest skate had done so less than smaller individuals.

Diet comparisons (%IRIDC) between defined size classes. AMP Amphipod, CNEI Crustacea not elsewhere included, EUP Euphausiid (mainly Antarctic krill), FSH Fish, MYS Mysid, POL Polychaete and OTH Others
Total lengths of a subset of prey (~25% of all prey individuals) were taken, with only prey that had not undergone significant digestion included. There was no clear relationship between prey size and predator size (Fig. 6). Examining the data by diet category (see section on % IRI calculations) showed that apart from the fish prey category, the prey had a fairly narrow size range. Diet categories “Others” and “Polychaetes” were not included because the polychaetes were invariably in a digestive state that precluded accurate measurement, and only two prey from the others category were measured (TL). The analysis was carried out on prey that were measured and in a digestive state of 2 or less on a 4-point subjective scale. The mean length of krill was 35 mm in 2007; 46 mm in 2008 and 51 mm in 2009.

Prey–predator size relationships in the diet of Amblyraja georgiana. NEI = not elsewhere included
Discussion
The Antarctic starry skate, Amblyraja georgiana, has a diverse diet, which includes benthic polychaetes and crustacea, fish, cephalopods and Antarctic krill. In 2007, the main component of diet was fish, mainly the notothenid Gobionotothen gibberifrons (Lönnberg), whereas in the subsequent 2 years, euphausiids, mainly Antarctic krill, were the main prey. Our findings suggest that this skate is an opportunistic predator and is probably able to adapt its diet according to preference or alternatively to what is most readily available as prey. The most common fish prey, G. gibberifrons, is a benthic species that is widely distributed on the shelves of South Georgia and the South Sandwich Islands, South Shetland, South Orkneys and Heard islands and also the shelf of the Antarctic Peninsula (Gon and Heemstra 1990). As well as being important prey for A. georgiana, G. gibberifrons also features in the diet of other demersal predators at South Georgia, such as Patagonian toothfish (Collins et al. 2007).
There were clear ontogenetic shifts in the diet of A. georgiana, as indicated by the comparison of the combined index of relative importance for diet categories defined a priori (%IRIDC). A balanced description of diet benefits from the use of several measures (Hyslop 1980), and a combined index (IRI, Pinkas et al. 1971) may reduce bias in the interpretation of dietary importance (Bigg and Perez 1985; Liao et al. 2001). Despite some issues noted with the use of this index (Hansson 1998; Tirasin and Jorgensen 1999), bootstrapping used to calculate confidence intervals can help to enable statistical comparisons between sample groups of a study (Braccini et al. 2005; Main et al. 2009) or related studies (Collins and Main 2009). Diet composition of skate grouped in size classes showed that smaller (younger) skate took proportionately more amphipods and polychaetes than larger (older) skate, which had preyed more on other fish. Occurence of this kind of dietary shift, with larger individuals exhibiting more piscivorous habits, has also been found in other skate species (e.g. Brickle et al. 2003; Bizzarro et al. 2007). For example, two diet shifts were described for the beaked skate Dipturus chilensis (Guichenot) in the Atlantic off Patagonia, in which the addition of fishes and molluscs to the diet was found at around 35 cm TL, followed by a reduction in dietary importance of crustaceans at around 85 cm TL (Garcia de la Rosa et al. 2004).
The feeding strategies adopted by skates vary, with some evidence showing that certain skate species are benthic scavengers. For example, stomachs from the spotted ray, Raja montagui Fowler, contained fish remains that were discarded from a vessel directly before trawl sampling (Olaso et al. 2002). In contrast, Orlov (1998 and references therein) presented an argument that skates are also able to hunt for pelagic prey, stating that pelagic fish may be an important component of skate diet in various ocean regions. All of the size classes of skate in the present study had fed on Antarctic krill, Euphausia superba. The clear importance of Antarctic krill in the diet of A. georgiana was of particular interest in relation to the view that Antarctic krill tend to occupy a pelagic niche in the top ~150 m of the water column (Marr 1962). This has since been challenged by observations of krill actively feeding close to the seabed on the continental shelf (Gutt and Siegel 1994) and at abyssal depths (Clarke and Tyler 2008). The position of a skate’s mouth, being situated on its ventral surface, indicates that these animals are well adapted for benthic feeding. Other pelagic species were found in the diet of A. georgiana, e.g. the amphipod, Themisto gaudichaudii, and the myctophid fish, Protomyctophum bolini, but these occurred in small numbers and could have been secondarily ingested. There is little doubt that krill were directly consumed by A. georgiana, as they were sometimes the only prey and were often relatively undigested. We therefore surmise that A. georgiana did not scavenge on dead or moribund krill, instead taking them live and probably at the seabed. Our findings hence indicate the importance of further study of Antarctic krill behaviour and how it may contribute to benthic–pelagic coupling.
There were appreciable differences in diet between the 3 years studied. The proportion of krill in the diet was low (15% IRIDC) in 2007 and high (>70% IRIDC) in both 2008 and 2009. Acoustic surveys routinely used to provide an index of krill abundance in the Scotia Sea (see Brierley et al. 1998, 1999) indicated that krill was abundant at South Georgia in 2008, although multiple indicators demonstrated low krill availability in 2009 (Hill et al. 2009). The high proportion of euphausiids in skate diet in 2009 is in contrast to the poor feeding success of krill-dependent mackerel icefish, Champsocephalus gunnari, feeding in the same areas and water depths during the same period (Collins and Main 2009). We consider that behavioural adaptations of skate, as predators accustomed to feeding at the seabed, may have allowed these animals to continue to feed on krill during this period of generally low krill availability in 2009; while the krill-dependent pelagic fish, Champsocephalus gunnari, which are thought to make vertical migrations into the water column in order to feed, had found less krill. Of further interest is the mean size of krill in the skate diet. Of the 3 years studied, the largest mean size of krill present in the stomachs of A. georgiana was in 2009 (51 mm TL in 2009 compared with 35 mm in 2007 and 46 mm in 2008). Although krill spawn both on and off the shelf at South Georgia, local recruitment by surviving larvae is low, resulting in relatively large mean size of krill in the area. The presence of krill at South Georgia is therefore mainly a result of advection (Hofmann and Murphy 2004; Tarling et al. 2007). As stated, multiple indicators showed that 2009 was a poor year for krill at South Georgia (Collins and Main 2009; Hill et al. 2009) and the relatively large mean length of krill in skate stomachs in January 2009 (51 mm) supports this, since it is probable that in times of reduced advection of krill to South Georgia, local recruitment would fail and only krill that had already grown to a larger size would survive and remain.
Previous descriptions of skate diet are mainly from temperate waters. The present study collected samples from sub-Antarctic waters mainly during daylight and from three separate seasons. The diet appeared to be much less diverse in the summer (2009) survey than in the previous two surveys (Fig. 3c). It is not clear whether this was seasonally driven or could have been related to the ecosystem anomaly reported for the region in 2009 (Hill et al. 2009). More data would be required in order to provide a comparison between seasonal diets of the Antarctic starry skate. The ability of A. georgiana to find and prey on krill during times when krill-dependent predators had not done so is perhaps surprising and needs further study. This is important in assessing the ability of fish populations to survive through periods of fluctuating krill availability and to give better insight into the ecology and behaviour of Antarctic krill.
References
Arkhipkin AI, Baumgartner N, Brickle P, Laptikhovsky VV, Pompert JHW, Shcherbich ZN (2008) Biology of the skates Bathyraja brachyurops and B. griseocauda in waters around the Falkland Islands, Southwest Atlantic. ICES J Mar Sci 65:560–570
Atkinson A, Siegel V, Pakhomov E, Rothery P (2004) Long-term decline in krill stock and increase in salps within the Southern Ocean. Nature 432:100–103
Bigg MA, Perez MA (1985) Modified volume: a frequency-volume method to assess marine mammal food habits. In: Beddington JR, Beverton RJH, Lavigne DM (eds) Marine mammals and fisheries. George Allen & Unwin, London, pp 277–283
Bizzarro JJ, Robinson HJ, Rinewalt CS, Ebert DA (2007) Comparative feeding ecology of four sympatric skate species off central California, USA. Environ Biol Fish 80:197–220. doi:10.1007/s10641-007-9241-6
Boltovskoy D (ed) (1999) South Atlantic zooplankton, vol 1. Backhuys Publishers, Leiden
Braccini JM, Gillanders BM, Walker TI (2005) Sources of variation in the feeding ecology of the piked spurdog (Squalus megalops): implications for inferring predator-prey interactions from overall dietary composition. ICES J Mar Sci 62:1076–1094. doi:10.1016/j.icesjms.2005.04.004
Brickle P, Laptikhovsky V, Pompert J, Bishop A (2003) Ontogenetic changes in the feeding habits and dietary overlap between three abundant rajid species on the Falkland Islands’ shelf. J Mar Biol Assoc UK 83:1119–1125
Brierley AS, Ward P, Watkins JL, Goss C (1998) Acoustic discrimination of Southern Ocean zooplankton. Deep-Sea Res Part II-Top Stud Oceanogr 45:1155–1173
Brierley AS, Watkins JL, Goss C, Wilkinson MT, Everson I (1999) Acoustic estimates of krill density at South Georgia, 1981–1998. CCAMLR Sci 6:47–57
Brierley AS, Goss C, Grant SA, Watkins JL, Reid K, Belchier M, Everson I, Jessop MJ, Afanasyev V, Robst J (2002) Significant intra-annual variability in krill distribution and abundance at South Georgia revealed by multiple acoustic surveys during 2000/01. CCAMLR Sci 9:71–82
Clarke A, Tyler PA (2008) Adult antarctic krill feeding at abyssal depths. Curr Biol 18:282–285. doi:10.1016/j.cub.2008.01.059
Collins MA, Main CE (2009) Spatial patterns in mackerel icefish diet provides insights into krill abundance and distribution. WG-EMM-09/27. Working Group on Ecosystem Monitoring and Management, CCAMLR, Bergen
Collins MA, Ross KA, Belchier M, Reid K (2007) Distribution and diet of juvenile Patagonian toothfish on the South Georgia and Shag Rocks shelves (Southern Ocean). Mar Biol 152:135–147. doi:10.1007/s00227-007-0667-3
Cortes E (1997) A critical review of methods of studying fish feeding based on analysis of stomach contents: application to elasmobranch fishes. Can J Fish Aquat Sci 54:726–738
Ebert DA, Bizzarro JJ (2007) Standardized diet compositions and trophic levels of skates (Chondrichthyes: Rajiformes: Rajoidei). Environ Biol Fish 80:221–237. doi:10.1007/s10641-007-9227-4
Efron B, Tibshirani RJ (1993) An introduction to the bootstrap. Chapman & Hall, New York
Everson I, Parkes G, Kock KH, Boyd IL (1999) Variation in standing stock of the mackerel icefish Champsocephalus gunnari at South Georgia. J Appl Ecol 36:591–603
Everson I, Kock KH, Ellison J (2000) Inter-annual variation in the gonad cycle of the mackerel icefish. J Fish Biol 57:103–111
Ferry LA, Cailliet GM (1996) Sample size and data analysis: are we characterizing and comparing diet properly? In: Gutshop ‘96. Feeding ecology and nutrition in fish. American Fisheries Society Symposium. Physiology Section, pp 71–80
Garcia de la Rosa SB, Sanchez F, Prenski B (2004) Biological characterization and exploitation state of skate (Dipturus chilensis). In: Sanchez RP, Bezzi SI (eds) The Argentine Sea and its fisheries resources. Vol 4. Marine fishes of commercial interest. Universidad Nacional de Mar del Plata, Mar del Plata, pp 53–66
Gon O, Heemstra PC (1990) Fishes of the Southern Ocean, First edn. South African Institute for Aquatic Biodiversity, Grahamstown
Gray AE, Mulligan TJ, Hannah RW (1997) Food habits, occurrence, and population structure of the bat ray, Myliobatis californica, in Humbolt Bay, California. Environ Biol Fish 49:227–238
Gutt J, Siegel V (1994) Benthopelagic aggregations of krill (Euphausia superba) on the deeper shelf of the Weddell Sea (Antarctic). Deep Sea Res (I Oceanogr Res Pap) 41:169–178
Hansson S (1998) Methods of studying fish feeding: a comment. Can J Fish Aquat Sci 55:2706–2707. doi:10.1139/cjfas-55-12-2706
Hecht T (1987) A guide to otoliths of Southern Ocean fishes. S Afr J Antarct Res 17:1–87
Hill SL, Waluda CM, Fielding S, Belchier M, Venables HJ, Murphy EJ, Collins MA, Agnew DA, Trathan P (2009) Ecosystem indicators of an extreme anomaly at South Georgia in 2009. WG-EMM-09. Working Group on Ecosystem Monitoring and Management, CCAMLR, Bergen
Hofmann EE, Murphy EJ (2004) Advection, krill and Antarctic marine ecosystems. Antarct Sci 16:487–499. doi:10.1017/so954102004002275
Hyslop EJ (1980) Stomach contents analysis–a review of methods and their application. J Fish Biol 17:411–429
Kock KH, Wilhelms S, Everson I, Groger J (1994) Variations in the diet composition and feeding intensity of mackerel icefish Champsocephalus gunnari at South Georgia (Antarctic). Mar Ecol Prog Ser 108:43–57
Koen Alonso M, Crespo EA, Garca NA, Pedraza SN, Mariotti PA, Bern Vera B, Mora NJ (2001) Food habits of Dipturus chilensis (Pisces: Rajidae) off Patagonia, Argentina. ICES J Mar Sci 58: 288–297
Liao H, Pierce CL, Larscheid JG (2001) Empirical assessment of indices of prey importance in the diets of predacious fish. Trans Am Fish Soc 130:583–591
Main CE, Collins MA, Mitchell R, Belchier M (2009) Identifying patterns in the diet of mackerel icefish (Champsocephalus gunnari) at South Georgia using bootstrapped confidence intervals of a dietary index. Polar Biol 32:569–581
Marr JWS (1962) The natural history and geography of the Antarctic krill (Euphausia superba, Dana). Discovery Reports, vol 32
McEachran JD, Dunn KA (1998) Phylogenetic analysis of skates, a morphologically conservative clade of elasmobranchs (Chondrichthyes: Rajidae). Copeia 2:271–290
Murphy EJ, Watkins JL, Reid K, Trathan PN, Everson I, Croxall JP, Priddle J, Brandon MA, Brierley AS, Hofmann E (1998) Interannual variability of the South Georgia marine ecosystem: biological and physical sources of variation in the abundance of krill. Fish Oceanogr 7:381–390
Murphy EJ, Watkins JL, Trathan PN, Reid K, Meredith MP, Thorpe SE, Johnston NM, Clarke A, Tarling GA, Collins MA, Forcada J, Shreeve RS, Atkinson A, Korb R, Whitehouse MJ, Ward P, Rodhouse PG, Enderlein P, Hirst AG, Martin AR, Hill SL, Staniland IJ, Pond DW, Briggs DR, Cunningham NJ, Fleming AH (2007) Spatial and temporal operation of the Scotia Sea ecosystem: a review of large-scale links in a krill centred food web. Philos Trans R Soc Lond, Ser B: Biol Sci 362:113–148. doi:10.1098/rstb.2006.1957
Olaso I, Sanchez F, Rodriguez-Cabello C, Velasco F (2002) The feeding behaviour of some demersal fish species in response to artificial discarding. Sci Mar (Barc) 66:301–311
Pinkas L, Oliphant MS, Iverson ILK (1971) Food habits of albacore, bluefin tuna, and bonito in California waters (1971). Calif Fish Game 152:1–105
Reid K (1996) A guide to the use of otoliths in the study of predators at South Georgia. British Antarctic Survey, Cambridge
Rinewalt CS, Ebert DA, Cailliet GM (2007) Food habits of the sandpaper skate, Bathyraja kincaidii (Garman, 1908) off central California: seasonal variation in diet linked to oceanographic conditions. Environ Biol Fish 80:147–163. doi:10.1007/s10641-007-9218-5
Robinson HJ, Cailliet GM, Ebert DA (2007) Food habits of the longnose skate, Raja rhina (Jordan and Gilbert, 1880), in central California waters. Environ Biol Fish 80:165–179
San Martin MJ, Braccini JM, Tamini LL, Chiaramonte GE, Perez JE (2007) Temporal and sexual effects in the feeding ecology of the marbled sand skate Psammobatis bergi Marini, 1932. Mar Biol 151(2):505–513. doi:10.1007/s00227-006-0499-6
Tarling GA, Cuzin-Roudy J, Thorpe S, Shreeve R, Ward P, Murphy EJ (2007) Recruitment of Antarctic krill Euphausia superba in the South Georgia region: adult fecundity and the fate of larvae. Mar Ecol Prog Ser 331:161–179
Tirasin EM, Jorgensen T (1999) An evaluation of the precision of diet description. Mar Ecol Prog Ser 182:243–252
Treloar MA, Laurenson LJB, Stevens JD (2007) Dietary comparisons of six skate species (Rajidae) in south-eastern Australian waters. Environ Biol Fish 80:181–196. doi:10.1007/s10641-007-9233-6
Acknowledgments
We would like to thank the scientists and crew working on the three surveys and Helena Wiklund and Mike Thurston for help with the identification of polychaetes and amphipods, respectively. Thanks also to Katrin Schmidt for some constructive comments on the manuscript and Alex Brown for some help with graphics. This work contributed to the British Antarctic Survey Discovery 2010 project.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Main, C.E., Collins, M.A. Diet of the Antarctic starry skate Amblyraja georgiana (Rajidae, Chondrichthyes) at South Georgia (Southern Ocean). Polar Biol 34, 389–396 (2011). https://doi.org/10.1007/s00300-010-0894-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-010-0894-9