
Abstract
The distribution and diet of juvenile (<750 mm) Patagonian toothfish are described from four annual trawl surveys (2003–2006) around the island of South Georgia in the Atlantic sector of the Southern Ocean. Recruitment of toothfish varies inter-annually, and a single large cohort dominated during the four years surveyed. Most juveniles were caught on the Shag Rocks shelf to the NW of South Georgia, with fish subsequently dispersing to deeper water around both the South Georgia and Shag Rocks shelves. Mean size of juvenile toothfish increased with depth of capture. Stomach contents analysis was conducted on 795 fish that contained food remains and revealed that juvenile toothfish are essentially piscivorous, with the diet dominated by notothenid fish. The yellow-finned notothen, Patagonotothen guntheri, was the dominant prey at Shag Rocks whilst at South Georgia, where P. guntheri is absent, the dominant prey were Antarctic krill and notothenid fish. The diet changed with size, with an increase in myctophid fish and krill as toothfish grow and disperse. The size of prey also increased with fish size, with a greater range of prey sizes consumed by larger fish.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Patagonian toothfish (Dissostichus eleginoides) belongs to the notothenioids or Antarctic cods that are endemic to the southern hemisphere and dominate Antarctic fish assemblages (Kock 1992). It is circumpolar in distribution, being found around sub-Antarctic islands such as South Georgia, Heard Island and Kerguelen Island and also extends north onto the Patagonian shelf. To the south it is replaced by the congeneric Antarctic toothfish (Dissostichus mawsoni), which is found at high latitudes around the Antarctic continent (Gon and Heemstra 1990).
Patagonian toothfish reach large size (>2 m, >100 kg) and are long lived with adult fish believed to reach 50 years old (Horn 2002). Growth is relatively quick for the first 10 years, while the fish inhabit relatively shallow water, but following the onset of maturity [700–800 mm total length (TL)] growth is very slow. Spawning is thought to occur in deep-water, with both eggs and larvae pelagic (Evseenko et al. 1995).
The large size of toothfish, coupled with high quality flesh, led to the development, in the mid 1980s, of a valuable long-line fishery, targeting large adult fish in deep water (>500 m) (Agnew 2004). The fishery began in Chilean waters, but rapidly expanded to cover the geographic range of toothfish (Agnew 2004). At South Georgia the long-line fishery began in 1988, targeting large adult fish in deep-water although toothfish had previously been taken in bottom trawls on the shelf. Since the mid 1990s the fishery has been managed under the auspices of the Commission for the Conservation of Antarctic Marine Living Resources (CCAMLR), with mean annual catches of around 4,000 tonnes (Agnew 2004).
Ecologically sustainable management of a fishery requires an understanding of the distribution and ecology of the exploited species throughout the life cycle and interactions with other species in the ecosystem, and this underpins CCAMLR’s ecosystem approach to fisheries management (Constable et al. 2000). Whilst the distribution and ecology of the adult part of the toothfish population has been elucidated through the fishery (e.g. Pilling et al. 2001; Agnew 2004) and the use of baited cameras (Yau et al. 2002; Collins et al. 2006), the distribution and ecology of the pre-recruits or juveniles is poorly documented.
The diet of Patagonian toothfish has been studied across its geographic range (Duhamel 1981; McKenna 1991; Garcia de la Rosa et al. 1997; Goldsworthy et al. 2001, 2002; Pilling et al. 2001; Arkhipkin et al. 2003; Barrera-Oro et al. 2005), but most of these studies have focused on adult toothfish from the fishery. At South Georgia adult toothfish (>750 mm TL) are thought to be opportunistic predators and scavengers (Pilling et al. 2001), but data on the diet of fish prior to entering the fishery (TL ∼750 mm) is limited to a study from a single survey in March–April 1996 (Barrera-Oro et al. 2005).
Here we examine the distribution and diet of pre-recruit toothfish from trawl surveys undertaken at South Georgia and Shag Rocks in four consecutive seasons and consider the role of toothfish in the South Georgia ecosystem.
Materials and methods
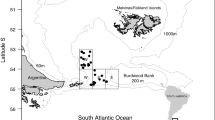
During January 2003, 2004, 2005 and 2006 bottom trawl surveys were undertaken on the FPRV Dorada in the area of South Georgia and Shag Rocks. South Georgia is situated between the Polar Front and the Southern Antarctic Circumpolar Current Front (SACCF), with circulation generally flowing from west to east in the Antarctic Circumpolar Current (see Fig. 1). The surveys used a commercial sized otter trawl (FP-120), which was fished, during daylight, for approximately 30 min at each station. The trawl had a headline height of 4 m, and fished with a wingspread of 18–20 m and a cod-end mesh of 40 mm. In January 2003 trawl stations were arranged in a series of transects radiating away from the island and covering depths of 100–900 m. In 2004–2006 the trawl stations were arranged in a random, stratified design, to assess the abundance of mackerel icefish and pre-recruit toothfish.

The location of South Georgia and Shag Rocks in relation to the main fronts and currents in the Scotia Sea. SACCF Southern Antarctic Circumpolar Current Front, SB Southern Boundary of Antarctic Circumpolar Current
During all surveys, all captured toothfish were sampled, and were measured (to 10 mm category below), weighed and sexed. Except in 2003, when a subsample was taken, all toothfish stomachs that contained any food items were carefully dissected from the fish, and immediately frozen at −20°C.
In the laboratory stomachs were thawed and the total contents weighed prior to being sorted into species or species groups. Contents were identified to the lowest taxonomic level using published guides (Hulley 1981; Nesis 1987; Gon and Heemstra 1990) and reference collections. Partially digested fish were identified from sagital otoliths using reference material and published guides (Hecht 1987; Reid 1996). Partially digested cephalopods were identified using reference collections of beaks. Individual prey items were weighed and measured (TL for fish; ML for cephalopods), with the size of highly digested prey estimated from otolith to length relationships (Hecht 1987; Reid 1996). Items that were completely undigested were considered to represent trawl feeding and were excluded from subsequent analyses.
Diet was expressed using percent mass (% M), percent frequency of occurrence (% F), percent number (% N) and percent index of relative importance (% IRI: see Cortes 1996). Percent mass was based on the weight of the prey found in the stomach and not on reconstituted mass.
S = number of stomachs containing food remains
Statistical analyses were undertaken using the statistical software MINITAB Release 14 and Sigma Plot 9.01. Inter-annual comparisons of stomach fullness (expressed as % body weight) were investigated using a one-way ANOVA on arcsin-transformed data. Regression analyses were undertaken to investigate the relationships between depth (independent) and fish length and between toothfish size (independent) and prey size. Assumptions about normality and constant variance were tested prior to analyses.
Results
Distribution of juvenile toothfish
Juvenile Patagonian toothfish (<750 mm TL) were found throughout the South Georgia and Shag Rocks shelves, but the density of juvenile fish was considerably greater on the Shag Rocks shelf, where 84% of the juveniles were caught (Table 1; Figs. 2, 3). Toothfish comprised 2.8–9.1% (by weight) of the fish catches at Shag Rocks and 0.7–6.7% of catches at South Georgia (Table 2). No Antarctic toothfish (Dissostichus mawsoni) were caught during the surveys.

Distribution (numbers caught) of juvenile Patagonian toothfish (Dissostichus eleginoides) (<750 mm TL) around South Georgia and Shag Rocks from surveys in 2003, 2004, 2005 and 2006

Length frequency distribution of Patagonian toothfish (Dissostichus eleginoides) from each survey showing the size of fish sampled (excludes trawls that targeted toothfish for tagging)
In each year the catch was dominated by a single cohort that was of size 200–250 mm in 2003 (putative 1+ year old), 300–360 mm in 2004, 380–460 mm in 2005 and 430–530 mm in 2006 (Fig. 3). A second, numerically smaller, cohort was seen in 2003 at around 400 mm TL, which was present in small numbers in 2005 and 2006. A third cohort was detected in 2006 at size 260–340 mm (putative 2+ years old) that was not seen the previous year.
Juvenile fish were generally confined to the continental shelf areas, with the largest catches taken at depths of less than 300 m and larger fish were more frequently caught at South Georgia (Fig. 3). For trawls that caught three or more Patagonian toothfish, mean size increased significantly with depth of capture (regression: F = 43.61; P < 0.001; Fig. 4), and although large fish are occasionally caught in shallow water, smaller fish are restricted to shallow depths.

Mean length of Patagonian toothfish (Dissostichus eleginoides) in relation to depth of capture from the four surveys. Error bars show standard deviations of length, only trawls with 3 or more fish included
Diet of juvenile toothfish
Stomach fullness was generally high, and less than 25% of stomachs were empty (23% in 2004, 19% in 2005; 24% in 2006). Average stomach fullness (ratio of stomach weight to body weight) was significantly higher in 2004 (2.52% BW) than 2005 (1.86%) and 2006 (2.12%) (ANOVA: F = 4.632; P < 0.01) (2003 excluded as full set of data not available). For stomachs containing food, contents weight averaged 2.78% of body weight (range 0.01–12.6%). There was no relationship between stomach fullness and time of day, but all trawls were conducted during daylight.
Stomach contents were examined from 795 toothfish that had full or partially full stomachs, of which 636 were from fish caught at Shag Rocks and 159 from South Georgia caught fish. The size distribution of sampled fish was approximately proportional to the size range caught, with the exception of the small sized fish (200–250 mm TL) caught in 2003, which were under-represented.
Juvenile toothfish (<750 mm) were predominantly ichthyophagous, with fish accounting for 95% of the diet by mass, 51% numbers, 88% frequency and 89% IRI (Table 3). The diet composition differed between South Georgia and Shag Rocks (Table 3; Fig. 5), with more crustaceans taken at South Georgia, but this may reflect the larger average size of fish caught off South Georgia compared to Shag Rocks (see above). At South Georgia more krill (52% by number) was taken, but when the diet is considered in terms of percent mass fish prey accounted for 89% of the diet at South Georgia, compared to 97% at Shag Rocks.

Diet of juvenile toothfish by size category from Shag Rocks (a) and South Georgia (b)
The fish component of the diet differed substantially between the two locations. At Shag Rocks the yellow-finned notothen (Patagonotothen guntheri) dominated the diet in each of the years (85% by mass; 95% IRI). Catches of both toothfish (Fig. 2) and P. guntheri (Fig. 6) were highest at the eastern end of the Shag Rocks shelf, although there was no significant correlation between catches of the two species at Shag Rocks. On the South Georgia shelf the diet was more diverse with the notothenids Lepidonotothen larseni (20% mass; 21% IRI) and Trematomus hansoni (23% mass; 3.9% IRI) the main fish prey species. Myctophid fish were also consumed, with Gymnoscopelus nicholsi and Protomyctophum bolini the most common species taken. There was a single incidence of southern blue whiting (Micromesistius australis), which is rarely found at South Georgia and a single incidence of an undescribed species of the Chiasmodontidae genus Pseudoscopelus (Marcelo Melo, personal communication; specimen lodged at the Natural History Museum, London: BMNH.2006.8.19.1).

Distribution of catches of the yellow-finned notothen (Patagonotothen guntheri) from surveys in 2003, 2004, 2005 and 2006
The main crustacean prey species were Antarctic krill (Euphausia superba), the mysids (Antarctomysis ohlini and A. maxima) and the pelagic amphipod (Themisto gaudichaudii), with the decapod, Notocrangon antarcticus, and isopods of the genus Natatolana (Family Cirolanidae) occasionally taken. Cephalopods were rare in the diet, with the octopus Adelieledone polymorpha and the squid Psychroteuthis glacialis the only species identified.
Ontogenetic changes in diet
The diet of toothfish changed with size at both Shag Rocks and South Georgia (Fig. 5: note sample sizes were smaller for larger fish and at South Georgia). At Shag Rocks the diet of fish size <500 mm TL was dominated by the notothenioid, Patagonotothen guntheri, which reflects the association with the Shag Rocks shelf. In fish greater than 400 mm TL there was an increase in krill and myctophids. At South Georgia, smaller fish consumed krill and other crustaceans, with larger fish taking notothenioid fish.
Fish prey size increased significantly with toothfish size (Regression: F = 83.571; P < 0.001; n = 832; Fig. 7) but, more clearly, the range of prey sizes taken increased with fish size. The main prey species, Patagonotothen guntheri, forms a cluster at sizes 70–200 mm TL, consumed by toothfish of 200–600 mm TL, but the relationship between predator size and prey size was still significant (Regression: F = 49.706; P < 0.001; n = 546). The larger prey items were other notothenioid fish, notably Trematomus hansoni and Champsocephalus gunnari. The myctophid prey, with the exception of a single large Gymnoscopelus bolini were of small size. The number of prey items consumed also increased with predator size.

Relationship between prey size and predator size in Patagonian toothfish (Dissostichus eleginoides)
The length of Patagonotothen guntheri consumed by toothfish was generally slightly smaller than that caught by the survey (not illustrated), the exception being in 2005, when the survey caught relatively smaller P. guntheri than in the previous seasons.
Discussion
Distribution
Whilst adult toothfish are distributed in deep-water all around South Georgia and Shag Rocks (Agnew et al. 1999; Agnew 2004), the data from this study suggest that the recruitment of juvenile toothfish occurs predominantly at Shag Rocks, with small numbers of juvenile fish caught on the South Georgia shelf, most notably at the SW edge. This is consistent with the data of Garcia de la Rosa et al. (1997), who only found small toothfish at Shag Rocks. Barrera-Oro et al. (2005) did catch some small fish on the South Georgia shelf, but the majority of fish <600 mm TL were caught on the Shag Rocks shelf. The association between toothfish recruitment and Shag Rocks is not clear, but may be related to temperature and oceanography. Water temperatures at Shag Rocks are slightly warmer than on the South Georgia shelf (M.A. Collins, unpublished data) and, as the Patagonian toothfish is at the southern edge of its range, and, unlike D. mawsoni, does not possess anti-freeze glycoproteins (Gon and Heemstra 1990), temperature may be a limiting factor.
Although two other cohorts were detected, this study essentially monitored a dominant cohort through four consecutive seasons. The cohort, first detected at size 200–250 mm TL, where probably 1+ years old in 2003 and would therefore be 4+ in 2006 (M. Belchier, unpublished data). The dominance of a single cohort through four years of sampling suggests strong interannual variability in recruitment. Toothfish are thought to spawn in winter in deep-water around South Georgia and Shag Rocks (Agnew et al. 1999). The eggs and larvae are both pelagic, with developmental stages thought to last 3 and 6 months respectively (Evseenko et al. 1995), and successful recruitment will be dependent on transport in near surface currents. The oceanography of the Scotia Sea is highly complex (Fig. 1) and subject to inter-annual variability, which may be the main factor driving recruitment variability. A detailed analysis of variability in growth and recruitment in Patagonian toothfish from 14 surveys from 1987 to 2006 is in progress (Belchier and Collins, in preparation).
In common with other species that scavenge as adults (see Collins et al. 2005), the Patagonian toothfish has a bigger-deeper trend (Arkhipkin et al. 2003; Laptikhovsky et al. 2006), and although this study was largely focused in shallow areas it showed a distinct pattern, which is supported by evidence from the fishery (Agnew et al. 1999). Larger fish do occasionally occur in shallow water and fish over a metre in length have been caught in trammel nets in <200 m depth in Cumberland Bay (personal observation).
Diet
Although this study only gives a summer snapshot of toothfish diet, it is clear that pre-recruit toothfish in the South Georgia/Shag Rocks area are essentially piscivorous, which is largely consistent with previous studies in the area (Zhivov and Krivoruchko 1990; Barrera-Oro et al. 2005) and dietary studies of similar sized toothfish in other parts of the range (Duhamel 1981; Garcia de la Rosa et al. 1997; Goldsworthy et al. 2002; Arkhipkin et al. 2003).
The difference in the specific composition of the toothfish diet between Shag Rocks and South Georgia is largely a consequence of distinct differences in the ichthyofauna between the two areas. The Shag Rocks and South Georgia shelves are separated by a deep (∼1,500 m) channel, and many of the notothenids and channichthyids that are common on the South Georgia shelf are rare or absent at Shag Rocks (see Table 2). At Shag Rocks the demersal fauna is less diverse and dominated by Lepidonotothen squamifrons, mackerel icefish and Patagonotothen guntheri, with the latter being absent from South Georgia (Gon and Heemstra 1990). Lepidonotothen squamifrons are, in some years, abundant at Shag Rocks and were the main fish prey identified by Barrera-Oro et al. (2005), however they are usually large fish and maybe too large for a juvenile toothfish to consume. Hence the most abundant fish species of suitable size is usually P. guntheri and the dominance of this species in toothfish diet may simply be a consequence of the distribution of the two species, with conditions that favour toothfish recruitment also favouring P. guntheri. Both species are abundant at the eastern end of the Shag Rocks shelf, which may be more productive than other parts of the Shag Rocks shelf. Interestingly, on the Falkland shelf the congeneric Patagonotothen ramsayi is the main prey of pre-recruit toothfish of sizes <600 mm TL (Arkhipkin et al. 2003).
The specific composition of the toothfish diet in this study differs from that identified from sampling in March–April 1996 by Barrera-Oro et al. (2005) and from 1985 to 1986 by Zhivov and Krivoruchko (1990). The Barrera-Oro et al. (2005) study found Lepidonotothen kempi (=L. squamifrons) to be the main toothfish prey at South Georgia and Shag Rocks and neither study found Patagonotothen guntheri to be so dominant [10% occurrence in Zhivov and Krivoruchko (1990) and not recorded by Barrera-Oro et al. (2005)]. Differences between the studies may reflect seasonal or inter-annual variability in prey availability, but in both of the other studies a large part of the fish diet was unidentified (49% unidentified in Barrera-Oro et al. 2005).
Patagonotothen guntheri, the dominant prey species at Shag Rocks, is a semi-pelagic notothenid that is one of the most abundant species caught during trawl surveys at Shag Rocks (Table 2), but is at the southern end of its range (Gon and Heemstra 1990). It is a pelagic feeder, consuming large copepods (e.g. Rhincalanus gigas), pelagic amphipods (e.g. Themisto gaudichaudii) and krill (M.A. Collins, unpublished data). On the South Georgia shelf the most important prey species were Lepidonotothen larseni and Trematomus hansoni, which also feed on macro-zooplankton. The mackerel icefish (Champsocephalus gunnari), which is abundant at both South Georgia and Shag Rocks (Table 2) and is commercially fished, was only occasionally found in toothfish stomachs in this study.
Even within the limited size range studied here, a change in diet with size of toothfish was detected. The shift from Patagonotothen guntheri to other notothenioids is associated with dispersion of fish from Shag Rocks to the South Georgia shelf. There is also an increase in krill consumption and, to a lesser extent, myctophid fish (particularly at Shag Rocks) with increased size. The principal myctophid species taken was Gymnoscopelus nicholsi, which is a relatively large species (upto 180 mm TL) abundant on the slope (300–1,000 m) around South Georgia and Shag Rocks, and although it is a pelagic species it is frequently caught in bottom trawls. This dietary change is associated with the ontogenetic migration into deeper water, where the available prey will differ, for instance Patagonotothen guntheri are rarely caught in depths greater than 300 m.
The migration to deep water is probably accompanied by changes in foraging behaviour as well as diet with large adults scavenging in addition to taking live prey (Garcia de la Rosa et al. 1997; Pilling et al. 2001; Arkhipkin et al. 2003), making them susceptible to baited long-lines and attracted to baited cameras (Collins et al. 1999, 2006; Yau et al. 2002). Arkhipkin et al. (2003) suggested that larger toothfish generally take less active prey than small toothfish, although adult fish are capable of bursts of high speed swimming (Yau et al. 2002), and consequently the diet of the larger toothfish is considerably different from that of the juveniles. At South Georgia the diet of adult fish appears more diverse than juveniles, with more cephalopods (Onychoteuthidae, Gonatidae, Chiroteuthidae and octopods) and crustaceans (krill, the decapod Nauticaris sp. and isopods) taken (Garcia de la Rosa et al. 1997; Pilling et al. 2001). The main fish families consumed were Myctophidae, Moridae and Zoarcidae, but in both studies over half the fish were not identified.
Although the size of prey increased with size of toothfish, it is clear that it is the range of prey size that increases, with the larger toothfish taking small prey as well as large prey items. The size of P. guntheri taken by toothfish was generally slightly smaller than that taken by the trawl survey and this is likely to be a consequence of both selectivity of the trawl and selectivity by the toothfish.
The diet of juvenile toothfish comprised a mixture of both pelagic and demersal species, and it is not known how much time toothfish spend foraging above the seafloor. For instance the main prey species P. guntheri feeds pelagically, but is also caught in bottom trawls. Time spent foraging above the seafloor will clearly impact their catchability in a bottom trawl, but potentially make toothfish more susceptible to diving predators (see below).
Given the importance of P. guntheri in the diet of toothfish recruits and the co-occurrence of the two species, it is likely that any exploitation of P. guntheri would impact on toothfish populations. Fishing for P. guntheri at Shag Rocks is likely to have a by-catch of small toothfish, and the removal of a large biomass of P. guntheri would reduce the available food for toothfish. Whilst there is currently no fishery for P. guntheri it has been estimated that around 170,000 tonnes were fished from the Shag Rocks area between 1969 and 1990 (Anon 1990a, b; Kock 1992). Currently a relatively small pelagic trawl fishery, targeting mackerel icefish (Champsocephalus gunnari), operates on the South Georgia and Shag Rocks shelves, which does have the potential to catch juvenile toothfish.
The role of toothfish in the South Georgia marine ecosystem
In order to have an ecologically sustainable fishery it is important to have knowledge of the diet of a target species and of potential predators that may be competing with the fishery. Whilst knowledge of the diet of toothfish is now substantial, little is known about the predators of toothfish. In shallow water, potential predators of juveniles include king and gentoo penguins, fur and elephant seals, but with increased size and habitat depth the range of potential predators is likely to decline (Table 4). From studies undertaken at on South Georgia, toothfish are rarely taken in the diets of fur seals or penguins, and only are occasionally taken by these species elsewhere (see Table 4). Toothfish have been reported in the diet of Weddell seals, of which there is a small population at South Georgia, and these are a potential predator. Albatross and white-chinned petrels are known to take toothfish, but these are, almost certainly, fish that escape from hooks or discards from fishing vessels. In deeper water, the only likely predators are elephant seals, sperm whales and large squid such as the Antarctic giant squid, Mesonychoteuthis hamiltoni. Both sperm and killer whales are known to take toothfish from longlines during hauling (Ashford et al. 1996; Kock et al. 2006; Purves et al. 2004), but toothfish habitat is beyond the normal diving capabilities of killer whales. Little is know about the ecology of Mesonychoteuthis hamiltoni, but these large squid are probably capable of catching and consuming large toothfish, and are occasionally caught on long-line hooks at South Georgia (M.A. Collins, unpublished data). Although cannibalism was not recorded in this study it has been recorded in larger toothfish (Arkhipkin et al. 2003) and may occur between cohorts. Overall the evidence from predators indicates low levels of predation, which is a consequence of the depth distribution and size of the toothfish.
References
Abe T, Iwami T (1989) Notes on fishes from the stomachs of whales taken in the Antarctic II. On Dissostichus and Ceratius, with an appendix (Japanese names of important Antarctic fishes). Proc NIPR Symp Pol Biol 2:78–82
Adams NJ, Klages NT (1989) Temporal variation in the diet of the gentoo penguin Pygoscelis papua at Sub-Antarctic Marion Island. Colon Waterbirds 12:30–36
Agnew D, Heaps L, Jones C, Watson A, Berkieta K, Pearce J (1999) Depth distribution and spawning pattern of Dissostichus eleginoides at South Georgia. CCAMLR Sci 6:19–36
Agnew DJ (2004) Fishing South: History and Management of South Georgia Fisheries. Government of South Georgia and the South Sandwich Islands
Anon (1990a) Statistical bulletin, vol 2 (1980–1989). CCAMLR, Hobart
Anon (1990b) Statistical bulletin, vol I (1970–1979). CCAMLR, Hobart
Arkhipkin A, Brickle P, Laptikhovsky V (2003) Variation in the diet of the Patagonian toothfish with size, depth and season around the Falkland Islands. J Fish Biol 63:428–441
Ashford JR, Rubilar PS, Martin AR (1996) Interactions between cetaceans and longline fishery operations around South Georgia. Mar Mam Sci 12:452–457
Barrera-Oro ER, Casaux RJ, Marschoff ER (2005) Dietary composition of juvenile Dissostichus eleginoides (Pisces, Nototheniidae) around Shag Rocks and South Georgia, Antarctica. Polar Biol 28:637–641
Boyd IL, Walker TR, Poncet J (1996) Status of southern elephant seals at South Georgia. Antarct Sci 8:237–244
Brown CR, Klages NT (1987) Seasonal and annual variation in diets of macaroni (Eudyptes chrysolophus chrysolophus) and southern rockhopper (E. chrysocome chrysocome) penguins at sub-Antarctic Marion Island. J Zool Lond 212:7–28
Calhaem I, Christoffel DA (1969) Some observations of the feeding habits of a Weddell seal, and measurements of its prey, Dissostichus mawsoni, at McMurdo Sound, Antarctica. NZ J Mar Fresh Res 3:181–190
Catard A, Weimerskirch H, Cherel Y (2000) Exploitation of distant Antarctic waters and close shelf-break waters by white-chinned petrels rearing chicks. Mar Ecol-Prog Ser 194:249–261
Cherel Y, Duhamel G (2004) Antarctic jaws: cephalopod prey of sharks in Kerguelen waters. Deep-Sea Res I 51:17–31
Cherel Y, Ridoux V, Rodhouse PG (1996) Fish and squid in the diet of king penguin chicks, Aptenodytes patagonicus, during winter at sub-antarctic Crozet Islands. Mar Biol 126:559–570
Cherel Y, Weimerskirch H, Trouve C (2000) Food and feeding ecology of the neritic-slope forager black- browed albatross and its relationships with commercial fisheries in Kerguelen waters. Mar Ecol-Prog Ser 207:183–199
Cherel Y, Weimerskirch H, Trouve C (2002) Dietary evidence for spatial foraging segregation in sympatric albatrosses (Diomedea spp.) rearing chicks at Iles Nuageuses, Kerguelen. Mar Biol 141:1117–1129
Clarke MR (1980) Cephalopods in the diet of sperm whales of the southern hemisphere and their bearing on sperm whale biology. Discovery Reports XXXV11. Institute of Oceanographic Sciences
Collins MA, Yau C, Nolan CP, Bagley PM, Priede IG (1999) Behavioural observations on the scavenging fauna of the Patagonian slope. J Mar Biol Assoc UK 79:963–970
Collins MA, Bailey DM, Ruxton G, Priede IG (2005) Trends in body size across an environmental gradient: a differential response in scavenging and non-scavenging demersal deep-sea fish. Proc R Soc Lond B 272:2051–2057
Collins MA, Everson I, Paterson R, Bagley PM, Yau C, Belchier M, Hawkins S (2006) In situ observations of the scavenging fauna of the South Georgia slope. In: Shotton R (ed) Deep Sea 2003: conference on the governance and management of deep-sea fisheries. Part 2: conference poster papers and workshop papers. FAO, Rome, pp 487
Constable AJ, de la Mare WK, Agnew DJ, Everson I, Miller D (2000) Managing fisheries to conserve the Antarctic marine ecosystem: practical implementation of the Convention on the Conservation of Antarctic Marine Living Resources (CCAMLR). ICES J Mar Sci 57:778–791
Cortes E (1996) A critical review of methods of studying fish feeding based on analysis of stomach contents: application to elasmobranch fishes. Can J Fish Aquat Sci 54:726–738
Duhamel G (1981) Characteristiques biologiques des principals especes de poissons du plateau continental des Iles Kerguelen. Cybium 5:19–32
Evseenko SA, Kock KH, Nevinsky MM (1995) Early life history of the Patagonian toothfish, Dissostichus eleginoides Smitt, 1898 in the Atlantic Sector of the Southern Ocean. Antarct Sci 7:221–226
Garcia de la Rosa SB, Sanchez F, Figueroa D (1997) Comparative feeding ecology of Patagonian toothfish (Dissostichus eleginoides) in the South-western Atlantic. CCAMLR Sci 4:105–124
Goldsworthy SD, He X, Tuck GN, Lewis M, Williams R (2001) Trophic interactions between the Patagonian toothfish, its fishery, and seals and seabirds around Macquarie Island. Mar Ecol-Prog Ser 218:283–302
Goldsworthy SD, Lewis M, Williams R, He X, Young JW, van den Hoff J (2002) Diet of Patagonian toothfish (Dissostichus eleginoides) around Macquarie Island, South Pacific Ocean. Mar Freshw Res 53:49–57
Gon O, Heemstra PC (1990) Fishes of the Southern Ocean. J.L.B. Smith Institute of Ichthyology, Grahamstown
Green K, Burton HR, Williams R (1989) The diet of Antarctic fur seals Arctocephalus gazella (Peters) during the breeding season at Heard Island. Antarct Sci 1:317–324
Hecht T (1987) A guide to otoliths of Southern Ocean fishes. S Afr J Antarct Res 17:1–87
Horn PL (2002) Age and growth of Patagonian toothfish (Dissostichus eleginoides) and Antarctic toothfish (D. mawsoni) in waters from the New Zealand subantarctic to the Ross Sea, Antarctica. Fish Res 56:275–287
Hulley PA (1981) Results of the research cruises of FRV “Walther Herwig” to South America. 58. Family Myctophidae (Osteichthyes, Myctophiformes). Archiv für Fischereiwissenschaft 31 (Supplement 1):300 pp + 3 pp of index
Kock KH (1992) Antarctic fish and fisheries. Cambridge University Press, Cambridge
Kock KH, Purves MG, Duhamel G (2006) Interactions between cetaceans and fisheries in the Southern Ocean. Polar Biol 29:379–388
Kooyman GL, Cherel Y, Le Maho Y, Croxall JP, Thorson PH, Ridoux V, Kooyman CA (1992) Diving behaviour and energetics during foraging cycles in king penguins. Ecol Monogr 62:143–163
Korabelnikov LV (1959) The diet of sperm whales in the Antarctic seas. Priroda 3:103–104
Lake S, Burton H, van den Hoff J (2003) Regional, temporal and fine-scale spatial variation in Weddell seal diet at four coastal locations in east Antarctica. Mar Ecol-Prog Ser 254:293–305
Laptikhovsky V, Arkhipkin A, Brickle P (2006) Distribution and reproduction of the Patagonian toothfish Dissostichus eleginoides Smitt around the Falkland Islands. J Fish Biol 68:849–861
Lescroel A, Ridoux V, Bost CA (2004) Spatial and temporal variation in the diet of gentoo penguin (Pygoscelis papua) at Kerguelen Islands. Polar Biol 27:206–216
McKenna JE (1991) Trophic relationships within the Antarctic demersal fish community of South Georgia Island. Fish Bull 89:643–654
Nesis KN (1987) Cephalopods of the world. T.N.P publications, New York
Olsson O, North AW (1997) Diet of the king penguin Aptenodytes patagonicus during three summers at South Georgia. Ibis 139:504–512
Pilling GM, Purves MG, Daw TM, Agnew DA, Xavier JC (2001) The stomach contents of Patagonian toothfish around South Georgia (South Atlantic). J Fish Biol 59:1370–1384
Plotz J (1986) Summer diet of Weddell seals (Leptonychotes weddelli) in the eastern and southern Weddell Sea, Antarctica. Polar Biol 6:97–102
Purves MG, Agnew DJ, Balguerias E, Moreno CA, Watkins B (2004) Killer whale (Orcinus orca) and sperm whale (Physeter macrocephalus) interactions with longline vessels in the Patagonian toothfish fishery at South Georgia, South Atlantic. CCAMLR Sci 11:111–126
Reid K (1995) The diet of Antarctic fur seals (Arctocephalus gazella Peters 1875) during winter at South Georgia. Antarct Sci 7:241–249
Reid K (1996) A guide to the use of otoliths in the study of predators at South Georgia. British Antarctic Survey
Reid K, Arnould JPY (1996) The diet of Antarctic fur seals Arctocephalus gazella during the breeding season at South Georgia. Polar Biol 16:105–114
Reid K, Nevitt GA (1998) Observation of southern elephant seal, Mirounga leonina, feeding at sea near South Georgia. Mar Mam Sci 14:637–640
Robinson SA, Hindell MA (1996) Foraging ecology of gentoo penguins Pygoscelis papua at Macquarie Island during the period of chick care. Ibis 138:722–731
Slip DJ, Hindell MA, Burton HR (1994) Diving behaviour of southern elephant seals from Macquarie Island: an overview. In: Le Boeuf BG, Laws RM (eds) Elephant seals: population ecology, behaviour and physiology. University of California Press, Berkeley, pp 253–270
Testa JW, Siniff DB, Ross MJ, Winter JD (1985) Weddell seal—Antarctic cod interactions in McMurdo Sound, Antarctica. In: Siegfried WR, Condy PR, Laws RM (eds) Antarctic nutrient cycles and food webs. Springer, Berlin, pp 561–565
Watkins WA, Daher MA, Fristrup KM, Howald TJ, Disciara GN (1993) Sperm whales tagged with transponders and tracked underwater by sonar. Mar Mam Sci 9:55–67
Xavier JC, Croxall JP, Trathan PN, Wood AG (2003) Feeding strategies and diets of breeding grey-headed and wandering albatrosses at South Georgia. Mar Biol 143:221–232
Yau C, Collins MA, Bagley PM, Everson I, Priede IG (2002) Scavenging by megabenthos and demersal fish on the South Georgia slope. Antarct Sci 14:16–24
Young E (2003) Monsters of the deep. New Sci 2 Aug 2003:24–30
Zhivov VV, Krivoruchko VM (1990) On the biology of the Patagonian toothfish, Dissostichus eleginoides, of the Antarctic part of the Atlantic. J Ichthyol 30:142–146
Acknowledgments
Thanks to Len Featherstone (Master) and the crew of the FPRV Dorada for their efforts during the surveys in 2003 and 2004. Sarah Clark, Therese Cope, Mike Endicott, Inigo Everson, Suzi Hawkins, Tom Marlow, Richard Mitchell, Tony North, Martin Purves, Jacek Szlakowski, Jamie Watts and Will Reid assisted with sorting trawl catches. Thanks to Geraint Tarling for assistance in identification of crustacean prey, Peter Rothery for statistics advice and to Claire Waluda and Sally Thorpe for help with the figures. South Georgia Groundfish Surveys were funded by the Government of South Georgia and the South Sandwich Islands. This is a contribution to the BAS Discovery 2010 Programme.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by O. Kinne.
Rights and permissions
About this article
Cite this article
Collins, M.A., Ross, K.A., Belchier, M. et al. Distribution and diet of juvenile Patagonian toothfish on the South Georgia and Shag Rocks shelves (Southern Ocean). Mar Biol 152, 135–147 (2007). https://doi.org/10.1007/s00227-007-0667-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-007-0667-3