
Abstract
Phenotypic plasticity, a widespread phenomenon in boreal freshwater fishes, is less apparent in the marine realm and the organism–environment interactions producing this variation are undetermined. A sample of 40 specimens of Trematomus newnesi, an inshore Antarctic fish from King George/25 de Mayo Island in the South Shetlands, was composed of 52.5% typical morphs, 27.5% large-mouth morphs and 20% intermediate morphs. Measurements of percentage buoyancy on the morphs of this sample were 3.73, 3.84 and 3.83%, respectively with no significant differences among means. Both mean dry skeletal weight as a percentage of body weight and mean oral jaw weight as a percentage of dry skeletal weight were significantly greater in large-mouth morphs compared to typical morphs. Diversification in head and jaw morphology is not accompanied by diversification in ecology as represented by buoyancy and, in spite of external appearances, measurements of buoyancy offer no support for the hypothesis that the large-mouth morph is more benthic than the typical semipelagic morph. Although a trophic basis for this polymorphism is possible, it has not yet been documented. Our discussion centers on the status of key open questions regarding morphism and highlights areas requiring more research.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Differential use of benthic and water column habitats is a major axis of diversification in the adaptive radiation of Antarctic notothenioid fishes, and the divergent buoyancies of various species are reflected in occupation of these habitats (Eastman and De Vries 1982; Eastman 1993; Eastman and Sidell 2002). The common inshore nototheniid Trematomus newnesi is semipelagic and exemplifies this phyletic trend away from the ancestral benthic habitat toward habitation of the water column. This species is found in the permanent and seasonal pack ice zones around Antarctica and the adjacent islands mainly in inshore waters from 20–45 m depths on rocky bottoms with macroalgae beds (De Witt et al. 1990; Barrera-Oro 2002). It also may be found as deep as 400 m (Tiedtke and Kock 1989). T. newnesi feeds opportunistically on zooplanktonic prey in the water column or at the undersurface of the pack ice, with the euphausiids Euphausia superba and E. crystallorophias, amphipods, and larval fish especially important in the diet (Casaux et al. 1990, 2003; De Witt et al. 1990; Vacchi and La Mesa 1995; La Mesa et al. 2000; Barrera-Oro 2003; Barrera-Oro and Piacentino 2007). T. newnesi is also known to forage on the substrate (De Witt et al. 1990) and to live and feed in macroalgae beds as a subadult (Barrera-Oro and Piacentino 2007).
While many aspects of its biology are well known, our understanding of the ecology of T. newnesi is confounded by its existence as populations of phenotypically plastic morphs, a feature that distinguishes it from all other notothenioids. Also unusual is the fact that this phenotypic plasticity has not yet been documented as an example of trophic polymorphism. We employ West-Eberhard’s (2003), p. 34 definition of phenotypic plasticity as “the ability of an organism to react to an environmental input with a change in form, state, movement, or rate of activity”. This is a growing and vigorous area of research in evolutionary biology with recent syntheses on the topic appearing as books (West-Eberhard 2003; De Witt and Scheiner 2004). Phenotypic plasticity likely plays a role in the diversification of taxa, especially in changing or heterogeneous environments (Schlichting 2004). It promotes the occupation of new niches by providing phenotypic alternatives (Skúlason et al. 1999; Schlichting 2004) as well as by contributing to the structuring of food webs, genetic differentiation and speciation (Agrawal 2001).
The population of T. newnesi in McMurdo Sound consists of two morphs: the typical morph and a large-mouth morph with wider and blunter head, longer upper jaw, wider gape and dark uniform coloration (Eastman and De Vries 1997). Recently these two morphs as well as an intermediate morph have been found at Potter Cove, King George/25 de Mayo Island in the South Shetland Islands and at Petermann Island, western Antarctic Peninsula (Piacentino and Barrera-Oro 2009). Piacentino and Barrera-Oro also discovered that eye diameter relative to head length, smaller in the large-mouth morph, also separates the morphs. They were able to identify the morphs in specimens as small as 60 mm TL and as large as 241 mm TL. Intermediate morphs are now also known from McMurdo Sound (Eastman and De Vries, unpublished data). The most perplexing aspect of morphism in T. newnesi is the apparent lack of association with divergence in habitat or diet. Based on inferences from morphology and colour, Eastman and De Vries (1997) speculated that the large-mouth morph was more benthic than the typical semipelagic morph, but they did not measure buoyancy and found no differences in diet. Barrera-Oro and Piacentino (2007) identified an ontogenetic change in diet with larger T. newnesi in Potter Cove tending to be more pelagic and planktivorous, but they did not perform a strict comparison of diets between morphs. In some aspects of their morphology, the T. newnesi morphs resemble the intraspecific benthic–pelagic morphs in salmonids especially Arctic charr Salvelinus alpinus (Skúlason et al. 1989, 1999; Jonsson and Jonsson 2001).
Notothenioids lack a swim bladder, hence interspecific modification of buoyancy is attributable to alterations in density as a result of reduction in skeletal ossification and/or deposition of lipids (Eastman 1993). Measurements of buoyancy are expressed as percentage buoyancy (%B), calculated as weight in water/weight in air × 100. Interspecific variation in this measurement among notothenioids encompasses a range from neutrally buoyant species with no weight in water (%B ≈ 0) to heavy benthic species (%B ≈ 6.0). Most notothenioids, however, are intermediate between these two extremes (Eastman 1993; Eastman and Sidell 2002). As measured by the standard deviation, intraspecific variation in %B is ±0.2–0.5%B units across widely separated populations of a species (Near et al. 2009). Only one buoyancy measurement is available for T. newnesi; a 188-SL specimen from McMurdo Sound had a %B of 2.62 (Eastman 1993).
The availability of the three morphs of T. newnesi in Potter Cove and the proximity of laboratory facilities at Jubany Station for accurate determination of buoyancy, allowed us to test the hypothesis of Eastman and De Vries that morphs may exhibit ecological diversification based on buoyancy. In this paper we present: (1) a comparison of measurements of buoyancy for three morphs of T. newnesi; (2) a comparison of skeletal weights for large-mouth and typical morphs and (3) a discussion of the status of key questions regarding the morphs highlighting areas requiring more research.
Materials and methods
We collected 50 specimens of T. newnesi at Potter Cove, King George/25 de Mayo Island (Fig. 1a), close to the scientific station Jubany (62°14′S; 58°40′W) during the summer period 30 December 2008 to 3 March 2009. Casaux et al. (1990) provide detailed information on the biota and abiotic features of this site. We used two types of gear for sampling and deployed gear from rubber boats at different sites in the cove, primarily where the seabed was uniformly rocky and covered with red and brown macroalgae (Fig. 1b). A bottom trawl (mouth 1 m2, length 2 m and mesh 4 mm) was towed during daylight for 15–30 min at depths of 6–30 m (average, 12 m). Trammel nets (length 15 m, width 1.5 m, inner mesh 2.5 cm, outer mesh 12 cm) were fixed to rocks, cast in various directions and set on the bottom at depths of 10–40 m (average, 14 m) for 16–24 h. After capture the fish were maintained alive in the aquarium at the station until processed and measured for buoyancy. Thirty-five of the specimens were held for 1–3 h, four for 48 h and one for 23 days. Water temperature in the tanks was 1–1.7°C. Six specimens died during captivity and four had weights that were erroneous (i.e. we misread the balance display or did not properly tare the balance in three cases, and the %B was >3SD below the mean in one case, also likely due to a balance error). Thus only 40 of the 50 fish caught were used for determination of buoyancy.

Map of King George/25 de Mayo Island in the South Shetland Islands (a), showing the sampling sites for Trematomus newnesi in Potter Cove (b). The shaded area and zone 1 correspond to areas of bottom trawling and trammel net deployment, respectively
In the laboratory at Jubany we obtained total (TL) and standard (SL) lengths of the fish and determined their buoyancy. This involves weighing the fish in water of a temperature similar to its habitat and then in air, with the relative proportion of these weights expressed as the percentage buoyancy. Near et al. (2009) recently summarized the procedure and apparatus used in making these measurements. We followed this protocol with the exception that our specimens were not anesthetized. We used electronic balances with accuracies of 0.01 g and, in the case of specimens <70 mm SL, 0.001 g. Water temperature in the container used for measurements was 1–2°C. For some of our analyses we grouped the fish into small (62–69 mm SL), medium (119–149 mm SL) and large (150–187 mm SL) size ranges. We determined sex and maturity stages on the basis of a five-point scale used for notothenioids (Kock and Kellermann 1991). Following a method of counting annuli similar to that described by Barrera-Oro and Casaux (1996), we estimated ages by reading scales from a limited number of the smaller (≤69 mm SL) and larger (≥150 mm SL) specimens.
For identification of the T. newnesi morphs, we employed the descriptions, drawings and photographs in Eastman and De Vries (1997) and Piacentino and Barrera-Oro (2009). We used variation in the following characters, most are obvious to the human eye, and in proportional measurements to separate the morphs (condition in the typical morph given first): (1) position of the posterior end of the upper jaw (maxilla) relative to the middle of the eye → anterior or posterior; (2) head width → narrower or wider; (3) head shape in dorsal view → V-shaped or U-shaped; (4) gape width (maximum jaw width/head length) → smaller or larger; (5) eye (orbit) length/head length → larger or smaller; (6) colour → mottled with yellow or orange-brown coloration especially on pectorals and caudal or more homogeneous dark grey. We designated as intermediate morphs those fish that showed intermediate states for some of these characters. Characters such as length of the upper jaw relative to the eye and head shape do not necessarily co-occur in intermediate morphs and thus they may not exhibit intermediate states for both characters. The morphometric character upper jaw length/head length can be use in conjunction with or in place of the qualitative character (1) above relating position of the maxilla to the middle of the eye. Relative eye size is not distinguishable without measurement.
We prepared dry skeletons from five frozen specimens each of the large-mouth (SL = 174–193 mm) and typical (SL = 162–179) morphs. This involved macerating the fish in soapy water for 3–7 days at room temperature (23°C), cleaning the bones with running water and compressed air, and drying at room temperature to a constant weight (6 weeks). Weights for oral jaws include left and right premaxillae, maxillae and dentaries and these weights were obtained on a balance with an accuracy of 0.0001 g. We also used needle-point calipers to measure length of the ascending process of the premaxilla and total length of the premaxilla, maxilla, dentary and pharyngeal jaws (fifth ceratobranchials) in the dried bones of this sample. Four of these specimens were from the 2009 sample described above, while the other six were from a previous (2008) sample for which measurements of buoyancy were not made.
We used the programs Excel and SPSS 16.0 for statistical analysis. To normalize distributions, we transformed data to natural logarithms (ln) or performed an arcsine transformation on percentage measurements. We employed Shapiro–Wilks tests to determine if data were normally distributed. Data values in the text are untransformed but reported levels of significance are for ln- or arcsine-transformed data. Our null hypotheses were that there were no differences among morphs in means for percentage buoyancy, standard length, body weight, condition factor, percentage skeletal weight and percentage oral jaw weight. We used one-way ANOVAs to test the equality of means among the three morphs and independent t tests to compare data between large-mouth and typical morphs.
Results
Morph frequency, size, age and sex ratio
Of the 40 fish analysed for buoyancy, 52.5% were the typical morph, 27.5% the large mouth and the remaining 20% were intermediate forms. Figure 2 provides the length–frequency distribution of the morphs. The sample included a wide range of sizes from small juveniles (71 mm TL, 62 mm SL) to large adults (216 mm TL, 186 mm SL). The length intervals between 70 and 99 mm SL were not represented, possibly because of net selectivity and/or escape of fish. There was a predominance of large-mouth morphs among large fish ≥200 mm TL (171 mm SL). Likewise, the smaller specimens (SL <110 mm), and especially those <70 mm SL, were exclusively typical, while the medium-size fish (SL = 120–169 mm) were typical, intermediate, or in smaller proportions, large-mouth morphs. The youngest and oldest fish belonged to age groups 1 and 7 years, respectively. Stages of gonadal development were 1 (immature) to 3 (developing). In 27% of the specimens sex could not be determined macroscopically; in the remaining specimens, the ratio between males and females was 1.6 to 1.

Length–frequency distribution of the morphs of 50 specimens of Trematomus newnesi from Potter Cove. Buoyancy was determined in 40 of these specimens
Buoyancy
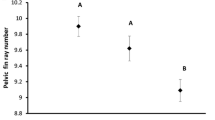
Figure 3 displays the results of our measurements of buoyancy and descriptive statistics for the three morphs. The values for the entire sample encompassed a range of 1.2%B units which is at the low end of the range of intraspecific variation in this measurement for the four other notothenioid species studied to date (Near et al. 2009). At 0.2–0.3%B units, dispersion around the mean (SD) was less than usual. A one-way ANOVA indicated no significant differences among mean %B of the three morphs (F 2,39 = 0.548; P = 0.583; Tukey HSD P = 0.707); this is also indicated by the overlap of the 95% confidence levels in Fig. 3. A one-way ANOVA also indicated no significant differences in %B among standard length-based groups of small, medium and large sized-specimens (F 2,39 = 0.938; P = 0.400; Tukey HSD P = 0.467). One-way ANOVA assuming unequal variances indicated that the large-mouth morphs were significantly heavier (F 2,39 = 12.604; P < 0.0001; Tamhane P < 0.0001) with a significantly greater standard length (F 2,39 = 11.737; P < 0.0001; Tamhane P < 0.0001) than the other two morphs. Trend lines in scatter plots (not shown) indicated that %B increased slightly during ontogeny but linear regression of %B against both standard length and weight revealed that these correlations were not significant (Pearson = 0.211; P = 0.096 and Pearson = 0.202; P = 0.106, respectively).

Measurements of buoyancy and descriptive statistics for 40 specimens of three morphs of Trematomus newnesi from Potter Cove. Circles are individual measurements, vertical lines are means and shaded rectangles are 95% confidence intervals for the mean
As seen in Fig. 3, the scatter of %B measurements for the typical morph accounted for most (1.1%B units) of the range of 1.2%B units in our measurements. All ten specimens in the small-size (≤69 mm SL) group were typical morphs (21 total) and their weighs in air were 2.5–3.9 g. The error in weighing these small fish may be considerably greater than for larger specimens because the procedure for weighing in air involves blotting fish dry to remove excess water, and the degree to which this is done can introduce variability in the weights in air. These small specimens weighed fivefold less than the next smallest specimens (20 g) and are the smallest notothenioids that have been subject to determination of buoyancy. Variability may be an inherent characteristic of determinations of buoyancy of these small specimens.
We also note that one of our specimens of the large-mouth morph, as part of a physiology experiment, was held unfed in an aquarium at Jubany Station for 23 days before its buoyancy was measured. Nevertheless its %B (3.77) and condition factor (1.101) are near the mean values for each parameter. This suggests, as previously indicated (Near et al. 2009), that factors such as gut contents, reproductive status and minor fluctuations in body condition have relatively little influence on measurements of buoyancy in notothenioids.
Skeletal weights and jaw morphology
Both mean dry skeletal weight as a percentage of body weight (1.68 vs. 1.38%; t = 3.004; P = 0.02) and mean oral jaw weight as a percentage of dry skeletal weight (11.10 vs. 8.47%; t = 3.739; P = 0.01) were significantly greater in a sample of five large-mouth morphs compared to five typical morphs. Figure 4 shows the oral and lower pharyngeal (fifth ceratobranchials) jaws from large-mouth and typical morphs of nearly identical standard length. The oral jaws of the large-mouth morph weighed 62% more on a dry weight basis and 44% more as a percentage of skeletal weight than those of the typical morph. Furthermore, in this particular comparison the maxilla, premaxilla and dentary were 26, 33 and 26% longer, respectively in the large-mouth morph. The discrepancy between the length of the lower pharyngeal jaws was only 16%. The oral and pharyngeal jaws of the two morphs were of similar shape; the ventral process of the body of the dentary was notably more massive in the large-mouth morph. The teeth were also of similar morphology, conical with sharp tips.

The oral and lower pharyngeal jaws of (top) a large-mouth morph (SL = 178 mm; weight = 117.9 g) and (bottom) a typical morph (SL = 179 mm; weight = 112 g) of similarly sized specimens of Trematomus newnesi from Potter Cove. Shown are lateral views of the left maxillae, premaxillae and dentaries and anterior views of the lower pharyngeal bones. Teeth are missing because in most notothenioids they are not attached to the jaws by a bony ankylosis and are thus lost during preparation of the skeleton. Scale bar is 10 mm long
Although the mean length of the ascending process of the premaxilla relative to premaxillary length was longer in typical (28.31%) than in large-mouth morphs (25.13%), the difference was not significant (N = 5; t = 1.715; P = 0.14). This suggests that there is probably no difference in the degree of premaxillary protrusion between the two morphs. The angle between the ascending process and the anterior–posterior axis of the bone was slightly larger in the typical morph (Fig. 4).
Discussion
Buoyancy in trematomid species and T. newnesi morphs
Phenotypic diversity among phyletically related species of fishes typically reflects the diversity in their ecology. The extent of habitat divergence among trematomid species is sufficient to be manifest in values for percentage buoyancy. For example, the epibenthic Trematomus loennbergii (2.28%) and cryopelagic Pagothenia borchgrevinki (2.75%) have significantly lower values than the benthic T. bernacchii (3.37%) (Eastman 1993). The relatively high mean value (3.78%) for our sample of T. newnesi from Potter Cove, compared to a single previous measurement from McMurdo Sound at 2.62% (Eastman 1993), is not unusual and is actually near the low end of variation for the range of intraspecific measurements of buoyancy in notothenioids which span 1–2.5%B units (Near et al. 2009). T. newnesi has traditionally been considered semipelagic and another water column species, the streamlined epibenthic T. eulepidotus, also has a relatively high value for %B at 3.89% (Eastman and Sidell 2002). While values for %B in nototheniids are in general agreement with documented aspect of the life history of most species, there are exceptions as in the case of these two species of Trematomus, where appearance, including body shape, streamlining and lack of substrate contact adaptations, does not accurately predict buoyancy. Lepidonotothen larseni and Gobionotothen gibberifrons serve as another example. The former, with a laterally compressed streamlined body and a terminal mouth, has the appearance of a pelagic or semipelagic species but it has a significantly higher %B (5.62%) than G. gibberifrons (4.54%) which has a dorsoventrally flattened body typically associated with benthic browsing (Eastman and Sidell 2002). In some nototheniids this dissociation between percentage buoyancy and external morphology may reflect a broad feeding spectrum.
The relative skeletal weights for the T. newnesi morphs from Potter Cove (PC) may be compared with those of the same and other trematomids from McMurdo (McM) Sound (Eastman, unpublished data) when arranged in a heavier to lighter sequence: T. newnesi (large mouth, PC), 1.68%; T. bernacchii (McM), 1.59%; T. newnesi (typical, McM), 1.42%); T. newnesi (typical, PC), 1.38% and Pagothenia borchgrevinki (McM), 1.30%. These are all relatively light skeletons, as is characteristic of trematomids, but they do reflect a trend for the more pelagic species to have the lightest skeletons as exemplified by P. borchgrevinki. Although in T. newnesi from Potter Cove the skeleton of the large-mouth morph is significantly heavier than that of the typical morph, their percentage buoyancies are not significantly different. This is likely attributable to the higher mean body weights and condition factors in large-mouth morphs. While the differences in condition factors were not significant in the comparison between the five particular specimens of each morph used for the preparations of the skeletons, for the entire sample one-way ANOVAs assuming unequal variances indicated that the large-mouth morphs were significantly heavier (F 2,39 = 12.604; P < 0.0001; Tamhane P < 0.0001) with a significantly greater condition factor (F 2,39 = 12.745; P < 0.0001; Tamhane P < 0.0001) than the other two morphs. Compared to typical morphs of the same length, much of the additional weight in the large-mouth morphs is attributable to viscera, visceral lipid and muscle, and these lower-density tissues probably offset greater skeletal weight.
As for the values for percentage buoyancy among the intraspecific morphs of T. newnesi, Eastman and De Vries (1997) assumed the fusiform shape and colouring of the typical morph suggested that it was more pelagic than the large-mouth morph. However, our measurements of morphs from Potter Cove showed no significant differences in the buoyancy of the three morphs. The diversification in head and jaw morphology is not accompanied by diversification in ecology as represented by buoyancy. Thus, the morphs of T. newnesi are not polar marine equivalents of the numerous well-studied examples of benthic and pelagic morphs and species in a diverse array of lacustrine fish taxa where habitat and dietary differences have been documented (Skúlason et al. 1989; Robinson and Wilson 1994, 1996; Robinson and Schluter 2000).
Ecology of the T. newnesi morphs in Potter Cove
Based on analysis of specimens of T. newnesi collected in this and previous studies at Potter Cove, there is no evidence to support the premise that the typical and large-mouth morphs are segregated by habitat. The relevant observations are these. First, historically sampling has been carried out using demersal (trammel nets) and bottom (trawl net) gear and both morphs have been always caught together near the bottom. Second, because we only became aware of the existence of the morphs in Potter Cove in the last 2 years, two previous studies on the diet of this species were carried out on the entire sample of fish without a comparative analysis between morphs (Casaux et al. 1990; Barrera-Oro and Piacentino 2007). Furthermore, our most recent study, which dealt exclusively with early juvenile stages (4–154 mm TL), does not support the hypothesis of niche diversification because all fish had fed on a few benthic–demersal taxa associated with algal communities, with no differences in feeding habitats between fish sizes (Barrera-Oro and Piacentino 2007).
The neritic waters of Potter Cove (depth <120 m), where most notothenioids usually occur on rocky bottoms with macroalgae beds at <70 m, are probably not an appropriate site for testing a hypothesis postulating a difference in diet between the morphs. In Potter Cove the depth distribution of the main pelagic prey of large juveniles and adult T. newnesi, such as krill in summer and hyperiid amphipods (Casaux et al. 1990), may reach the substrate therefore being equally available to both the supposedly “more benthic” large-mouth as well as the “more semipelagic” typical morphs. A more suitable dietary comparison would involve sampling morphs collected in deeper offshore waters somewhere else within the range of T. newnesi.
Taking stock of the morphs of T. newnesi: still more unknowns than knowns
Because ecological (responses to environmental variation) and evolutionary (genetic change via natural selection) dynamics are closely intertwined, we have limited understanding of their relative contributions to phenotypic plasticity under natural conditions (Ozgul et al. 2009). While this is certainly true for T. newnesi, we will nevertheless consider various aspects of this topic with an eye toward updating the status of the morphs and guiding future research. First, does the morphism have a genetic basis? The methodology of modern population genetics involving DNA sequence variations, such as restriction fragment-length polymorphisms and single-nucleotide polymorphisms, has not yet been applied to this question. Six polymorphic microsatellites are also now available for T. newnesi (Van Houdt et al. 2006). In the absence of genetic data, it cannot be said whether the morphology seen in the large-mouth morph is environmentally induced (ecophenotypic), heritable or a combination of both. Research in this area is a high priority.
Which morph is derived (in the sense that it represents a departure from “normal” morphology)? Without genetic data and assuming there has been no phyletic diversification, we hypothesize that the large-mouth morphology is derived because it appears relatively late in ontogeny (see below) and lacks the shape and colouring of a typical larva (i.e. it is not derived by paedomorphy from the more common typical morph). All notothenioids have pelagic larval and/or juvenile stages that last from several months to over 1 year (Loeb et al. 1993) and, in the case of T. newnesi, those from the Antarctic Peninsula region have a pelagic stage that persists until larvae are 30–34 mm SL (Kellermann 1989, p. 126). Because it is the derived state, we focus the remainder of our analysis on the large-mouth morph.
What is the incidence and geographic distribution of the large-mouth morph? The frequency in various populations is remarkably similar at 28–31% in McMurdo Sound (Eastman and De Vries 1997 and additional unpublished data) and 29% at King George/25 de Mayo Island (Piacentino and Barrera-Oro 2009). Although the frequency is unknown, the large-mouth morph has also been documented from Petermann Island, western Antarctic Peninsula (Piacentino and Barrera-Oro 2009), Cape Adare (Eastman and De Vries 1997) and Terra Nova Bay (Dr. Mario La Mesa, 1997–1998 Italian Expedition, personal communication). Because they have been discovered in both high and low Antarctic localities on opposite sides of the continent, it is likely that morphs are a general characteristic of T. newnesi and will eventually be found throughout the circum-Antarctic range of this species.
When in ontogeny does the large-mouth morph become recognizable? The large-mouth morphology is not obvious until specimens are juveniles of about 60 mm TL (Piacentino and Barrera-Oro 2009) and age 1 year. It would be desirable to refine this estimate by employing a large size range of large-mouth and typical morphs as subjects for statistical analysis, specifically for obtaining the slopes and intercepts of regression lines and the associated allometric data for gape width, upper jaw length and head depth against standard length. This procedure would establish a precise ontogenetic trajectory for the appearance of the proportionately larger head in the large-mouth morph and validate the hypothesis that the large-mouth morph is derived. In their morphometric and phylogenetic analysis of the ecomorphological trend toward habitation of the water column in ten nototheniid species, including seven species of Trematomus (but not T. newnesi), Klingenberg and Ekau (1996) used multivariate allometric techniques to determine that juveniles and adults share a common pattern of ontogenetic growth. They attributed the morphological differences between species to lateral transpositions of growth trajectories. This means that interspecific differences (i.e. the morphology associated with divergence into water column and benthic habitats) develop in embryonic and larval life, rather than considerably later in the growth period like those of the large-mouth morph of T. newnesi. The late ontogenetic appearance of large-mouth morph morphology also suggests there is a reduced likelihood that it has a genetic basis (Robinson and Schluter 2000, p. 68).
Do differential growth rates between the two morphs account for some of the observed phenotypic plasticity? The ratio of eye diameter to head length is smaller in the large-mouth morph (Piacentino and Barrera-Oro 2009). It has been observed that small relative eye size is linked to fast somatic growth in rainbow trout, Oncorhynchus mykiss (Pankhurst and Montgomery 1994). If this is also valid for the morphs of T. newnesi, the prediction is that a typical morph would be older than a large-mouth morph of the same size. We have age data obtained from scales for a few specimens of each morph, but our sample is too small to adequately test this hypothesis. This is a priority for future work.
Are there environmental stimuli that induce the development of the large-mouth morph? These have not been identified to date. Phenotypic plasticity is frequently associated with habitat instability and variability or with new and depauperate environments (Robinson and Schluter 2000; Agrawal 2001; West-Eberhard 2003; Schlichting 2004). Antarctic waters are as variable as any in the marine realm with considerable fluctuation in sea ice cover, light, benthic scour by icebergs, seasonal productivity and the presence of krill swarms, a major trophic resource that is patchily distributed (Gutt 2001; Thatje et al. 2005, 2008; Barnes and Conlan 2007). Nevertheless, we cannot say anything specific about environmental stimuli other than to note that the large-mouth morph should be able to consume larger prey.
What developmental mechanism is responsible for the morphological changes associated with the wide and deep head and large jaws of the large-mouth morph? Vertebrate skeletal tissues remodel adaptively in response to biomechanical loading stimuli and this phenomenon is well documented in the jaws of closely related fishes with different diets (Wimberger 1994). What we see in the heads and jaws of the T. newnesi morphs is likely the result of this mechanism. Although the oral and pharyngeal jaws are larger in the large-mouth morph (Fig. 4), both morphs have relatively delicate jaws and the teeth are weakly ankylosed to the jaws or held by thin ligament. Nothing is known about the masticatory forces generated during feeding in notothenioids but the ability to seize, engulf, transport and swallow larger prey would be facilitated by the larger gape and wider and deeper head of the large-mouth morph. Like many advanced bony fishes (Barton 2007), trematomids are suctorial feeders.
What maintains the large-mouth morphology in the population? There is adaptive value in the variation in feeding behaviour commonly seen in fishes with different diets, and this may involve both flexibility in feeding modes and switching of prey (Helfman 1994). Dietary plasticity is well documented in trematomids (Pakhomov 1998), including T. newnesi (La Mesa et al. 2000). Furthermore, it is possible that the degree of development of morphological plasticity is linked to the extent of behavioural plasticity in various fish taxa (Wimberger 1994; Skúlason et al. 1999). Under this scenario, the large-mouth morph is maintained and perpetuated in the population because they have sufficient behavioural plasticity to utilize a variety of prey including large and/or seasonally abundant items. The linkage between behavioural and morphological plasticity can influence the direction of selection on trophic morphology in a self-perpetuating cycle in individuals and populations, and this interaction has likely played an important role in the repeated appearance of benthic and limnetic forms of unrelated fishes (West-Eberhard 2003). This is especially likely to occur under extreme or patchy trophic conditions (Skúlason et al. 1999; West-Eberhard 2003), as might be seen in a highly seasonal inshore Antarctic marine habitat.
To sum up, in the absence of genetic data and of evidence of ecological diversification, the morphism in T. newnesi may be termed intraspecific ecophenotypic plasticity that involves the size of the head and jaws in three sympatric morphs. In Potter Cove the morphs do not exhibit ecological diversification based on measurements of buoyancy and, although a trophic basis for the polymorphism is possible, it has not yet been documented.
References
Agrawal AA (2001) Phenotypic plasticity in the interactions and evolution of species. Science 294:321–326
Barnes DKA, Conlan KE (2007) Disturbance, colonization and development of Antarctic benthic communities. Phil Trans R Soc B 362:11–38
Barrera-Oro E (2002) The role of fish in the Antarctic marine food web: differences between inshore and offshore waters in the southern Scotia Arc and west Antarctic Peninsula. Antarct Sci 14:293–309
Barrera-Oro E (2003) Analysis of dietary overlap in Antarctic fish (Notothenioidei) from the South Shetland Islands: no evidence of food competition. Polar Biol 26:631–637
Barrera-Oro ER, Casaux RJ (1996) Validation of age determination in Notothenia coriiceps by means of a tag/recapture experiment at Potter Cove, South Shetland Islands. Arch Fish Mar Res 43:205–216
Barrera-Oro ER, Piacentino GLM (2007) Feeding habits of juvenile Trematomus newnesi (Pisces, Nototheniidae) at Potter Cove, South Shetland Islands, Antarctica. Polar Biol 30:789–796
Barton M (2007) Bond’s biology of fishes, 3rd edn. Thomson Brooks/Cole, Belmont, CA
Casaux RJ, Mazzotta AS, Barrera-Oro ER (1990) Seasonal aspects of the biology and diet of nearshore nototheniid fish at Potter Cove, South Shetland Islands, Antarctica. Polar Biol 11:63–72
Casaux R, Barrera-Oro E, Baroni A, Ramón A (2003) Ecology of inshore notothenioid fish from the Danco Coast, Antarctic Peninsula. Polar Biol 26:157–165
De Witt TJ, Scheiner SM (eds) (2004) Phenotypic plasticity: functional and conceptual approaches. Oxford University Press, New York
De Witt HH, Heemstra PC, Gon O (1990) Nototheniidae. In: Gon O, Heemstra PC (eds) Fishes of the Southern Ocean. JLB Smith Institute of Ichthyology, Grahamstown, pp 279–331
Eastman JT (1993) Antarctic fish biology: evolution in a unique environment. Academic Press, San Diego
Eastman JT, De Vries AL (1982) Buoyancy studies of notothenioid fishes in McMurdo Sound, Antarctica. Copeia 2:385–393
Eastman JT, De Vries AL (1997) Biology and phenotypic plasticity of the Antarctic nototheniid fish Trematomus newnesi in McMurdo Sound. Antarct Sci 9:27–35
Eastman JT, Sidell BD (2002) Measurements of buoyancy for some Antarctic notothenioid fishes from the South Shetland Islands. Polar Biol 25:753–760
Gutt J (2001) On the direct impact of ice on marine benthic communities, a review. Polar Biol 24:553–564
Helfman GS (1994) Adaptive variability and mode choice in foraging fishes. In: Stouder DJ, Fresh KL, Feller RJ (eds) Theory and application in fish feeding ecology. University of South Carolina Press, Columbia, pp 3–17
Jonsson B, Jonsson N (2001) Polymorphism and speciation in Arctic charr. J Fish Biol 58:605–638
Kellermann A (1989) Catalogue of early life history stages of Antarctic notothenioid fishes. In: Kellermann A (ed) Identification key and catalogue of larval Antarctic fishes, BIOMASS Sci Ser No 10. Alfred-Wegener-Institut, Bremerhaven, pp 45–136
Klingenberg CP, Ekau W (1996) A combined morphometric and phylogenetic analysis of an ecomorphological trend: pelagization in Antarctic fishes (Perciformes: Nototheniidae). Biol J Linn Soc 59:143–177
Kock K-H, Kellermann A (1991) Reproduction in Antarctic notothenioid fish. Antarct Sci 3:125–150
La Mesa M, Vacchi M, Sertorio TZ (2000) Feeding plasticity of Trematomus newnesi (Pisces, Nototheniidae) in Terra Nova Bay, Ross Sea, in relation to environmental conditions. Polar Biol 23:38–45
Loeb VJ, Kellermann AK, Koubbi P, North AW, White MG (1993) Antarctic larval fish assemblages: a review. Bull Mar Sci 53:416–449
Near TJ, Jones CD, Eastman JT (2009) Geographic intraspecific variation in buoyancy within Antarctic notothenioid fishes. Antarct Sci 21:123–129
Ozgul A, Tuljapurkar S, Benton TG, Pemberton JM, Clutton-Brock TH, Coulson T (2009) The dynamics of phenotypic change and the shrinking sheep of St Kilda. Science 325:464–467
Pakhomov EA (1998) Feeding plasticity of the Antarctic fish Trematomus hansoni Boulenger, 1902 (Pisces: Nototheniidae): the influence of fishery waste on the diet. Polar Biol 19:289–292
Pankhurst NW, Montgomery JC (1994) Uncoupling of visual and somatic growth in the rainbow trout Oncorhynchus mykiss. Brain Behav Evol 44:149–155
Piacentino GLM, Barrera-Oro E (2009) Phenotypic plasticity in the Antarctic fish Trematomus newnesi (Nototheniidae) from the South Shetland Islands. Polar Biol 32:1407–1413
Robinson BW, Schluter D (2000) Natural selection and the evolution of adaptive genetic variation in northern freshwater fishes. In: Mousseau TA, Sinervo B, Endler JA (eds) Adaptive genetic variation in the wild. Oxford University Press, New York, pp 65–94
Robinson BW, Wilson DS (1994) Character release and displacement in fishes: a neglected literature. Am Nat 144:596–627
Robinson BW, Wilson DS (1996) Genetic variation and phenotypic plasticity in a trophically polymorphic population of pumpkinseed sunfish (Lepomis gibbosus). Evol Ecol 10:631–652
Schlichting CD (2004) The role of phenotypic plasticity in diversification. In: De Witt TJ, Scheiner SM (eds) Phenotypic plasticity: functional and conceptual approaches. Oxford University Press, Oxford, pp 191–200
Skúlason S, Noakes DLG, Snorrason SS (1989) Ontogeny of trophic morphology in four sympatric morphs of arctic charr Salvelinus alpinus in Thingvallavatn, Iceland. Biol J Linn Soc 38:281–301
Skúlason S, Snorrason S, Jónsson B (1999) Sympatric morphs, populations and speciation in freshwater fish with emphasis on arctic charr. In: Magurran AE, May RM (eds) Evolution of biological diversity. Oxford University Press, Oxford, pp 70–92
Thatje S, Hillenbrand C-D, Larter R (2005) On the origin of Antarctic marine benthic community structure. Trends Ecol Evol 20:534–540
Thatje S, Hillenbrand C-D, Mackensen A, Larter R (2008) Life hung by a thread: endurance of Antarctic fauna in glacial periods. Ecology 89:682–692
Tiedtke JE, Kock K-H (1989) Structure and composition of the demersal fish fauna around Elephant Island. Arch Fischereiwiss 39:143–169
Vacchi M, La Mesa M (1995) The diet of the Antarctic fish Trematomus newnesi Boulenger, 1902 (Nototheniidae) from Terra Nova Bay, Ross Sea. Antarct Sci 7:37–38
Van Houdt JKJ, Hellemans B, Van de Putte A, Koubbi P, Volckaert FAM (2006) Isolation and multiplex analysis of six polymorphic microsatellites in the Antarctic notothenioid fish, Trematomus newnesi. Mol Ecol Notes 6:157–159
West-Eberhard MJ (2003) Developmental plasticity and evolution. Oxford University Press, New York
Wimberger PH (1994) Trophic polymorphisms, plasticity, and speciation in vertebrates. In: Stouder DJ, Fresh KL, Feller RJ (eds) Theory and application in fish feeding ecology. University of South Carolina Press, Columbia, pp 19–43
Acknowledgments
We are very grateful to Carlos Bellisio, Luis Vila and Lic. E. Moreira for their help in field activities and laboratory procedures. In his capacity as a reviewer, Prof John C. Montgomery provided helpful comments concerning the influence of differential growth rates on phenotypic variation. J.T.E. was supported by US National Science Foundation grant ANT 04-36190.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Eastman, J.T., Barrera-Oro, E. Buoyancy studies of three morphs of the Antarctic fish Trematomus newnesi (Nototheniidae) from the South Shetland Islands. Polar Biol 33, 823–831 (2010). https://doi.org/10.1007/s00300-009-0760-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-009-0760-9