
Abstract
We consider the harvesting of a population in a stochastic environment whose dynamics in the absence of harvesting is described by a one dimensional diffusion. Using ergodic optimal control, we find the optimal harvesting strategy which maximizes the asymptotic yield of harvested individuals. To our knowledge, ergodic optimal control has not been used before to study harvesting strategies. However, it is a natural framework because the optimal harvesting strategy will never be such that the population is harvested to extinction—instead the harvested population converges to a unique invariant probability measure. When the yield function is the identity, we show that the optimal strategy has a bang–bang property: there exists a threshold \(x^*>0\) such that whenever the population is under the threshold the harvesting rate must be zero, whereas when the population is above the threshold the harvesting rate must be at the upper limit. We provide upper and lower bounds on the maximal asymptotic yield, and explore via numerical simulations how the harvesting threshold and the maximal asymptotic yield change with the growth rate, maximal harvesting rate, or the competition rate. We also show that, if the yield function is \(C^2\) and strictly concave, then the optimal harvesting strategy is continuous, whereas when the yield function is convex the optimal strategy is of bang–bang type. This shows that one cannot always expect bang–bang type optimal controls.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Many species of animals like whales, elephant seals, bisons and rhinoceroses, are at risk of being harvested to extinction (Gulland 1971; Reiter et al. 1981; Ludwig et al. 1993; Primack 2006). Excessive harvesting has already led to both local and gobal extinctions of species (Lande et al. 1995). In fact, a significant percentage of the endangered birds and mammals of the world are threatened by harvesting, hunting or other types of overexploitation (Lande et al. 1995), and there are similar problems for many species of fish (Hutchings and Reynolds 2004). This is why harvesting strategies have to be carefully chosen. After significant harvests, it takes time for the harvested population to get back to the pre-existing level. Moreover, the harvested population fluctuates randomly in time due to environmental stochasticity. As a result, an overestimation of the ability of the population to rebound can lead the harvester to overharvest the population to extinction (Lande et al. 1995). A less common but nevertheless important problem is an insufficient rate of harvesting. Because of instraspecific competition, the population is bounded in a specific environment, so an extraction rate that is too low would lead to a loss of precious resources. For the same reason, choosing an efficient extraction strategy for valuable species is important (Kokko 2001).
We present a stochastic model of population harvesting and find the optimal harvesting strategy that maximizes the asmptotic yield of harvested individuals. We consider a novel framework, the one of optimal ergodic harvesting. This is based on the theory of ergodic control (Arapostathis et al. 2012). In most stochastic models that exist in the literature, for example Lande et al. (1995), Alvarez and Shepp (1998) and Lungu and Øksendal (1997), the population is either assumed to become extinct in finite time, or it can end up being harvested to extinction. In our framework, if the population goes extinct under some harvesting strategy, the asymptotic yield is 0 and therefore this strategy cannot be optimal. If one wants to ensure that harvested species are preserved, this framework is a natural candidate. Our aim is to present a theory of optimal harvesting that includes the risks of extinction from both environmental noise and harvesting. We assume that the population is homogeneous and can be described by a one dimensional diffusion. The harvesting rate is assumed to be bounded, as infinite harvesting rates would imply an unlimited harvesting capacity, something that is clearly not realistic.
In most cases, environmental noise can be introduced in the system by transforming differential equations into stochastic differential equations (SDE). Such techniques require dealing with significant mathematical difficulties, but their use is not just a case of honoring generality. First, there are direct effects of stochasticity on the predictions of the model, and the parameters quantifying it show up in the results. Second, any realistic biological system will depend on environmental variables that are not, or cannot be, accounted for. The role of stochasticity is to ensure that the solutions proposed are robust to such omissions. For example, if avoiding extinction is important, deterministic models can give misleading solutions even when their parameters are corrected for noise (Smith 1978). The transformation to SDE works especially well when the environmental fluctuations are small and there is no chaos (Lande et al. 1995). We focus on models with environmental stochasticity and neglect the demographic stochasticity which arises from the randomness of birth and death rates of each indiviual of a population. Throughout the paper we assume that environmental stochasticity mainly affects the growth rate of the population (see Turelli 1977; Beddington and May 1977; May et al. 1978; Leigh 1981; Braumann 2002; Gard 1988; Evans et al. 2015, 2013; Schreiber et al. 2011; Hening and Nguyen 2018a for more details). For computational tractability and for clarity of exposition, we look at a one-dimensional model. Nevertheless, our framework works for any model that can be written as a system of stochastic differential equations (satisfying some mild assumptions—see Arapostathis et al. 2012).
A major limitation of existing models in the literature is the dependence of the optimal solutions on parameters that are hard to quantify. For example, in Lande et al. (1995) the level at which the population becomes extinct—the minimal viable population—must be assumed; without it the yields become infinite. In Alvarez and Shepp (1998) the yield must be time discounted to avoid maximizing over yield infinities, and this requires providing a time value for resources. The minimal viable population is a difficult scientific question (Shaffer 1981; Traill et al. 2007), and the time value of yields is a difficult economics and policy question, because it implies the comparison of the utility of present and future generations (Drèze and Stern 1987). In contrast, our model sidesteps the issue by assuming no time preference—and therefore no bias towards extracting in the present, and resolves the problem of maximizing over infinite yields naturally by looking at asymptotic behavior.
A particular case of our model was studied in Abakuks and Prajneshu (1981).Footnote 1 The authors limited themselves to the analysis of harvesting strategies that were of bang–bang type. In Abakuks (1979), one of the co-authors in Abakuks and Prajneshu (1981) proved that an optimal gathering strategy was necessarily of a bang–bang type in a continuous time Markov chain model, making use of the simplifying assumption of a finite state space. Here, instead, we look at very general possible harvesting strategies in a continuous state stochastic model, and show that the optimal one is of bang–bang type. Our contribution is therefore twofold. We generalize the setting of Abakuks and Prajneshu (1981) significantly by looking at very general density-dependent growth rates, not just the logistic case. Moreover, we prove what the authors of Abakuks and Prajneshu (1981) intuited, namely that the optimal strategy is of bang–bang type; and furthermore that this is true for the larger class of convex yield functions.
Stochastic optimal control applications are common in the finance literature. Following the seminal contributions of Merton (1969, 1971), objective functions that are integrals of time discounted instantaneous utility flows are now standard. The crucial simplifying assumption is that of time-additive total utility. The utility flow usually depends on consumption flows, and therefore indirectly on other variables and stochastic constraints. With the time-additive utility assumption, our general yield function can also be interpreted as an instantaneous utility function dependent on yield, and our objective function can be the asymptotic expected utility flow dependent on yield. Because a population stock cannot grow indefinitely in our biological model, we diverge from the general finance literature, where financial returns do not usually depend on the size of the holdings of an individual.
Finally, we generalise a result from one of the stochastic models in Smith (1978), where the equivalent to our yield function has a specific simple form. We show that, when the yield function is weakly convex, the optimal control is bang–bang. However, if the yield function is strictly concave, then the optimal harvesting strategy has to be continuous, in contrast to the bang–bang type optimal strategy we find for a linear yield function. This generalization is useful for economic welfare analysis (a more general form of cost-benefit analysis), which typically relies on a concave utility function, equivalent to the concave yield function herein. In economic models, concavity is assumed to model risk aversion [see Mas-Colell et al. (1995, Proposition 6.C.1) for justification], and for the convenience of interior solutions to maximisation problems. Concave utility leads to a trade-off between risk and returns in asset choice (Merton 1971), so the connection between yield concavity and strategy continuity mentioned above is suggestive of risk management. However, risk management interpretations from the finance literature are not directly applicable here. First, financial asset returns are assumed reasonably to not be decreasing in the asset value owned by investors.Footnote 2 Moreover, the risk-return trade-off is captured in models with choice between at least two assets with different risk profiles.Footnote 3 If anything, finding a bang–bang optimal strategy when yield is linear is more related to finding corner solutions in maximisation problems with linear utility. A bang–bang strategy uses one of the two extremes of the harvesting rate, depending on the momentary population stock.
The rest of the paper is organized as follows. In Sect. 2 we introduce our model and results. We prove that, if the population in the absence of harvesting survives, the yield function is the identity and the harvesting rate is bounded above by some number \(M>0\), then the optimal strategy is always a bang–bang type solution: there exists an \(x^*>0\) such that one does not harvest if the current population size lies in the interval \(\left[ 0,x^*\right] \) and harvests at the maximal possible rate, M, if the current population size lies in the interval \(\left( x^*,\infty \right) \). The proofs of the above results are collected in Appendix A. In Sect. 4 we apply our results to the special setting of the logistic Verhulst–Pearl model. In Sect. 3 (proofs in Appendix B) we show that if the yield function is strictly concave, the optimal harvesting strategy is continuous, and when the yield function is more generally weakly convex, the optimal strategy is bang–bang.
Finally, in Sect. 5 we offer some numerical simulations that show how the optimal harvesting strategies and optimal asymptotic change with respect to the parameters of the model. We also provide a discussion of our results.
2 Optimal ergodic harvesting
We consider a population whose density \({\tilde{X}}(t)\) at time \(t\ge 0\), in the absence of harvesting, follows the stochastic differential equation (SDE)
where \((B(t))_{t\ge 0}\) is a standard one dimensional Brownian motion. This describes a population \({\tilde{X}}\) with per-capita growth rate given by \(\mu (x)>0\) when the density is \(\tilde{X}=x\). The infinitesimal variance of fluctuations in the per-capita growth rate is given by \(\sigma ^2\).
The following is a standing assumption throughout the paper.
Assumption 2.1
The function \(\mu :[0,\infty )\rightarrow \mathbb {R}\) satisfies:
-
\(\mu \) is locally Lipschitz.
-
\(\mu \) is decreasing.
-
As \(x\rightarrow \infty \) we have \(\mu (x)\rightarrow -\infty \).
-
The function \(p(x):=x\mu (x)\) has a unique maximum.
-
There is no interval \((u,v)\subset \mathbb {R}_+\) such that \(p(\cdot )\) is constant on (u, v).
The behavior of (2.1) is not hard to study. In the particular case when \(\mu (x)=\overline{\mu }- \kappa x\) see Evans et al. (2015) and Dennis and Patil (1984). The methods there can be easily adapted to our setting. Alternatively, one could use the general results from Hening and Nguyen (2018a). The process \(\tilde{X}\) does not reach 0 or \(\infty \) in finite time and the stochastic growth rate \(\mu (0)-\frac{\sigma ^2}{2}\) determines the long-term behavior in the following way:
-
If \(\mu (0)-\frac{\sigma ^2}{2}>0\) and \( \tilde{X}(0)=x>0\), then \((\tilde{X}(t))_{t\ge 0}\) converges weakly to its unique invariant probability measure \(\nu \) on \((0,\infty )\).
-
If \(\mu (0)-\frac{\sigma ^2}{2}<0\) and \( \tilde{X}(0)=x>0\), then \(\lim _{t\rightarrow \infty } \tilde{X}(t)=0\) almost surely.
We let \(\mathbb {R}_+:=[0,\infty )\) and \(\mathbb {R}_{++}:=(0,\infty )\) throughout the paper.
Assume that the population is harvested at time \(t\ge 0\) at the stochastic rate\(h(t)\in U:=[0,M]\) for some fixed \(M>0\). Adding the harvesting to (2.1) yields the SDE
A stochastic process \((h(t))_{t\ge 0}\) taking values in U is said to be an admissible strategy if \((h(t))_{t\ge 0}\) is adapted to the filtration \((\mathcal {F}_t)_{t\ge 0}\) generated by the Brownian motion \((B(t))_{t\ge 0}\). Let \(\mathfrak {U}\) be the class of admissible strategies. An important subset of \(\mathfrak {U}\) is the class \(\mathfrak U_{sm}\) of stationary Markov strategies, that is, admissible strategies of the form \(h(t)=v(X(t))\) where \(v:\mathbb {R}_{++}\mapsto U\) is a measurable function. By abuse of terminology, we often refer to the map \(v(\cdot )\) as the stationary Markov strategy. Using a stationary Markov strategy \(v(\cdot )\), (2.2) becomes
Remark 2.1
The sigma algebra \(\mathcal {F}_t\) gives one the information available from time 0 to time t. An admissible harvesting strategy is therefore a strategy which can take into account all the information from the start of the harvesting to the present. These strategies are much more general than constant strategies. Stationary Markov strategies are the harvesting strategies which only depend on the present state of the population density.
We associate with X(t) the family of generators \(({\mathcal L}_u)_{u\in [0,M]}\) defined by their action on \(C^2\) functions with compact support in \(\mathbb {R}_{++}\) as
We will call \(\Phi :\mathbb {R}_+\rightarrow \mathbb {R}_+\) a yield function if the following assumption holds.
Assumption 2.2
The function \(\Phi :\mathbb {R}_+\rightarrow \mathbb {R}_+\) satisfies:
-
\(\Phi \) is continuous.
-
\(\Phi (0)=0\).
-
\(\Phi \) has subpolynomial growth that is, there is \(n \in \mathbb {N}\) such that \(\frac{\Phi (x)}{x^n} \rightarrow 0\) for \(x\rightarrow \infty \).
Our aim is to find the optimal strategy h(t) that almost surely maximizes the asymptotic yield
In other words we want to find v such that, for any initial population size \(X(0)=x>0\), we have with probability 1 that
We note that many of the existing models that look at the optimal harvesting of a population in a stochastic environment (Lungu and Øksendal 1997; Alvarez and Shepp 1998; Lande et al. 1995) assume that the yield function \(\Phi \) is the identity i.e. \(\Phi (x)=x, x\ge 0\). This assumption is not always justifiable (see Alvarez 2000) and as such we present in Sect. 3 results for more general functions \(\Phi \).
Remark 2.2
We note that if X has an invariant probability measure \(\pi \) on \(\mathbb {R}_{++}\), then for any \(X(0)=x>0\) almost surely
In particular, if X goes extinct, that is, for any \(X(0)=x>0\) we have with probability 1
then the only invariant ergodic measure of X on \(\mathbb {R}_+\) is \(\delta _0\) the point mass at 0, and hence, we get that with probability 1
Our method for maximizing the asymptotic yield forces the optimal harvesting to be such that the population persists.
Remark 2.3
By Arapostathis et al. (2012, Theorems 2.2.2 and 2.2.12), the controlled systems (2.2) and (2.3) have unique local solutions on \(\mathbb {R}_{++}\) for any admissible control h(t) and stationary Markov control v respectively. Note that one can find \(N>0\) large enough such that
With this fact in hand, we can use the arguments from Khasminskii (2012, Theorem 3.5), to obtain the existence of global solutions on \(\mathbb {R}_{++}\) of (2.2) and (2.3). In particular we get that
The main result of the paper is the following.
Theorem 2.1
Assume that \(\Phi (x)=x, x\in (0,\infty )\) and that the population survives in the absence of harvesting, that is \(\mu (0)-\frac{\sigma ^2}{2}>0\). Furthermore assume that the drift function \(\mu (\cdot )\) satisfies Assumption 2.1. The optimal control (the optimal harvesting strategy) v has the bang–bang form
for a unique \(x^*\in (0,\infty )\). Furthermore, we have the following upper bound for the optimal asymptotic yield
3 Continuous versus bang–bang optimal harvesting strategies
As showcased in Theorem 2.1, when \(\Phi \) is the identity function the optimal harvesting strategy is of bang–bang type. In Appendix B we prove the following result.
Theorem 3.1
Suppose Assumption 2.1 holds and the yield function satisfies
-
(1)
\(\Phi \in C^2(\mathbb {R}_+)\),
-
(2)
\(\Phi \) is strictly concave.
Then the optimal harvesting strategy is continuous and given by
Furthermore, the HJB equation for the system becomes
Remark 3.1
We cannot find the exact form of the optimal harvesting strategies in this case. Note that in Theorem 2.1 we have \(\Phi (x)=x\) which is not strictly concave nor strictly convex.
Intuitively, this is not unlike maximising a strictly concave objective function under a linear constraint. The optimal choice usually moves smoothly over the domain as the direction of the constraint changes. However, when the objective function is weakly convex, e.g. linear, the optimum will jump on the allowed interval.
Here, we show that if the yield function is weakly convex, the optimal control is bang–bang. The optimal strategy has a similar form to the one for linear yield, if a further assumption on the joint rates of change of the population growth rate and the yield function is made.
Theorem 3.2
Assume that \(\Phi :\mathbb {R}_+\rightarrow \mathbb {R}_+\) is weakly convex, \(\Phi \) grows at most polynomially, \(\Phi \in C^1(\mathbb {R_+})\) and the population survives in the absence of harvesting, that is \(\mu (0)-\frac{\sigma ^2}{2}>0\). Furthermore assume that the drift function \(\mu (\cdot )\) satisfies the following modification of Assumption 2.1:
-
(i)
\(\mu \) is locally Lipschitz.
-
(ii)
\(\mu \) is decreasing.
-
(iii)
As \(x\rightarrow \infty \) we have \(\mu (x)\rightarrow -\infty \).
-
(iv)
The function
$$\begin{aligned} G(x)= \Phi (xM)\left( 1-\frac{2}{\sigma ^2}\mu (x)\right) - xM\Phi '(xM) \end{aligned}$$(3.2)has a unique extreme point in \((0,\infty )\) which is a minimum, and is not constant on any interval \((u,v)\subset \mathbb {R}_+\).
If the assumptions (i)–(iii) hold, the optimal control has a bang–bang form (i.e., the harvesting rate is either 0 or the maximal M). If assumptions (i)–(iv) hold, the optimal harvesting strategy v has a bang–bang form with one threshold
for some \(x^*\in (0,\infty )\).
4 The logistic case: \(\mu (x)=\overline{\mu }-\kappa x\)
Throughout this section we provide a thorough analysis of the logistic Verhulst–Pearl model. As such, we will assume that the growth rate is \(\mu (x)=\overline{\mu }-\kappa x\) for positive constants \(\overline{\mu }, \kappa >0\). It is clear that this \(\mu (\cdot )\) satisfies Assumption 2.1. If we harvest according to a constant strategy\(\ell >0\) then the SDE (2.3) becomes
It is then easy to see that, as long as \(\overline{\mu }-\ell -\frac{\sigma ^2}{2}>0\), the asymptotic yield is
We can maximize this yield \(L(\ell )\), which is quadratic in \(\ell \). The maximum will be at
and the maximal asymptotic yield (among constant harvesting strategies) is
Note that \(L(\ell ^*)\) is also called maximum sustainable yield (MSY) in the literature. Since \(x\mu (x)=\overline{\mu }x-\kappa x^2\) we note that
Combining this with (2.7) one sees that the optimal asymptotic yield \(\rho ^*\) satisfies
Note that Theorem 2.1 does not give us information about \(x^*\), the point at which one starts harvesting.
One possible strategy to find out more information about \(x^*\) is the following: Look at controls of bang–bang type that have a threshold at \(\eta \) and then maximize over all possible \(\eta \). This will then give us a way of finding \(x^*\). Let \(w(x;\eta )\) be the harvesting strategy
For this control w our diffusion (2.3) (with \(h\equiv w\)) is of the form
for
and
Standard diffusion theory shows (see Hening and Kolb 2018; Borodin and Salminen 2012) that the boundary 0 is natural and the boundary \(\infty \) is entrance for the process X from (4.2). As a result, when \(\mu -\frac{\sigma ^2}{2}>0\), one can show using Borodin and Salminen (2012) that the density \(\rho :(0,\infty )\rightarrow (0,\infty )\) of the invariant measure \(\pi \) is of the form
where \(C_1\) is a normalizing constant given by
In this case the harvesting yield is
By Theorem 2.1 the point \(x^*\) has to satisfy:
It is clear that H is differentiable, that \(x^*\) exists and satisfies \(x^*\in (0,\infty )\). Therefore, \(x^*\) is a solution of
The condition above can be restated as an equation involving incomplete gamma functions. We were not able to prove analytically that (4.5) has a unique solution. Berg and Pedersen (2006, 2008) show possible analytical methods that can be applied to such equations in a simple case. However, numerical experiments that we have done support this conjecture (see Fig. 1).

Typical shape of the asymptotic yield function H(x) as a function of the harvesting threshold x, where one begins to harvest. Here for \(\sigma ^2=1\) and \(M=\overline{\mu }=\kappa =1\)
Conjecture 4.1
There exists a unique \(x^*\in (0,\infty )\) such that \(H'(x^*)=0\). Furthermore, the optimal harvesting strategy is given by
5 Discussion and future research
We have analysed a population whose dynamics evolves according to generalization of the logistic Verhulst–Pearl model in a stochastic environment, but subjected to strategic harvesting. The rate at which the population gets harvested is bounded above by a constant \(M>0\), and the harvested infinitesimal amount is proportional to the current size of the population. We show that the harvesting strategy v, which describes the harvesting rate and is chosen to maximize the asymptotic harvesting yield
is of bang–bang type, i.e. there exists \(x^*>0\) such that

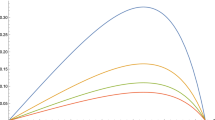
Graph of the maximal asymptotic yield H(x) as a function of the harvesting threshold x for different values of the growth rate \(\overline{\mu }\). We take \(\sigma ^2=1\), \(M=\kappa =1\), and \(\overline{\mu }=1\) (blue), \(\overline{\mu }=1.5\) (orange), \(\overline{\mu }=2\) (green), \(\overline{\mu }=2.5\) (red), and \(\overline{\mu }=3\) (purple) (color figure online)
5.1 Logistic Verhulst–Pearl
In the particular case when \(\mu (x)=\overline{\mu }- \kappa x\), we can give more information about \(x^*\) as follows: The harvesting yield function \(H(\eta )\) is determined, by letting the jump in the bang–bang control be at \(\eta \). That is, we look at the yield when the control is
The typical behavior of the point \(x^*\) where H is maximized and of \(H(x^*)\) as the parameters \(\overline{\mu }, \kappa \) and M change was analyzed numerically and is presented in Figs. 2, 3 and 4, with the normalization \(\sigma ^2=1\).
We note from numerical experiments that increasing the growth rate \(\overline{\mu }\) increases the threshold \(x^*\) at which one should start optimally harvesting (Fig. 2). This is an intuitive result, since an increased growth rate increases the maximal equilibrium value of the population in the equivalent deterministic growth model with competition (Smith 1978). Therefore, it should also increase asymptotic harvesting yield, as well as the point at which harvesting should start. Moreover, higher growth rates make the population get faster to the point \(x^*\) where one starts harvesting, reducing the cost of a delay.
If one increases the maximal harvesting rate M then the harvesting threshold \(x^*\) is also increased (Fig. 3). This also makes sense because if \(\overline{\mu }-\frac{\sigma ^2}{2}-M<0\), then a population with constant harvesting rate M will go extinct almost surely. An increase in the harvesting threshold \(x^*\) is necessary to make sure that there is no extinction. Moreover, as M gets larger one can wait longer to start harvesting. With a larger maximal rate available, there is less chance that there will be losses because the population overshoots the optimal extraction point. Similarly, increasing the harvesting rate M also increases the maximal asymptotic harvesting yield, for the obvious reason that there is better control on the population level and therefore extraction can happen closer to the optimal level.
In contrast, if one increases the intraspecific competition rate \(\kappa \), then the harvesting threshold decreases (Fig. 4). The equilibrium value of the population in the equivalent deterministic model (Smith 1978) decreases with \(\kappa \), and as a result so does the extraction rate. Evidently, even in the stochastic model, if competition is very strong the population cannot spend much time at high densities, and therefore one has to start harvesting early. An increase in \(\kappa \) will also decrease the maximal asymptotic harvesting yield.

Graph of the maximal asymptotic yield H(x) as a function of the harvesting threshold x for different values of the maximal harvesting rate M. We take \(\sigma ^2=1\), \(\overline{\mu }=\kappa =1\), and \(M=0.1\) (blue), \(M=0.2\) (orange), \(M=0.5\) (green), \(M=1\) (red), \(M=2\) (purple), and \(M=5\) (brown) (color figure online)

Graph of the maximal asymptotic yield H(x) as a function of the harvesting threshold x for different values of the intra-competition rate. We take \(\sigma ^2=1\), \(M=\overline{\mu }=1\), and \(\kappa =1\) (blue), \(\kappa =2\) (orange), \(\kappa =3\) (green), \(\kappa =4\) (red), and \(\kappa =5\) (purple) (color figure online)
We are able to prove that the maximal asymptotic yield \(\rho ^*\) satisfies the inequality
In particular, the bang–bang optimal strategy has a higher asymptotic yield than the optimal constant harvesting strategy. Moreover, the bang–bang optimal strategy gives a lower asymptotic yield than the optimal constant harvesting strategy in the absence of noise. This means that the analysis of the more complex stochastic model was fruitful, recommending a qualitatively different strategy. Moreover, environmental fluctuations decrease the maximal asymptotic yield and, because the correction is negative, protecting a population from extinction requires a careful measurement of natural fluctuations when designing optimal harvesting. When environmental stochasticity was not taken into account, harvesting often lead populations to extinction (Lande et al. 1995).
Real populations do not evolve in isolation. As a result, ecology is concerned with understanding the characteristics that allow species to coexist. Harvesting can disturb the coexistence of species. In future research we intend to tackle multi-dimensional analogues of the setting treated in the current article. Natural models for which one can add harvesting would be predator-prey food chains (Gard and Hallam 1979; Gard 1984; Hening and Nguyen 2018b, c; Tyson and Lutscher 2016), more general Kolmogorov systems (Schreiber et al. 2011; Hening and Nguyen 2018a) and structured populations where there can be asymmetric harvesting (Evans et al. 2013, 2015; Hening et al. 2018; Roth and Schreiber 2014; Benaïm and Schreiber 2009; Schreiber and Ryan 2011). In the multi-dimensional setting the Hamilton–Jacobi–Bellman (HJB) equation becomes a PDE and the analysis becomes significantly more complex. New tools will have to be developed to tackle these problems.
Above we have imposed a bound on the extraction rate, M. This was because it is a realistic feature, but it was also practical for the analysis. Nevertheless, it is interesting to consider the case when the extraction rate is unbounded. A practical model with no extraction limit corresponds to having unlimited control over a target population, which is sometimes the case. Such a model would have the benefit of not requiring a nuisance parameter that may be hard to determine.
5.2 Concave and convex yield functions
We have also studied the more general case involving concave and convex yield functions. When the yield function is strictly concave, it was shown that the optimal control is not bang–bang, but continuous in the population parameter. Vice-versa, when the yield function is weakly convex, we have shown that the optimal control is necessarily bang–bang. Moreover, if a certain further assumption on the relative rate of growth of \(\mu \) and \(\Phi \) holds, we can also show that the bang–bang optimal control has a single threshold \(x^*\) where the extraction rate goes from 0 to M—as in the linear special case.
This generalization allows us to think of applications of population harvesting where the yield function is in fact a utility function, or some other more general social welfare measure.
5.3 Unbounded harvesting
If we allow for general, possibly unbounded, harvesting we would have to study the Skorokhod SDE
where \((Z_t)_{t\ge 0}\) is supposed to be non-negative, increasing, right-continuous and adapted to \((\mathcal {F}_t)_{t\ge 0}\)—we denote the set of all such strategies by A. Then the problem is to maximize the asymptotic yield, i.e. find
We want to find the harvesting strategy \((Z_t^*)_{t\ge 0}\in A\), which we call the optimal harvesting strategy, such that
The analysis above, for the bounded harvesting rate, determined that the optimal strategy has a bang–bang property, where extraction is maximal after some cut-off. This suggests that raising the maximum would not change the bang–bang property, but determining that result required a bounded extraction rate. Thinking of the limiting behavior of the yield function above shows the difficulty: as \(M \rightarrow \infty \), the density of the distribution above the cut-off \(x^*\) goes to 0 (see (4.3)). The conjectured optimal solution is akin to having a reflective boundary at \(x^*\), and the yield is determined by the time spent close to the boundary.
Conjecture 5.1
Assume that the population survives in the absence of harvesting i.e. \(\mu -\frac{\sigma ^2}{2}>0\). The optimal extraction strategy \((Z^*_t)_{t\ge 0}\) has the form
for some \(x^*\in (0,\infty )\), where \(L(t,x^*)\) is the local time at \(x^*\) of the process \(\tilde{X}\) from (5.1).
This conjecture is supported by the results from Alvarez and Shepp (1998) where the authors study the maximization of the discounted yield
and \(\tau :=\inf \{t\ge 0: \tilde{X}_t=0\}\) is the extinction time. It is shown in Alvarez and Shepp (1998) that the optimal harvesting strategy is of the form (5.2). One possible approach to prove Conjecture 5.1 would be to use the results from Alvarez and Shepp (1998) and then let the discount factor r go to 0.
Notes
We thank the anonymous referee who has brought the paper to our attention.
The assumption may not apply in models with large institutional investors.
An ecological model extension that would link this literature to our model would consider optimal extraction policy to maximise a time discounted concave total-yield function when there are at least two populations, situated in different environments with no growth limitation.
\(|\mu (x) - \mu (0)| < M|x|\) for some real \(M>0\) as \(\mu \) locally Lipschitz by asssumption. Therefore \(|g(x) - g(0)| < Mx^2\), so g differentiable at 0. Moreover, \(g'(0) = - \mu (0)\), and \(\mu (0)>0\) by assumption, so \(x_\iota \ne 0\).
References
Abakuks A (1979) An optimal hunting policy for a stochastic logistic model. J Appl Probab 16(2):319–331
Arapostathis A, Borkar V S, Ghosh M K (2012) Ergodic control of diffusion processes. Cambridge University Press, Cambridge
Alvarez LHR (2000) Singular stochastic control in the presence of a state-dependent yield structure. Stoch Process Appl 86(2):323–343
Abakuks A, Prajneshu (1981) An optimal harvesting policy for a logistic model in a randomly varying environment. Math Biosci 55(3–4):169–177
Alvarez LHR, Shepp LA (1998) Optimal harvesting of stochastically fluctuating populations. J Math Biol 37(2):155–177
Beddington JR, May RM (1977) Harvesting natural populations in a randomly fluctuating environment. Science 197(4302):463–465
Berg C, Pedersen HL (2006) The Chen–Rubin conjecture in a continuous setting. Methods Appl Anal 13(1):63–88
Berg C, Pedersen HL (2008) Convexity of the median in the gamma distribution. Ark Mat 46(1):1–6
Braumann CA (2002) Variable effort harvesting models in random environments: generalization to density-dependent noise intensities. Math Biosci 177(178):229–245 (Deterministic and stochastic modeling of biointeraction (West Lafayette, IN, 2000))
Benaïm M, Schreiber SJ (2009) Persistence of structured populations in random environments. Theor Popul Biol 76(1):19–34
Borodin AN, Salminen P (2012) Handbook of Brownian motion-facts and formulae. Birkhäuser, Basel
Dennis B, Patil GP (1984) The gamma distribution and weighted multimodal gamma distributions as models of population abundance. Math Biosci 68(2):187–212
Drèze J, Stern N (1987) The theory of cost-benefit analysis. Handb Pub Econ 2:909–989
Evans SN, Hening A, Schreiber SJ (2015) Protected polymorphisms and evolutionary stability of patch-selection strategies in stochastic environments. J Math Biol 71(2):325–359
Ethier S N, Kurtz T G (2009) Markov processes: characterization and convergence. Wiley, New York
Evans SN, Ralph PL, Schreiber SJ, Sen A (2013) Stochastic population growth in spatially heterogeneous environments. J Math Biol 66(3):423–476
Gard TC (1984) Persistence in stochastic food web models. Bull Math Biol 46(3):357–370
Gard TC (1988) Introduction to stochastic differential equations. M. Dekker, New York
Gard TC, Hallam TG (1979) Persistence in food webs. I. Lotka-Volterra food chains. Bull Math Biol 41(6):877–891
Gulland JA (1971) The effect of exploitation on the numbers of marine animals. In: Proceedings of the advanced study institute on dynamics of numbers in populations, pp 450–468
Hening A, Kolb M (2018) Quasistationary distributions for one-dimensional diffusions with singular boundary points (submitted). arXiv:1409.2387
Hening A, Nguyen D (2018) Coexistence and extinction for stochastic Kolmogorov systems. Ann Appl Probab. arXiv:1704.06984
Hening A, Nguyen D (2018) Persistence in stochastic Lotka-Volterra food chains with intraspecific competition (preprint). arXiv:1704.07501
Hening A, Nguyen D (2018) Stochastic Lotka-Volterra food chains. J Math Biol. arXiv:1703.04809
Hening A, Nguyen D, Yin G (2018) Stochastic population growth in spatially heterogeneous environments: the density-dependent case. J Math Biol 76(3):697–754
Hutchings JA, Reynolds JD (2004) Marine fish population collapses: consequences for recovery and extinction risk. AIBS Bull 54(4):297–309
Khasminskii R (2012) Stochastic stability of differential equations. In: Rozovskiĭ B, Glynn PW (eds) Stochastic modelling and applied probability, vol 66, 2nd edn. Springer, Heidelberg (With contributions by G. N. Milstein and M. B. Nevelson)
Kokko H (2001) Optimal and suboptimal use of compensatory responses to harvesting: timing of hunting as an example. Wildl Biol 7(3):141–150
Leigh EG (1981) The average lifetime of a population in a varying environment. J Theor Biol 90(2):213–239
Lande R, Engen S, Saether B-E (1995) Optimal harvesting of fluctuating populations with a risk of extinction. Am Nat 145(5):728–745
Ludwig D, Hilborn R, Walters C (1993) Uncertainty, resource exploitation, and conservation: lessons from history. Ecol Appl 3:548–549
Lungu EM, Øksendal B (1997) Optimal harvesting from a population in a stochastic crowded environment. Math Biosci 145(1):47–75
May RM, Beddington JR, Horwood JW, Shepherd JG (1978) Exploiting natural populations in an uncertain world. Math Biosci 42(3–4):219–252
Mas-Colell A, Whinston MD, Green JR (1995) Microeconomic theory, vol 1. Oxford University Press, New York
Merton RC (1969) Lifetime portfolio selection under uncertainty: the continuous-time case. Rev Econ Stat 51:247–257
Merton RC (1971) Optimum consumption and portfolio rules in a continuous-time model. J Econ Theory 3(4):373–413
Primack RB (2006) Essentials of conservation biology. Sinauer Associates, Sunderland
Reiter J, Panken KJ, Le Boeuf BJ (1981) Female competition and reproductive success in northern elephant seals. Anim Behav 29(3):670–687
Roth G, Schreiber SJ (2014) Persistence in fluctuating environments for interacting structured populations. J Math Biol 69(5):1267–1317
Schreiber SJ, Benaïm M, Atchadé KAS (2011) Persistence in fluctuating environments. J Math Biol 62(5):655–683
Shaffer ML (1981) Minimum population sizes for species conservation. Bioscience 31(2):131–134
Smith JB (1978) An analysis of optimal replenishable resource management under uncertainty. Digitized Theses, Paper 1074
Schreiber SJ, Ryan ME (2011) Invasion speeds for structured populations in fluctuating environments. Theor Ecol 4(4):423–434
Traill LW, Bradshaw CJA, Brook BW (2007) Minimum viable population size: a meta-analysis of 30 years of published estimates. Biol Conserv 139(1):159–166
Tyson R, Lutscher F (2016) Seasonally varying predation behavior and climate shifts are predicted to affect predator-prey cycles. Am Nat 188(5):539–553
Turelli M (1977) Random environments and stochastic calculus. Theor Popul Biol 12(2):140–178
Acknowledgements
We thank two anonymous referees for very insightful comments and suggestions that led to major improvements.
Author information
Authors and Affiliations
Corresponding author
Additional information
Dang H. Nguyen was in part supported by the National Science Foundation under Grant DMS-1207667 and is also supported by an AMS Simons travel grant. Tak Kwong Wong was in part supported by the HKU Seed Fund for Basic Research under the Project Code 201702159009, and the Start-up Allowance for Croucher Award Recipients.
Appendices
Appendix A: Proofs
In this appendix we present the framework of ergodic optimal control and prove the main results of our paper.
For any \(v\in \mathfrak U_{sm}\), denote the unique invariant probability measure of X(t) on \(\mathbb {R}_{++}\) by \(\pi _v\) if it exists. Define
Let \(p>0\). Since \(\lim _{x\rightarrow \infty }\mu (x)=-\infty \), there exist constants \( k_{1p}, k_{2p}>0\) such that
By Dynkin’s formula
Thus,
As a result, the family of occupation measures
is tight. If X(t) has an invariant probability measure on \(\mathbb {R}_{++}\), then \(\left( \Pi _{x,t}^v\right) _{t\ge 0}\) converges weakly to \(\pi _v\) because the diffusion is nondegenerate. This convergence and the uniform integrability (A.2) imply that
If X(t) has no invariant probability measures on \(\mathbb {R}_{++}\), then the Dirac measure with mass at 0 is the only invariant probability measure of X(t) on \(\mathbb {R}_+\). Moreover, any weak-limit of \(\left( \Pi _{x,t}^v\right) _{t\ge 0}\) as \(t\rightarrow \infty \) is an invariant probability measure of X(t) (Ethier and Kurtz 2009, Theorem 9.9 or Evans et al. 2015, Proposition 8.4). Thus, \(\left( \Pi _{x,t}^v\right) _{t\ge 0}\) converges weakly to the Dirac measure \(\delta _0\) as \(t\rightarrow \infty \). Because of (A.2) and \(\Phi (0)=0\), we have
Thus, we always have
Define
It will be shown later that \(\rho ^*>0\) whenever the population without harvesting persists, i.e. when \(\mu (0)-\sigma ^2/2>0\).
Theorem A.1
Suppose \(\mu (0)-\sigma ^2/2>0\), \(\mu (\cdot )\) satisfies Assumption 2.1 and \(\Phi (\cdot )\) satisfies Assumption 2.2. There exists a stationary Markov strategy \(v^*\in \mathfrak U_{sm}\) such that \(\pi _{v^*}\) exists and \(\rho _{v^*}=\rho ^*\). Moreover, for any admissible control h(t), we have
Proof
By (A.2) and since \(\Phi \) has a subpolynomial growth rate we can conclude that
Moreover, since \(\mu (0)-\sigma ^2/2>0\) we note that, since our population does not go extinct, \(\rho ^*>0\). On the other hand, since \(\Phi \) is continuous and \(\Phi (0)=0\) we get that \(\Phi (x)<\rho ^*\) for x is sufficiently small. This fact combined with (A.5) implies the existence of an optimal Markov strategy \(v^*\) according to Arapostathis et al. (2012, Theorem 3.4.5, Theorem 3.4.7). \(\square \)
Theorem A.2
Suppose \(\mu (0)-\sigma ^2/2>0\), \(\mu (\cdot )\) satisfies Assumption 2.1 and \(\Phi (\cdot )\) satisfies Assumption 2.2. The HJB equation
admits a classical solution \(V^*\in C^2(\mathbb {R}_+)\) satisfying \(V^*(1)=0\) and \(\rho =\rho ^*> 0\). The solution \(V^*\) of (A.6) has the following properties:
-
a)
For any \(p\in (0,1)\)
$$\begin{aligned} \lim _{x\rightarrow \infty }\dfrac{V^*(x)}{x^{p}}=0. \end{aligned}$$(A.7) -
b)
The function \(V^*\) is increasing, that is
$$\begin{aligned} V^*_x\ge 0,\, x\in \mathbb {R}_{++}. \end{aligned}$$(A.8)
A Markov control v is optimal if and only if it satisfies
almost everywhere in \(\mathbb {R}_+\).
Proof
Consider the optimal problem with the yield function
for some fixed \(x\in \mathbb {R}_{++}\) and \(h\in \mathfrak {U}\). Note that this is the \(\alpha \)-discounted optimal problem. Pick any \(0<x_1<x_2<\infty \) and let \(X^{x_1}\), \(X^{x_2}\) be the solutions to the controlled diffusion
with initial values \(x_1\), \(x_2\) respectively. Note that we are using a fixed admissible control h(t) which is the same for any initial value. The control h(t) here is not a Markov control which in general depends on the initial value. Since \(\mu (\cdot )\) is continuous and decreasing, for \(y_1, y_2>0\), there exists \(\xi (y_1, y_2)>0\) depending continuously on \(y_1, y_2\) such that \(\mu (y_1)-\mu (y_2)=-\xi (y_1, y_2)(\ln y_1-\ln y_2)\). Using Itô’s Lemma we have
which in turn yields
Therefore, if \(x_2>x_1\), we get that
This implies that \(J_h(\cdot )\) is an increasing function. Therefore, the optimal yield
is also increasing. By Arapostathis et al. (2012, Lemma 3.7.8), there is a function \(V^*\in C^2(\mathbb {R}_{++})\) satisfying (A.6) for a number \(\rho \) such that
Moreover,
for some sequence \((\alpha _n)_{n\in \mathbb {N}}\) that satisfies \(\alpha _n\rightarrow 0\) as \(n\rightarrow \infty \). This implies that \(V^*\) is an increasing function, i.e.
For any continuous function \(\psi :\mathbb {R}_{++}\mapsto \mathbb {R}\) satisfying
we have from (A.1) and Arapostathis et al. (2012, Lemma 3.7.2) that \({\mathbb {E}}^v_x |\psi (X(t))|\) exists and satisfies
and
where \(\xi _R=\inf \{t\ge 0: X(t)> R \text { or } X(t)<R^{-1}\}.\) Moreover, by using Arapostathis et al. (2012, Lemma 3.7.2) again we get that
where
and \(\tau _R:=\inf \{t\ge 0: X(t)\le R\}.\)
By Arapostathis et al. (2012, Formula 3.7.48), we have the estimate
which implies
Now, pick any \(\varepsilon >0\) and divide (A.16) on both sides by \(x^{p+\varepsilon }\). We get
and by letting \(x\rightarrow \infty \)
This implies, since p and \(\varepsilon >0\) are arbitrary, Eq. (A.7). Let \(\chi :\mathbb {R}_{++}\mapsto [0,1]\) be a continuous function satisfying \(\chi (x)=0\) if \(x<\frac{1}{2}\) and \(\chi (x)=1\) if \(x\ge 1\). Then \(\psi (x):=V^*(x)\chi (x)\) satisfies (A.12) because of (A.16). On the other hand, since \(V^*(x)\) is increasing and \(V^*(1)=0\), then \(V^*(x)\le 0\) when \(x\le 1\). Thus, we have
Let \(v^*\) be the measurable function satisfying (A.9).
By Dynkin’s formula
Letting \(R\rightarrow \infty \), we obtain from the monotone convergence theorem and (A.14) that
Letting \(t\rightarrow \infty \) and using (A.13) and (A.3), we have
This and (A.17) implies that \(\rho =\rho ^*=\rho _{v^*}\).
By the arguments from Arapostathis et al. (2012, Theorem 3.7.12), we can show that v is an optimal control if and only if (A.9) is satisfied. \(\square \)
When \(\Phi \) is the identity mapping the Eq. (A.9) becomes
which implies
Our main result is the following theorem.
Theorem 2.1
Assume that \(\Phi (x)=x, x\in (0,\infty )\) and that the population survives in the absence of harvesting, that is \(\mu (0)-\frac{\sigma ^2}{2}>0\). Furthermore assume that the drift function \(\mu (\cdot )\) satisfies Assumption 2.1. The optimal control (the optimal harvesting strategy) v has the bang–bang form
for a unique \(x^*\in (0,\infty )\). Furthermore, we have the following upper bound for the optimal asymptotic yield
Remark A.1
If \(V_x^*(x)=1\) then we note that (A.18) does not provide any information about v(x). However, in this case we can set the harvesting rate equal to anything since the yield function will not change. This is because our diffusion is non-degenerate and changing the values of the drift on a set of zero Lebesgue measure does not change the distribution of X.
We split up the proof of Theorem 2.1 into a few propositions. It is immediate to see that the HJB equation (A.6) becomes
Sketch of proof of Theorem 2.1
Since the optimal control is given by (A.18) we need to analyze the properties of the function \(V_x^*\) which by (A.19) satisfies a first order ODE. The analysis of this is split up into several propositions. Note that the ODE governing \(V_x^*\) is different, depending on whether \(V_x^*>1\) or \(V_x^*\le 1\).
In Proposition A.1 we analyze the ODE for when \(V_x^*\le 1\) and find its asymptotic behavior close to 0. Using this we can show in Proposition A.2 that one cannot have a \(\eta >0\) such that \(V_x^*(x)\le 0\) for all \(x\in (0,\eta ]\).
Similarly, in Proposition A.3 we show that there can exist no \(\zeta >0\) such that \(V_x^*(x)\ge 1\) for all \(x\ge \zeta \).
In Proposition A.4 we explore the possible ways \(V_x^*\) can cross the line \(y=1\) and find using soft arguments that there can be at most 3 crossings. Finally, we show that actually there must be exactly one crossing of \(y=1\) by \(V_x^*\) and that this crossing has to be from above. This combined with (A.18) completes the proof. \(\square \)
Proposition A.1
Assume \(\mu (\cdot )\) is locally Lipschitz on \([0,\infty )\). Then any solution \(\varphi _2\) of the ODE
satisfies
Proof
It follows from the method of integrating factors that the solution to the ODE (A.20) is
where the non-homogeneous term is \(\beta (y):=\frac{2(\rho -My)}{\sigma ^2 y^2}\), and the integrating factor is
for \(\gamma (y):=\frac{2(\mu (y) -M)}{\sigma ^2 y}\), and arbitrary \(x_0, x_1\in (0,\infty )\). Since \(\mu \) is locally Lipschitz at \(x=0\), there are constants \(L, K>0\) such that for any \(x\in [0,L]\), \(|\mu (x)-\mu _0|\le K x\), where \(\mu _0:=\mu (0)\). From now on, we choose \(x_1:=L\) (or any number between 0 and L). We have, for any \(x\in [0,x_1]\),
This implies that as \(x\rightarrow 0^+\),
On the other hand, from now on, if we choose \(x_0>0\) sufficiently close to 0 such that \(\rho -Mx>0\) and (A.23) holds for all \(x\in (0,x_0)\), then we have, for any \(0<x<x_0\),
where the constants \(C_i\) are given by
Now, using the asymptotic properties (A.23) and (A.24), we can analyze the limit of \(\varphi _2\) as follows.
Case 1:\(\mu _0 < M\).
In this case, we get from (A.23) and (A.24) that
Thus, we can apply l’Hôpital’s rule and obtain
since \(\rho >0\). This shows the limit (B.4).
Case 2:\(M\le \mu _0 \le M + \frac{\sigma ^2}{2}\).
For this range of \(\mu _0\), it follows from (A.23) and (A.24) again that
but \(\lim _{x\rightarrow 0^+} \zeta (x)\) exists and is finite. Hence, we can obtain the limit (B.4) by passing to the limit \(x\rightarrow 0^+\) in the solution formula (A.22).
Case 3:\(\mu _0 > M + \frac{\sigma ^2}{2}\).
In this final case, it follows from (A.23) and (A.24) that \(\lim _{x\rightarrow 0^+} \zeta (x) = 0\) and
exists and is finite. If \(J\ne 0\), then passing to the limit \(x\rightarrow \infty \) in the solution formula (A.22) will imply the limit (B.4). Otherwise, we can apply l’Hôpital’s rule and do the same computations we did in (A.25). This proves the limit (B.4).
Putting together Cases 1,2 and 3 completes the proof. \(\square \)
Proposition A.2
There does not exist any \(\eta >0\) such that \(V_x^*(x)\le 1, x\in (0,\eta ]\).
Proof
We will argue by contradiction. Assume there exists \(\eta >0\) such that \(V_x^*(x)\le 1, x\in (0,\eta ]\). Then by (A.19) we get that \(V_x^*\) follows the ODE (A.20) for all \(x\in (0,\eta )\). Making use of Proposition A.1 we get that
which contradicts that \(V_x^*\ge 0\) or that \(V_x^*(x)\le 1, x\in (0,\eta ]\). The proof is complete. \(\square \)
The above Proposition shows that the scenario from Fig. 5 cannot happen.

If \(V^*_x\) crosses \(y=1\) from below at \(x_0\), and it has not crossed from above before then we get a contradiction by Proposition A.2
Proposition A.3
There does not exist any \(\chi >0\) such that \(V_x^*(x)\ge 1\) for all \(x\ge \chi \).
Proof
Once again we will argue by contradiction. Assume there exists \(\chi >0\) such that \(V_x^*(x)\ge 1\) for all \(x\ge \chi \). By (A.19) \(V_x^*\) will follow the ODE
for all \(x\ge \chi \). As a result we get just as in Proposition A.1
where the non-homogeneous term is \(\beta (y):=\frac{2\rho }{\sigma ^2 y^2}\), and the integrating factor is
for \(\gamma (y):=\frac{2\mu (y)}{\sigma ^2 y}\), and arbitrary \(x_0, x_1\in (\chi ,\infty )\). Under Assumption 2.1 we can see that there exist constants \(L>0\) and \(c>0\) such that \(\mu (y) < -c\) for all \(y>L\), and hence, \(\int _{L}^x \frac{\mu (y)}{y} \;dy \le -c \int _{x_1}^x \frac{1}{y} \;dy = -c (\ln x - \ln x_1) \rightarrow -\infty \) as \(x\rightarrow \infty \). If we choose \(c> \frac{\sigma ^2}{2}\), \(x_1:=L\) we get
as \(x\rightarrow \infty \). If
then by (A.27) and the positivity of \(\zeta \) one has
which contradicts the growth condition (A.7). Therefore we need
Note that in this case
This implies, since \(\zeta (x)>0\), that for \(x>x_0\)
which contradicts the assumption that \(V_x^*(x)\ge 1\) for all \(x\ge \chi \). \(\square \)
The above Proposition shows that the scenario from Fig. 6 is not possible.

An impossible scenario, by Proposition A.3
Set \(g(x):=\rho - x\mu (x)\). By assumption \(p(x):=x\mu (x)\) has a unique maximum and \(\mu \) is locally Lipschitz and decreasing with \(\lim _{x\rightarrow \infty } \mu (x)=-\infty \). This implies that g(x) has a unique minimum for some \(x_\iota \in (0,\infty )\).Footnote 4 If \(g(x_\iota )<0\) then g intersects the x axis in exactly two points \(0<\alpha _1<\alpha _2<\infty \). If \(g(x_\iota )>0\) there is no intersection of g with the x axis. Finally, if \(g(x_\iota )=0\) there is exactly one intersection and this happens at \(x=x_\iota \).
Proposition A.4
The function \(V^*_x\) crosses the line \(y=1\) at most three times. More specifically, we have the following possibilities:
-
(I)
If \(g(x_\iota )<0\) then
-
(i)
For \(0\le x<\alpha _1\) the function \(V_x^*\) can only pass the line \(y=1\) at most once and the crossing has to be from below.
-
(ii)
For \(x>\alpha _2\) the function \(V_x^*\) can pass the line \(y=1\) at most once and the crossing has to be from below.
-
(iii)
For \(\alpha _1<x<\alpha _2\) the function \(V_x^*\) can pass the line \(y=1\) at most once and the crossing has to be from above.
-
(i)
-
(II)
If \(g(x_\iota )>0\) then the function \(V_x^*\) can pass the line \(y=1\) at most once and the crossing has to be from below.
-
(III)
If \(g(x_\iota )=0\) then \(V_x^*\) can cross the line \(y=1\) at most three times. In particular, the possible crossing(s) in \((0,x_\iota ) \cup (x_\iota ,\infty )\) must be from below.
-
(IV)
If \(V_x^*\) crosses the line \(y=1\) at \(x_0\) then we cannot have \(\varepsilon >0\) such that \(V_x^*=1\) on \((x_0,x_0+\varepsilon )\). In other words, the intersections have to be at separate points and we cannot ‘stick’ to \(y=1\).
Proof
It follows from the HJB Eq. (A.6) with \(\varphi :=V_x\) that if \(\varphi (x_0)=1\), then we have
Therefore, when \(\varphi \) crosses the line \(y=1\), we obtain some information from g. More precisely, we can infer the following:
-
(I)
When \(g(x_\iota )<0\) the function \(g(x) = \rho - x\mu (x)\) has exactly two zeros at \(\alpha _1, \alpha _2\) with \(0<\alpha _1<\alpha _2<\infty \).
-
(ii)
for \(0\le x<\alpha _1\) we have \(g(x) > 0\), hence \(\varphi \) is only allowed to cross the line \(y=1\) from below in this region;
-
(iii)
for \(x>\alpha _2\) we have \(g(x) > 0\), hence \(\varphi \) is only allowed to cross the line \(y=1\) from below in this region;
-
(iv)
for \(\alpha _1<x<\alpha _2\), \(g(x)<0\) and \(\varphi \) is only allowed to cross the line \(y=1\) from above in this region.
-
(ii)
-
(II)
If \(g(x_\iota )>0\) then \(g(x)> 0\) for all \(x\in \mathbb {R}_+\). The function \(V_x^*\) can pass the line \(y=1\) at most once and the crossing has to be from below.
-
(III)
If \(g(x_\iota )=0\) then g(x) has a unique intersection of the x axis at \(x_\iota \). As a consequence \(g(x)\ge 0\) and the function \(V_x^*\) can pass the line \(y=1\) at most thrice: at most once from below in the region \(x<x_\iota \), at most once from below in the region \(x>x_\iota \) and at most once from above or from below at the point \(x= x_\iota \).
-
(IV)
Since \(x\mu (x)\) is never constant on an interval, it is clear that for any \((u,v)\subset \mathbb {R}_+\) we cannot have \(V_x^*=1\) for all \(x\in (u,v)\).
\(\square \)
Remark A.2
By the analysis above one can note that at the intersection points (or roots) \(\alpha _{1,2}\) of the function g(x) with the x axis the derivative of \(\varphi \) is 0. This makes it more complicated to say, in case there is a crossing at a root, if the crossing is from above or from below. However, this does not require us to change our arguments. For example, if there is a crossing from below on \(0\le x<\alpha _1\) and there is a crossing at \(x=\alpha _1\) then the crossing at \(\alpha _1\) is necessarily from above. This then implies that there can be no crossing for \(x\in (\alpha _1,\alpha _2)\) because in this region the crossing has to be from above and there cannot be two crossings from above in a row.
Proof of Theorem 2.1
A direct consequence of Proposition A.4 is that \(V_x^*\) can cross the line \(y=1\) at most three times. We also know, given the at most two possible solutions \(\alpha _{1,2}\) of the equation \(g(x)=0\) how these crossings have to happen. Next, we eliminate all but one possibility.
-
i)
If we get a crossing from below in \((0,\alpha _1)\) this means that there exists \(\eta >0\) such that for all \(x\in (0,\eta )\) we have \(V_x^*(x)=\varphi _2(x) \le 1\). This is not possible by Proposition A.2. As such there can be no crossings in \((0,\alpha _1)\).
-
ii)
If we have a crossing from below in \((\alpha _2,\infty )\) then there is \(\zeta >0\) such that for all \(x\ge \zeta \)
$$\begin{aligned} V_x^*(x)=\varphi _1(x)\ge 1. \end{aligned}$$This is not possible by Proposition A.3. Therefore, there are no crossings in \( (\alpha _2,\infty )\).
-
iii)
We cannot have that \(V_x^*(x)\ge 1\) for all \(x\in (0,\infty )\) because then we get a contradiction by Proposition A.3. Similarly, we cannot have \( V_x^*(x)\le 1\) for all \(x\in (0,\infty )\) since we get a contradiction by Proposition A.2.
-
iv)
If \(g(x_\iota )>0\) then, in principle, there could be at most one crossing and this would have to be from below. But this creates a contradiction by either using Proposition A.2 or Proposition A.3. If there is no crossing then we get a contradiction by (iii) above.
-
v)
If \(g(x_\iota )=0\) then
-
(a)
If there is no crossing, then we get a contradiction by part iii) above.
-
(b)
If there are two crossings then we get contradictions from either Proposition A.2 or Proposition A.3.
-
(c)
If there are three crossings then we must have a crossing from below in \((0,x_\iota )\), one from above at \(x=x_\iota \) and one from below in \((x_\iota ,\infty )\). This yields a contradiction because of Proposition A.2.
-
(d)
If there is just one crossing and the crossing is from below then we get a contradiction by Proposition A.3.
-
(a)
-
vi)
By parts i)-iv) we get that there is exactly one crossing of the line \(y=1\), that this crossing is from above and that the crossing happens at a point in the interval \([\alpha _1,\alpha _2]\) when \(g(x_\iota )<0\) or at \(x_\iota \) if \(g(x_\iota )=0\).

The only case which doesn’t lead to a contradiction is when \(V^*_x\) crosses \(y=1\) only once and the crossing is from above
This, together with (A.18), implies that the optimal strategy is of bang–bang type
Moreover, one can see that \(g(x_\iota )\le 0\) which in turn forces
\(\square \)
Appendix B: Optimal harvesting with concave and convex yields: proofs
This appendix shows that for a class of yield functions \(\Phi \) one can get continuous optimal harvesting strategies. Therefore, the optimal harvesting strategy will be discontinuous. One might wonder under which conditions on \(\Phi \) the optimal harvesting strategies will be continuous (Fig. 7).
We proved in Theorem A.2 that the HJB equation
admits a classical solution \(V^*\in C^2(\mathbb {R}_+)\) satisfying \(V^*(1)=0\) and \(\rho =\rho ^*> 0\).
For any given \(\Phi \), we define
where A is a shorthand of \(V_x^*\), that is,
For any fixed x, we can see A as a constant. Using these shorthands, we can rewrite the HJB equation as
where \(L:=xM\). A direct computation yields
because \(\Phi (0) = 0\). Therefore, the critical point(s) will be given by \(\omega _c = [\Phi ']^{-1}(A)\), and
If \(\Phi \) is assumed to be strictly concave, the maximum on the right hand side of (B.1) can be found easily because \(F''=\Phi ''\).
Theorem 3.1
Suppose Assumption 2.1 holds and the yield function satisfies
-
(1)
\(\Phi \in C^2(\mathbb {R}_+)\),
-
(2)
\(\Phi \) is strictly concave.
Then the optimal harvesting strategy is continuous and given by
Furthermore, the HJB equation for the system becomes
Proof
Assume that \(\Phi \) is \(C^2\) and strictly concave. Since \(\Phi \) is \(C^2\) we have that \(\Phi ''<0\). In this case, \(\Phi '\) is strictly decreasing, so its inverse is well-defined. As a result, we have a unique critical point which is a maximum \(\omega _c = [\Phi ']^{-1}(A)\). A standard calculus result yields
where we used the fact that \(F(0)=0\) and the concavity of \(\Phi \) in the last equality.
Depending on the maximum point, we have the corresponding optimal Markov control:
because v is the solution to
In conclusion, in this case, v depends on \(A:=\dfrac{d V^*}{d x}(x)\) continuously. Hence, since \(V^*\in C^2\left( \mathbb {R}_+\right) \) we conclude that v is continuous.
The HJB Eq. (A.6) becomes
\(\square \)
The case when the yield function \(\Phi \) is convex is qualitatively similar to the case when the yield function is linear, and the optimal solution is of the bang–bang type. We can improve Theorem 2.1 as follows.
Theorem 3.2
Assume that \(\Phi :\mathbb {R}_+\rightarrow \mathbb {R}_+\) is weakly convex, \(\Phi \) grows at most polynomially, \(\Phi \in C^1(\mathbb {R_+})\) and the population survives in the absence of harvesting, that is \(\mu (0)-\frac{\sigma ^2}{2}>0\). Furthermore assume that the drift function \(\mu (\cdot )\) satisfies the following modification of Assumption 2.1:
-
(i)
\(\mu \) is locally Lipschitz.
-
(ii)
\(\mu \) is decreasing.
-
(iii)
As \(x\rightarrow \infty \) we have \(\mu (x)\rightarrow -\infty \).
-
(iv)
The function
$$\begin{aligned} G(x)= \Phi (xM)\left( 1-\frac{2}{\sigma ^2}\mu (x)\right) - xM\Phi '(xM) \end{aligned}$$(3.2)has a unique extreme point in \((0,\infty )\) which is a minimum, and is not constant on any interval \((u,v)\subset \mathbb {R}_+\).
If the assumptions (i)–(iii) hold, the optimal control has a bang–bang form (i.e., the harvesting rate is either 0 or the maximal M). If assumptions (i)–(iv) hold, the optimal harvesting strategy v has a bang–bang form with one threshold
for some \(x^*\in (0,\infty )\).
Proof
This proof is similar to the proof of Theorem 2.1 from Appendix A. By (A.9)
Dropping the common terms gives
With \(x>0\), the right hand side is a weakly convex function of u, so one of the end points of the interval U achieves the maximum. This already shows that the optimal control is bang–bang, but says nothing else of the shape of v(x). Since \(\Phi (0) = 0\), we get
This implies
The function \(\Phi (\cdot )\) is weakly convex, therefore, for \(\alpha \in (0,1)\), \(\Phi (\alpha x + (1-\alpha ) y ) \le \alpha \Phi (x) + (1-\alpha )\Phi (y)\). By assumption, it is also continuous and positive valued. So, for \(\alpha \in (0,1)\), \(\alpha \Phi (xM) \ge \Phi (\alpha xM)\), equivalent with \(\Phi (xM) \ge \frac{1}{\alpha } \Phi (\alpha xM)\), equivalent with \(\frac{\Phi (xM)}{xM} \ge \frac{\Phi (\alpha xM)}{\alpha xM}\) if \(x,M>0\). Therefore \(\frac{\Phi (xM)}{xM}\) must be positive and monotonically increasing in x for \(M>0\), \(x>0\). In particular \(\Phi '(0) = \lim _{x\rightarrow 0^+} \frac{\Phi (xM)}{xM}\) exists and it is greater or equal to 0. The HJB equation A.6 becomes
One can easily modify the proofs from Appendix A to show the following four propositions:
Proposition A.5
Assume \(\mu , \Phi \) satisfy the assumptions of Theorem 3.2. Then any solution \(\varphi _2\) of the ODE
satisfies
Proof
Proceed similarly to the proof of Proposition A.1, replacing the definition \(\beta (y): = \frac{2(\rho - \Phi (My))}{\sigma ^2 y^2}\). This time,
For \(y \in [0,x_0]\), we have \(\Phi '(0) \le \frac{\Phi (My)}{My} \le \frac{\Phi (Mx_0)}{Mx_0}\), so
For a general positive constant N,
where the integration constants are given by
Now the case-by-case analysis of Proposition A.1 can be repeated similarly because the constants of the dominant terms in the expression above do not depend on N. \(\square \)
Proposition B.2
There does not exist any \(\eta >0\) such that \(V_x^*(x)\le \frac{\Phi (xM)}{xM}, x\in (0,\eta ]\).
Proof
Noting that \(\sup _{x\in (0,\eta ]} \frac{\Phi (xM)}{xM} = \frac{\Phi (\eta M)}{\eta M}\), the proof is similar to the proof of Proposition A.2, relying on the application of Proposition A.5 to Eq. (B.3). \(\square \)
Proposition B.3
There does not exist any \(\chi >0\) such that \(V_x^*(x)\ge \frac{\Phi (xM)}{xM}\) for all \(x\ge \chi \).
Proof
It follows the proof of Proposition A.3 without change, because \(\frac{\Phi (xM)}{xM} \ge 0\). \(\square \)

The only case which doesn’t lead to a contradiction is when \(V^*_x\) crosses \(\frac{\Phi (xM)}{xM}\) only once and the crossing is from above
Proposition B.4
The function \(V^*_x\) intersects the curve \(\frac{\Phi (xM)}{xM}\) at most three times on \([0,\infty )\).
Proof
By (B.2) if we set \(f_x:=\varphi \), then at the intersections \(x:\; \varphi (x) = \frac{\Phi (xM)}{xM}\) we have
from the HJB equation. Now we want to compare \(\varphi _x\) with \(\left( \frac{\Phi (xM)}{xM}\right) '\) whenever there is a crossing, to infer the direction from which \(\varphi \) is crossing. To do that, consider the equation \(\varphi _x = \left( \frac{\Phi (xM)}{xM}\right) '\). Substituting and simplifying gives us the condition \(G(x) + \frac{2M\rho }{\sigma ^2} = 0\) where G(x) is defined in 3.2. Since G(x) has only one extremum by assumption, this equation has zero, one or two solutions. When there are two solutions, say \(\alpha _1, \alpha _2\), any intersection of \(\varphi \) with \(\frac{\Phi (xM)}{xM}\) for \(x \in (\alpha _1,\alpha _2)\) will have to be with \(\varphi \) coming from above, as \(\varphi _x <0\) in that interval. Using similar arguments to those in Proposition A.4, this implies, together with the condition on G from (3.2), that \(\varphi \) can intersect \(\frac{\Phi (xM)}{xM}\) at most three times. \(\square \)
The rest of the proof also mirrors the one of Theorem 2.1. Apply the four results above and find again that the optimal control is bang–bang with a single threshold \(x^*\),
for some \(x^*\in (0,\infty )\) (see Fig. 8). \(\square \)
Rights and permissions
About this article

Cite this article
Hening, A., Nguyen, D.H., Ungureanu, S.C. et al. Asymptotic harvesting of populations in random environments. J. Math. Biol. 78, 293–329 (2019). https://doi.org/10.1007/s00285-018-1275-1
Received:
Revised:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00285-018-1275-1