
Abstract
The occurrence and distribution of an actinobacteria group of bacteria capable of dissolving insoluble phosphates were investigated in this study in marine environments, especially in sediments of Chorao Island, Goa Province, India. A total of 200 bacterial isolates of actinobacteria was isolated. All isolates were screened for phosphate-solubilizing activity on Pikovskaya’s agar. Thirteen different isolates exhibiting maximum formation of halos (zone of solubilization) around the bacterial colonies were selected for quantitative estimations of P-solubilization. Quantitative estimations for P-solubilization were analyzed for up to 10 days at intervals of 24 h. Maximum solubilization from 89.3 ± 3.1 to 164.1 ± 4.1 μg ml−1 was observed after 6 days of incubation in six of all isolates, while the isolate NII-1020 showed maximum P-solubilization. The increase in solubilization coincided with the drop in pH. Many of these species showed wide range of tolerance to temperature, pH, and salt concentrations. Further, 16S rRNA gene sequence analyses were carried to identify the bacterial groups which are actively solubilized phosphate in vitro. Gene sequencing results reveal that all isolates were clustered into six different actinobacterial genera: Streptomyces, Microbacterium, Angustibacter, Kocuria, Isoptericola, and Agromyces. The presence of phosphate-solubilizing microorganisms and their ability to solubilize phosphate were indicative of the important role played by bacteria in the biogeochemical cycle of phosphorus and the plant growth in coastal ecosystems.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Phosphorus is a limiting nutrient, in spite of being a major component of seawater. Major forms are either bound to or strongly adsorbed to silt and clay. Phosphorus normally occurs as phosphates in both inorganic and organic compounds [3, 14, 26]. Moreover, phosphate uptake has been found to be dominated by bacteria [5]. Microorganisms assimilate inorganic phosphate and mineralize organic phosphorus compounds, and microbial activities were involved in the solubilization and mobilization of phosphates. Orthophosphate appears to be the preferred and apparently universally used substrate. Mainly, phosphorus is solubilized enzymatically or by acid hydrolysis. Though phosphate-solubilizing bacteria (PSB) like Pseudomonas, Serratia, Bacillus, Flavobacterium, and Corynebacterium have been reported from various ecosystems, such as terrestrial especially from soil and water ecosystem including coastal, offshore, and mangrove [4, 23], a series of observations indicates that the highest population of PSB (an average of 108 CFU per gram of sample) was found in fertile soils of forests, organic farming, and rhizosphere [30–34], as well as in mangrove and shorelines samples [8]. PSB form sea sediments were reported to be capable of accelerating the dissolution of apatite phosphate within the phosphorus cycle and interacting with the carbon cycle [24]. PSB living in both terrestrial and water ecosystems play a vital role in supplying phosphorus to plants [10]. The present study aims at studying the occurrence and identification of phosphate-solubilizing actinobacteria in the sediments of Chorao Island, Goa, India.
Materials and Methods
Sampling and Collection of Sediment Samples
Chorao Island
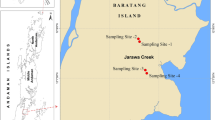
The Chorao Island is located between the Mandovi and the Mapusa Rivers in Goa, India. The western side of the island is of a peninsular shape and occupied by a thick mangrove forest of about 1.78 km2. The geographical location of the station is 15°32′34″ latitude and 73°55′15″ longitude (Fig. 1). Samples were collected from the middle of the section of the river and from the surface layer of the sediments with a Peterson grab. The central portions of the samples were aseptically transferred to sterile glass bottle and transferred to the laboratory for further analysis.

Sampling site location and phosphate-solubilizing ability of isolates on Pikovskaya’s media
Phosphate-Solubilizing Bacteria
Solubilization of precipitated tricalcium phosphate-TCP (Ca3(PO4)2) in unbuffered solid agar medium plates has been used widely as the initial criterion for the isolation of phosphate-solubilizing microorganisms. The sediment samples were serially diluted, spread plated on Pikovskaya’s agar (g l−1) (glucose—10 g, Ca3(PO4)2—5 g, (NH4)2SO4—0.5 g, KCl—0.2 g, MgSO4—0.1 g, MnSO4—traces, FeSO4—traces, yeast extract—0.5 g, and agar—15 g, pH 7.0), and incubated at 30 ± 2 °C for 1–2 weeks. A total of 200 bacteria isolates with P-solubilization halo zone on plates was purified with repeated culturing and maintained in 20 % glycerol at −80 °C. All the subsequent experiments were conducted after raising fresh culture. Potential isolates which are showing more halo zones of P-solubilization (>9 mm) were assessed for further characterization.
Characterization, Identification, and Phylogenetic Analysis
Preliminary biochemical characterization was carried out as per standard methodologies [6]. Growth versus incubation period was checked for a maximum duration of 240 h, and temperature range for growth was checked on nutrient broth from 10, 25, 37, 45, and 55 °C by inoculating 1 ml of an exponential culture (108 CFU ml−1) in 100 ml of nutrient broth and estimating the cell population at 12-h intervals. The effect of salt concentration of the growth organisms were performed from 0, 3, 5, 7, 9, and 12 % (w/v) for up to 240 h at 30 °C. The identity of the isolate was revealed on the basis of Biolog carbon source utilization; for evaluating carbon source utilization, bacterial suspensions were inoculated into Biolog GN2 Microplates as described in instruction manuals and incubated at 30 °C for 24 h. The results were interpreted using Biolog Micro Log 34.20.04 software (Biolog, Hayward, CA), and further validated by performing the 16S rRNA gene sequencing using an ABI PRISM BigDye Terminator cycle sequencing kit (as recommended by the manufacturer). The universal primers 27F and 1492R were used for the partial sequencing of the 16S rRNA gene. Sequences of the isolates were compared with the sequence data available by the BLAST search in the NCBI, GenBank database (http://www.ncbi.nlm.nih.nov) [1]. Multiple sequence alignments were performed using CLUSTALW [25]. The method of Jukes and Cantor [13] was used to calculate evolutionary distances. Phylogenetic and molecular evolutionary analyses were conducted using mega version 4.0 [16]. The phylogenetic trees were constructed by the neighbor-joining method [22] using the distance matrix from the alignment. Distances were calculated using the Kimura method [15]. All the sequences were submitted to NCBI GenBank.
Phosphate Solubilization
After qualitative estimation of the P-solubilizing activity which was carried out on Pikovskaya’s agar, quantitative estimation of P-solubilization was carried out as per standard methodology [17], by inoculating 1 ml of bacterial suspension (3 × 107 cells ml−1) in 50 ml of National Botanical Research Institute Phosphate broth (NBRIP) in Erlenmeyer flasks (150 ml), which contained the following ingredients (g l−1): glucose, 10.0; tricalcium phosphate (TCP), 10.0; MgCl2·6H2O, 5.0; MgSO4·7H2O, 0.25; KCl, 0.2; and (NH4)2SO4, 0.1 and incubating the flasks for 10 days at 30 °C. At the end of the incubation period, the cell suspension was centrifuged at 10,000 rpm for 10 min, and the phosphate content in the supernatant was spectrophotometrically estimated by the ascorbic acid method [18]. All the studies were repeated on three independent experiments to confirm the results. Sterile water-inoculated media was treated as blank. After incubation for 10 days, pH of the medium was recorded on a pH meter equipped with glass electrode.
Results
Isolation of Phosphate-Solubilizing Bacteria (PSB)
A total of 200 bacterial isolates of actinobacteria was isolated. All isolates were screened for phosphate-solubilizing activity on Pikovskaya’s agar. Thirteen different isolates exhibiting maximum formation of halos (zone of solubilization) in 9–23 mm zone of phosphate solubilization after 6 days of incubation around the bacterial colonies were selected for quantitative estimations of P-solubilization (Fig. 1). Phenotypic and biochemical characterization were performed using Biolog-based carbon source utilization and Hi25 Kit (HiMedia, Mumbai) presented in Table 1. Molecular analysis based on 16S rRNA gene sequencing results reveals all isolates grouped together into six different genera of actinobacteria, some of which are being reported for the first time as P-solubilizers. The isolates were shown optimum growth incubation of 144 h, 3–7 % (w/v) tolerance to salinity and temperature of 37 °C (Table 1), which can be used for the preparation of biofertilizer from local isolates based on geographical conditions.
Phosphate-Solubilization Activity
The details of the pH and amounts of soluble-P in the medium from 12 to 240 h of incubation are presented in Table 2. The solubilization of Ca3(PO4)2 in the liquid medium by different strains was accompanied by a significant drop in pH (3.0–4.5) from an initial pH of 7.0 ± 0.2 after 144 h was recorded. The soluble-P concentration in the medium ranged between 89.3 ± 3.1 and 164.1 ± 4.1 μg ml−1 with variations among different isolates. In the blank treatment without addition of any bacterial isolate, no soluble-P was detected or any drop in pH was observed. The maximum P-solubilization was recorded by Kocuria sp. NII-1020 and NII-1014, followed by Microbacterium sp. NII-1011, NII-1021, and NII-1012; Isoptericola sp. NII-1016 and 1029; Streptomyces sp. NII-1008 and NII-1005; and Agromyces sp. NII-1018, and very low P-solubilization levels were observed in Angustibacter sp. NII-1013 and in two Streptomyces sp. NII-1007 and NII-1002. Maximum drop in pH was associated with higher levels of P-solubilization, Kocuria sp. (NII-1020), where pH was decreased to 4.0 from initial pH 7.0; comparatively higher amounts of soluble-P (164.1 ± 4.1 μg ml−1) were detected in the medium. The maximum P-solubilization was detected after 144 h of incubation. The decrease in pH clearly indicates the production of organic acid and phosphatase, which is considered to be responsible for the phosphate-solubilization activity (Table 2).
Phylogenetic Analysis
The identification of phosphate-solubilizing bacterial strains based on 16S rRNA gene sequence and their phylogeny sequences are presented in Supplementary Fig. 1. All the 13 isolates were grouped together into six genera: four strains were grouped to Streptomyces, three isolates to Microbacterium, two isolates to Kocuria, two isolates grouped to Isoptericola sp., and one each to Agromyces and Angustibacter; some species showed less sequence similarities with the closest known species in the NCBI GenBank.
Nucleotide Accession Numbers
The 16S rRNA sequences of phosphate-solubilizing actinobacteria were deposited in the NCBI GenBank database, and the accession numbers are presented in Table 3.
Discussion
Usually phosphates precipitate because of the abundance of cations in the interstitial water of mangrove sediments making phosphorus mostly unavailable to plants. P-solubilizing bacteria as a potential supplier of soluble forms of phosphorus have a great advantage for mangrove plants. The generally anoxic conditions of the sediments beneath the aerobic zone would tend to favor dissolution of non-soluble phosphate through sulfide production. However, depending on the degree of aeration of the rhizosphere by root oxygen translocation, the bacteria might be of value in solubilizing phosphate near the roots where sediments are not always completely anoxic, being similar to the fact that upland rice tends to require more phosphate than paddy rice [35]. Surprisingly, very limited research has been focused on P-solubilizing efficiency of actinobacteria in the marine environment, either in temperate or tropical regions [2, 7, 9, 20, 28]. In an arid mangrove ecosystem in Mexico, nine strains of P-solubilizing bacteria were isolated from black mangrove (A. germinans) roots: Bacillus amyloliquefaciens, B. atrophaeus, Paenibacillus macerans, Xanthobacter agilis, Vibrio proteolyticus, Enterobacter aerogenes, E. taylorae, E. asburiae, and Kluyvera cryocrescens, and three strains from white mangrove (Languncularia racemosa) roots: B. licheniformis, Chryseomonas luteola, and Pseudomonas stutzeri [27]. Phosphate-solubilizing actinobacteria were recorded from all the sampling stations in Chorao Island, among which 13 isolated of six different genera showed positive phosphate-solubilizing activity. Production of halo zones on solid agar and release of phosphate in the medium could be ascribed to the release of organic acids by the microbes [23]. The ability of PSB to quantitatively dissolve phosphate and the pH of the liquid media with various P sources after 6-day incubation were measured using spectrophotometer. It shows that all of the tested isolates were able to dissolve the inorganic phosphate of Ca3(PO4)2 within the liquid medium of NBRIP The highest concentration of dissolved P was found in Kocuria sp. NII-1020 and NII-1014 with the amounts of 164.1 ± 4.1 and 145.1 ± 2.7 μg ml−1, respectively (Table 2). The lowest concentration was found in Streptomyces sp. NII-1002 with an amount of 89.3 ± 3.1 μg ml−1. Jeon et al. [12] showed that Pseudomonas fluorescens dissolves Ca3(PO4)2 after 5-day incubation and pH drops to 4.4. Perez et al. [19] stated that the acidity of a liquid culture is the main mechanism of phosphate dissolution. Whitelaw et al. [29] also stated that the concentration of phosphate dissolved in the culture media is in line with the acidity and concentration of amino acid as well as pH shift. Ramachandran et al. [21] stated that since the isolate is able to release inorganic phosphate from Ca3(PO4)2 in the liquid media, the bacteria have the potential to dissolve the bonded P, thus making it available for the plants.
De Souza et al. [8] reported the percentage rates of the occurrence of PSB from different niches, viz., beaches, islands, coasts, and offshore, and it was found that phosphate-solubilizing bacterial population is higher in coastal areas than in offshore areas. It is probable that the offshore organisms are poor solubilizers of inorganic phosphate because these organisms are hampered by an insufficient supply of easily degradable organic compounds and low uptake of carbon [8, 11]. Hence, the lower percentage in the occurrence of PSB observed in the present study may be due to the location of the stations in the deeper waters. However, in terms of phosphate-solubilization efficiency, the isolates showed a good result. This is the first and the only study to report the phosphate-solubilizing capacity of actinobacteria belonging to the genera Streptomyces, Microbacterium, Angustibacter, Kocuria, Isoptericola, and Agromyces, and their presence in mangrove sediments. The P-solubilizing activities of all of these species were demonstrated first by halo formation around bacterial colonies growing on a solid medium supplemented with insoluble calcium phosphate.
Conclusion
From the above study, it is concluded that actinomycetes/actinobacteria are also efficient strains for inducing phosphate-solubilization activity. The PSB are widely distributed in different niches, with the coastal areas having a higher density. Sediment was found to be the most efficient carrier. Hence, these isolates could serve continuously to fertilize a niche by solubilizing insoluble phosphorus compounds especially in environments where a low concentration of phosphorus causes various limitations. The contributions of PSB to phosphorous transfer can be analyzed by means of these bacteria in the sediments. It is believed that these studies will potentially enhance our awareness of the roles of microorganisms in aquatic systems and the mechanisms of eutrophication.
References
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman OJ (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acid Res 25:3389–3402
Ayyakkannu K, Chandramohan D (1971) Occurrence and distribution of phosphate solubilizing bacteria and phosphatase in marine sediments at Porto Novo. Mar Biol 11:201–205
Biswas DR, Narayanasamy G (2006) Rock phosphate enriched compost: an approach to improve low-grade Indian rock phosphate. Bioresour Technol 97:2243–2251
Chen YP, Rekha PD, Arun AB, Shen FT, Lai WA, Young CC (2006) Phosphate solubilizing bacteria from subtropical soil and their tricalcium phosphate solubilizing abilities. Appl Soil Ecol 34:33–41
Chung H, Park M, Madhaiyan M, Seshadri S, Song J, Cho H et al (2005) Isolation and characterization of phosphate solubilizing bacteria from the rhizosphere of crop plants of Korea. Soil Biol Biochem 37:1970–1974
Collins CH, Lyne PM (1980) Microbiological methods. Butterworth and Co., London
Craven PA, Hayasaka SS (1982) Inorganic phosphate solubilization by rhizosphere bacteria in a Zostera marina community. Can J Microbiol 28:605–610
De Souza MJBD, Nair S, Chandramohan D (2000) Phosphate solubilizing bacteria around Indian Peninsula. Indian J Mar Sci 29:48–51
Devendran K, Sundararaj V, Chandramohan D, Krishnamurthy K (1974) Bacteria and primary production. Indian J Mar Sci 3:139–141
Gyaneshwar P, Kumar GN, Parekh LJ, Poole PS (2002) Role of soil microorganism in improving P nutrition of plants. Plant Soil 245:83–93
Hoppe HG, Ullrich S (1999) Profiles of ecto-enzymes in the Indian Ocean: phenomena of phosphatase activity in the mesopelagic zone. Aqua Microb Ecol 19:139–148
Jeon JS, Lee SS, Kim HY, Ahn TS, Song HG (2003) Plant growth promotion in soil by some inoculated microorganisms. J Microbiol 41(4):271–276
Jukes TH, Cantor CR (1969) Evolution of protein molecules. In: Munro HN (ed) Mammalian protein metabolism. Academic Press, New York, pp 21–132
Kanako O, Motohiko K, Ho A, Kenichi K (2004) Phosphorus application affects root length distribution and water uptake of upland wheat in a column experiment. Soil Sci Plant Nutr 50:257–261
Kimura M (1980) A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J Mol Evol 16:111–120
Kumar S, Tamura K, Nei M (2004) MEGA3: integrated software for molecular evolutionary genetics analysis and sequence alignment. Brief Bioinform 5:150–163
Mehata S, Nautiyal CS (2001) An efficient method for qualitative screening of phosphate-solubilizing bacteria. Curr Microbiol 43:51–56
Murphy JP, Riley JP (1962) A modified single solution method for the determination of the phosphate in natural waters. Anal Chim Acta 27:31–36
Perez E, Sulbaran M, Ball MM, Yarzabal LA (2007) Isolation and characterization of mineral phosphate solubilizing bacteria naturally colonizing a limonitic crust in the south-eastern Venezuela region. Soil Biol Biochem 39:2905–2914
Promod KC, Dhevendaran K (1987) Studies on phospho bacteria in Cochin Backwater. J Mar Biol Assoc India 29:297–305
Ramachandran K, Srinivasan V, Hamza S, Anandaraj M (2007) Phosphate solubilizing bacteria isolated from the rhizosphere soil and its growth promotion on black pepper (Piper nigrum L.) cutting. Plant Soil Sci 102:325–331
Saitou N, Nei M (1987) The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 4:406–425
Seshadri S, Ignacimuthu S, Lakshminarsimhan C (2002) Variations of heterotrophic and phosphate solubilizing bacteria from Chennai, southeast coast of India. Indian J Mar Sci 31(1):69–72
Thingstad TF, Rassoulzadegan F (1995) Nutrient limitations, microbial food webs, and ‘biological pumps’: suggested interactions in a P-limited Mediterranean. Mar Ecol Prog Ser 117:299–306
Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W: improving sensitivity of progressive multiple sequence alignments through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acid Res 22:4673–7680
Vassilev N, Medina A, Azcon R, Vassilev M (2006) Microbial solubilization of rock phosphate on media containing agro-industrial wastes and effect of the resulting products on plant growth and P-uptake. Plant Soil 287:77–84
Vazquez P, Holguin G, Puente ME, Lopez-Cortes A, Bashan Y (2000) Phosphate-solubilizing microorganisms associated with the rhizosphere of mangroves in a semiarid coastal lagoon. Biol Fertil Soils 30:460–468
Venkateswaran K, Natarajan R (1983) Seasonal distribution of inorganic phosphate solubilizing bacteria and phosphatase producing bacteria in Porto Novo waters. Indian J Mar Sci 12:213–217
Whitelaw MA, Harden TJ, Heylar KR (1999) Phosphate solubilization in solution culture by the soil fungus Penicillium radicum. Soil Biol Biochem 31:655–665
Widawati S, Suliasih, Latupuapua HJD, Sugiharto A (2005) Biodiversity of soil microbes from rhizospore at Wamena Biological Garden (WBiG), Jayawijaya, Papua. Biodiversitas 6(1):6–11 (Indonesia)
Widawati S, Suliasih (2006) Augmentation of potential phosphate solubilizing bacteria (PSB) stimulate growth of green mustard (Brassica caventis Ocd.) in marginal soil. Biodiversitas 7(1):10–14 (Indonesia)
Widawati S, Rahmansyah M (2009) The influence of bacterial inoculation to jarak pagar (Jatropa curcas L.) growth. J Biol Indonesia 6(1):107–117 [Indonesia]
Widawati S, Suliasih (2009) The role of phosphate solubilizing bacteria and free living nitrogen fixing bacteria on the growth and adaptation of Gmelina arborea Roxb. grown on degraded land. International seminar to protect diversity of bioresource in the tropical area. Research center for biology Lipi, Cibinong, 25–26 November 2009
Widawati S (2011) Diversity and phosphate solubilization by bacteria isolated from Laki Island coastal ecosystem. Biodiversitas 12(1):17–21 (Indonesia)
Zuberer DA, Silver WS (1979) N2-fixation (acetylene reduction) and the microbial colonization of mangrove roots. New Phytol 82:467–471
Acknowledgments
The authors thank Dr S R Shetye, The Director, the National Institute of Oceanography for providing the necessary facilities and his encouragement. Syed G. Dastager acknowledges the financial supports received under the Grant Nos. OLP 1205 and OLP1209 from the Council of Scientific and Industrial Research (CSIR), New Delhi, India.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Dastager, S.G., Damare, S. Marine Actinobacteria Showing Phosphate-Solubilizing Efficiency in Chorao Island, Goa, India. Curr Microbiol 66, 421–427 (2013). https://doi.org/10.1007/s00284-012-0288-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00284-012-0288-z