
Abstract
Temporal niche partitioning may be influenced not only by interspecific competition, but also by weather conditions. Decreased food availability and dietary overlap between species can increase the degree of interspecific competition during winter, thereby promoting temporal niche partitioning. However, multiple species can be simultaneously active under similar weather conditions (high temperature and little snowfall) in winter to reduce energy costs and increase temporal niche overlap. In this study, we aimed to determine the degree of temporal niche partitioning among red foxes (Vulpes vulpes), raccoon dogs (Nyctereutes procyonoides), and Japanese martens (Martes melampus), and its variation with seasonal climate change in terms of interspecific competition and weather conditions. We obtained data on the target species through a camera-trap survey conducted in a heavy snowfall area in northeastern Japan. We analyzed the degree of temporal niche partitioning based on diel activity overlap, co-occurrence rates per night, and behavioral avoidance within 1 or 2 h. We also evaluated the relationship between the presence or absence of activity per night and nightly weather conditions (temperature, precipitation (snowfall), moonlight). We observed a high degree of temporal niche overlap among the three species. In particular, the degree of temporal niche overlap was higher in winter than that in other seasons because the activity of the three species was greatly affected by low temperatures and snowfall in winter. As a winter survival strategy, coping with the weather may be more important than avoiding competition. Our results conflict with the hypothesis predicting temporal niche partitioning in winter, suggesting that weather effects can be an important factor in varying temporal niche partitioning among carnivores.
Significance statement
This study revealed that the temporal niche overlap of three carnivores in northeastern Japan was higher in winter than that in other seasons because they were simultaneously active under similar weather conditions (high temperature and little snowfall) in winter to reduce energy costs. This indicates that coping with the weather may be more important than avoiding interspecific competition as a winter survival strategy. Our results conflict with the hypothesis predicting temporal niche partitioning among carnivores in winter due to restricted food resources and dietary overlap. This suggests that the different degrees of weather effects can be an important factor in varying temporal niche partitioning among carnivores. Because extreme changes in weather conditions such as extremely high temperatures, blizzards, and heavy rains, have occurred worldwide in recent years, weather conditions may significantly affect the niche partitioning among carnivores distributed in different environments worldwide.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Interspecific competition often occurs among sympatric carnivores (e.g., Durant 1998; Palomares and Caro 1999; Fedriani et al. 2000). The degree of interspecific competition is increased by taxonomic similarity, dietary overlap, and intermediate body mass differences between species (Palomares and Caro 1999; Donadio and Buskirk 2006). For example, larger coyotes (Canis latrans) can exclude smaller swift foxes (Vulpes velox) from the core areas of their home ranges (Kamler et al. 2003). Waggershauser et al. (2021) reported that larger red foxes (Vulpes vulpes) killed smaller pine martens (Martes martes), with a diet similar to that of red foxes, regardless of predation. To avoid such interspecific competition, carnivores need to divide their dietary, spatial (core areas of the home range), and/or temporal niches with those of other species (e.g., Linnell and Strand 2000; Kamler et al. 2012; Monterroso et al. 2014). Such niche partitioning establishes a carnivorous community where multiple sympatric species coexist (e.g., Monterroso et al. 2014). Carnivores influence higher and lower trophic levels through predator–prey interactions, and these effects spread to the structure and functioning of terrestrial ecosystems (Roemer et al. 2009; Ripple et al. 2014); thus, elucidating interspecific competition and niche partitioning is important for understanding carnivore community structure.
Temporal niche partitioning is a most important strategy for coexistence with other species (Monterroso et al. 2014; Frey et al. 2017; Watabe et al. 2022). Previous studies have reported that stone martens (Martes foina) shifted their diel activity patterns to avoid antagonistic encounters with larger European badgers (Meles meles) and European wildcats (Felis silvestris) (Tsunoda et al. 2020; Vilella et al. 2020). Similarly, Gómez-Ortiz et al. (2019) reported the partitioning of diel activity peaks between gray foxes (Urocyon cinereoargenteus) and white-nosed coatis (Nasua narica) with an overlapping dietary niche. Additionally, direct competition can be avoided spatiotemporally using time differences only when the encounter risk is high (e.g., Barrull et al. 2014; Torretta et al. 2016; Karanth et al. 2017; Zalewska et al. 2021). Barrull et al. (2014) showed that stone martens rarely appeared within 60 min of the detection of red foxes and Eurasian badgers at each site, indicating that martens avoided them at fine spatiotemporal scales. Revealing the temporal niche partitioning between species based on diel activities and spatiotemporal scales is important for elucidating the interspecific competition among carnivores (e.g., Barrull et al. 2014; Monterroso et al. 2014).
In general, the behavioral ecology of carnivores, such as their amount of activity (e.g., Doncaster and Macdonald 1997; Kauhala et al. 2007), food resources and diet (e.g., Padial et al. 2002; Baltrūnaitė 2006), home range (e.g., Takeuchi and Koganezawa 1992; Saeki et al. 2007), and diel activity patterns (e.g., Ikeda et al. 2016; Ogurtsov et al. 2018) change seasonally; therefore, the degree of temporal niche partitioning among species varies seasonally. Previous studies have reported that decreased availability of food resources and dietary overlap among species associated with seasonal changes intensified interspecific competition and resulted in increased temporal niche partitioning among species (e.g., Barrull et al. 2014; Monterroso et al. 2016; Petersen et al. 2019). Winter can significantly affect temporal niche partitioning by promoting a decrease in food availability and dietary overlap between species as compared to that in the other seasons. When accompanied by snow cover, the survival rate of rodents is reduced (Korslund and Steen 2006), and rodents hide under the snow, reducing food availability and limiting access for carnivores (Zhou et al. 2011; Willebrand et al. 2017). The restriction of food resources and foraging behavior narrows the dietary niche of carnivores and results in increased dietary overlap among species in winter (e.g., Padial et al. 2002; Carvalho and Gomes 2004; Baltrūnaitė 2006; Murdoch et al. 2010). Consequently, severe winters increase competition for food, which increases the temporal niche partitioning between carnivores to avoid agonistic interactions in winter. This process has been suggested in many studies as a hypothesis (e.g., Barrull et al. 2014; Bu et al. 2016; Torretta et al. 2016; Tsunoda et al. 2020; Vilella et al. 2020; Rossa et al. 2021).
Additionally, weather conditions, such as temperature and precipitation influence the degree of temporal niche partitioning among carnivores and affect the activity of carnivores by inducing thermal stress (Zalewski 2000) and decreasing prey hunting efficiency (Creel et al. 2016). Specifically, in winter, low temperatures cause high thermal stress, and snowfall imposes travel costs and reduces hunting efficiency (Zhou et al. 2011; Willebrand et al. 2017), greatly affecting energy loss for carnivores (Bartoń and Zalewski 2007; Pozzanghera et al. 2016; Mustonen and Nieminen 2018). Previous studies have shown that carnivores, such as Canidae and Martes sp., limit their activity at low temperatures and/or heavy snowfall to reduce energy loss, and increase their activity at high temperatures and/or little snowfall in winter (e.g., Ables 1969; Zalewski 2001; Kauhala et al. 2007). These results indicate that in severe winters, multiple species can be more active simultaneously under similar weather conditions (i.e., high temperature and/or little snowfall) to reduce the energy costs. Therefore, if the cost of coping with weather conditions outweighs the risk of competition from other species, it is possible that weather conditions may lead to temporal niche overlap between species rather than temporal niche partitioning during winter. However, few studies have evaluated the degree of temporal niche partitioning among carnivores from the perspective of both interspecific competition and weather conditions (Frey et al. 2017). Considering the effects of weather conditions and interspecific competition in the assessment of temporal niche partitioning among carnivores is important not only to improve the understanding of carnivore communities, but also to accumulate fundamental information for understanding the effects of climate change on community structure (Herfindal et al. 2017).
We aimed to determine the degree of temporal niche partitioning among carnivores and its variation with seasonal climate change, both in terms of interspecific competition and weather conditions. In this study, we focused on the winter season with large seasonal climate changes and targeted three species in a heavy snowfall area in northeastern Japan, red foxes, raccoon dogs (Nyctereutes procyonoides), and Japanese martens (Martes melampus). These three species are sympatric during the winter and non-winter seasons in the study region (Watabe and Saito 2021a). The diets of these three species are generally similar (Yamamoto 1994; Hisano et al. 2017), and red foxes and raccoon dogs belong to the same family (i.e., Canidae), and they differ in body mass from Japanese martens (Ohdachi et al. 2015); thus, competitive interactions can occur among the three species (Donadio and Buskirk 2006; Hisano et al. 2020). Several studies have reported that red foxes kill Martes in Europe and North America (Lindstrom et al. 1995; Palomares and Caro 1999; Waggershauser et al. 2021). Thus, the degree of temporal niche partitioning among the three species in the study region can vary with season.
We tested two contrasting hypotheses: H1, severe winters with heavy snowfall increase competition for food, which increases temporal niche partitioning among carnivores to avoid agonistic interactions (hereafter “niche-partitioning hypothesis”); and H2, severe winters with heavy snowfall increase temporal niche overlap among carnivores because all species seek to avoid poor environmental conditions due to increased associated energy costs. Previous studies reported a decrease in the availability and use of fruits, insects, rodents, and birds, which are the main winter food resources of these species (Otsu 1972; Misawa 1979; Tsukada and Nonaka 1996; Hirasawa et al. 2006; Adachi et al. 2016), in snowy winters in Japan, suggesting a large overlap in dietary niches among the three species. It has also been confirmed that winter weather conditions affect the activity of red foxes, raccoon dogs, and Martes sp. (e.g., Ables 1969; Zalewski 2001; Kauhala et al. 2007). Therefore, these three species are suitable targets for testing our hypotheses. Furthermore, because the large predator (wolves Canis lupus), which can influence the degree of temporal niche partitioning among the three species, is extinct in Japan (Ohdachi et al. 2015), we excluded the effect of the top predator to reveal interspecific interactions among the three medium-sized carnivores. In this study, we obtained data on the target species through camera-trap surveys and tested two hypotheses by evaluating the degree of temporal niche partitioning based on the diel activity overlap, co-occurrence rates per night, and behavioral avoidance within 1 or 2 h, evaluating the relationship between the presence/absence of activity (detection) per night, and nightly weather conditions (temperature, precipitation (snowfall), moonlight). By testing these two hypotheses and comparing our results with those of previous studies that supported the niche-partitioning hypothesis, we discuss the importance of considering both interspecific competition and weather conditions as factors affecting temporal niche partitioning among carnivores.
Materials and methods
Study area
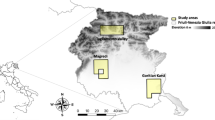
This study was conducted in forests surrounding the Kaminagawa and Nishiaraya areas in Tsuruoka City, Yamagata Prefecture, northeastern Japan, which have a cool temperate climate (Fig. 1). Data were corrected at two sites with different landscape properties. These areas were designated as heavy snowfall areas based on the Act of Special Countermeasures for Heavy Snowfall Areas in Japan (Ministry of Land, Infrastructure, and Transport 2021). Details of these areas are listed in Table S1. In addition to the three target species in this study, seven medium- and large-sized mammals are present in the Kaminagawa area: masked palm civets (Paguma larvata), Japanese badgers (Meles anakuma), Japanese hares (Lepus brachyurus), Japanese macaques (Macaca fuscata), Asiatic black bears (Ursus thibetanus), and Japanese serows (Capricornis crispus) (Watabe et al. 2020). These species were also observed in Nishiaraya during our preliminary surveys.

Study areas and locations of camera-trap sites in Kaminagawa and Nishiaraya areas. The land use data was created from the 1/25000 vegetation map for the period 2001–2012 published by the Ministry of Environment, Japan (Biodiversity Center of Japan 2012)
Camera-trap survey
We conducted camera-trap surveys using infrared-triggered cameras (models #BTC-6HD-940 and #BTC-6HD-APX; Browning, AL, USA) to obtain the target species data. We set 15 camera-trap sites in the Kaminagawa area between February 2019 and August 2021 and nine camera-trap sites in the Nishiaraya area between August 2020 and August 2021 (Fig. 1). In the Kaminagawa area, the number of camera-trap sites and the survey period differed by year (Table S2). The number of camera-trap sites was 7–11 in 2019, 7–12 in 2020, and 10–12 in 2021 for each season. Previous studies have reported that the effective detection distance of animals by infrared-triggered cameras decreases with increased vegetation density (Hofmeester et al. 2017) and that the detection rate of animals by cameras on forest roads is higher than that within forests (Di Bitetti et al. 2014). To mitigate the change in detection distance due to increased vegetation density and because the detection rates of the three focal species on the forest road were high in this study area (Watabe et al. 2020), we installed all cameras on trees along the forest roads. To prevent the cameras from being buried owing to a sudden increase in snow depth, we installed them approximately 150 cm above the ground. We adjusted the height and angle of view of the cameras and checked the battery and operation once or twice per week during the snowfall season and one to three times per month during the other seasons. We programmed the cameras to capture three consecutive images for each trigger with a 1-min delay between each trigger. We identified the focal species from three images and recorded the species, camera-trap site, date, and time as a single detection data. It was not possible to record data blind because our study involved focal animals in the field.
Data analysis
We defined seasons as winter (December–February), spring (March–May), summer (June–August), and autumn (September–November). In each season, we evaluated the degree of temporal niche partitioning between the target species and the effects of weather conditions on the activity of each species. Watabe et al. (2022) argued that evaluation using multiple analytical methods with data from different temporal scales is necessary to understand temporal niche partitioning between carnivores. Therefore, we evaluated niche partitioning on four different temporal scales. R v4.1.0 (R Core Team 2021) was used for all analyses.
We conducted the following two analyses prior to the main analyses described below. When cameras are set close together within a small area, positive spatial autocorrelation can arise because of repeated detections of the species, which can affect our main analyses (Tsunoda et al. 2020; Watabe et al. 2022). First, we used Mantel’s correlogram to assess spatial autocorrelation (Borcard and Legendre 2012). The proximity between cameras did not affect the spatial similarities in the detected species (Fig. S1). Therefore, we concluded that proximal camera-trap placement had little effect on the main analyses. Second, we used Pianka’s index (Pianka 1973) to ensure that spatial use was not biased among the three target species (i.e., no spatial partitioning) in the camera-trap survey area in each study area. We calculated the Pianka’s index for each species-pair using the number of detections of each target species per day per camera. There was no spatial partitioning between the species in each study area (Table S3). Therefore, we confirmed the validity of focusing on the assessment of temporal niche partitioning between the target species in the study areas.
As intermediate body mass differences (Palomares and Caro 1999; Donadio and Buskirk 2006) and high population densities (Karanth et al. 2017) can intensify interspecific competition among carnivores, they may influence the degree of temporal niche partitioning. Therefore, we preliminarily checked for differences in body mass ratios and population densities of the target species in this study and several previous studies supporting the niche-partitioning hypothesis. The population density of each species was calculated using the relative abundance index (RAI), which is the detection frequency per camera per day (O’Brien et al. 2003). Accordingly, the body mass ratios of foxes–martens and raccoon dogs–martens in this study were not significantly different from those of the target species in previous studies (Table S4). The interspecific body mass ratios of carnivores ranged from 2.0 to 5.4 (moderate ratio) (Table S4), which is a particularly high level of interspecific competition in carnivores (Palomares and Caro 1999; Donadio and Buskirk 2006). The RAIs of the three target species in this study tended to be equal to or greater than those of the target species in previous studies (Table S5). Although some studies have noted that RAI does not adequately reflect population density (Burton et al. 2015), the detection frequency of target species by camera-traps in this study was not lower than that of the species in previous studies, supporting the niche-partitioning hypothesis. Therefore, we were confident that the differences in body mass and population density of carnivores were not crucial factors that caused our results to differ from the niche-partitioning hypothesis.
Temporal niche partitioning based on diel activity overlap
We estimated the diel activity patterns of the target species as a probability density function using kernel density estimation (Ridout and Linkie 2009). To determine the overlap of diel activity patterns, we estimated the coefficient of temporal overlap (Δ) for each species-pair, which ranged from 0 (no overlap) to 1 (complete overlap) (Ridout and Linkie 2009; Meredith and Ridout 2021). According to Meredith and Ridout (2021), we used Δ4 if the smaller sample of the two datasets had more than 75 observations; otherwise, Δ1 was used. We defined Δ ≤ 0.5 as low overlap (temporal partitioning), 0.5 < Δ ≤ 0.75 as moderate overlap, and Δ > 0.75 as high overlap (temporal overlap). These thresholds have already been used previously (Monterroso et al. 2014; Marinho et al. 2020; Tsunoda et al. 2020; Panzeri et al. 2021; Viviano et al. 2021; Li et al. 2022; Watabe et al. 2022). To assess the reliability of the estimated Δ statistic, we estimated the mean and 95% confidence intervals (CI) by performing a smoothed bootstrap with 10,000 bootstrap samples (Meredith and Ridout 2021). To reduce duplicate counts of the same individual, we treated consecutive detections of the same species at a camera-trap site within 30 min as a single sample (Monterroso et al. 2014; Watabe and Saito 2021a, b). These analyses were performed using the “overlap” package (Meredith and Ridout 2021) in R.
Temporal niche partitioning based on co-occurrence during one night
We performed temporal co-occurrence analysis (Watabe et al. 2022) to determine the probability that the observed co-occurrence of two species was less than (i.e., spatiotemporal partitioning), greater than (i.e., spatiotemporal overlap), or not different from the expected co-occurrence if the two species occurred independently from each other in units of one night. We analyzed only night-time data because the three target species were mainly nocturnal in this study area (Watabe and Saito 2021a,b). We used probabilistic co-occurrence analysis for temporal co-occurrence analysis using a matrix containing the detection-nondetection (1/0) data per night at each camera site for each species pair (Griffith et al. 2016; Watabe et al. 2022). First, the observed co-occurrence rate for each species was calculated by dividing the number of detections for one species by the total number of nights at each site. Second, the expected co-occurrence was calculated by multiplying the observed co-occurrence rate of one species, the observed co-occurrence rate of the other species, and the total number of nights at each site. Finally, we compared the calculated expected co-occurrences with the number of observed co-occurrences. We also evaluated the degree of partitioning or overlap (co-occurrence) for significant pairs based on the value of the observed co-occurrence/expected co-occurrence. A smaller value indicates a high degree of partitioning and a larger value indicates a high degree of overlap. We defined one night as 60 min before sunset to 60 min after sunrise. To correct for multiple comparisons, we used a significance level adjusted using the Holm method. We obtained the sunrise and sunset times for each survey day using “rSetDayNightAttr” package (Furukawa 2019) in R. The analysis was performed using the “cooccur” (Griffith et al. 2016) package in R.
Temporal niche partitioning based on behavioral avoidance of fine scales
We performed time-to-event analyses (Cusack et al. 2017; Balme et al. 2019; Davis et al. 2021) to assess the behavioral avoidance of other species on fine spatiotemporal scales. Barrull et al. (2014) indicated that smaller martens may detect odors from larger competitors (foxes and badgers) within 1 h and avoid them on fine spatiotemporal scales. We assumed that the odors of carnivores would remain in the environment for several hours and used a spatiotemporal scale within 1 and 2 h. First, we created a matrix of detection records for each species comprising of the camera-trap site and date-time. We calculated the minimum time difference between each detection record of a particular species (after-species) and the former detection records of a particular species (before-species) for each camera-trap site. Among the calculated minimum time differences, we obtained the number of detections within 1 or 2 h (the observed number of detections). Second, we randomly permutated all the detection records of the after-species. For each detection record, we calculated the minimum time difference from the detection records of the before-species used in the first calculation (Watabe et al. 2022) and obtained the number of detections within 1 or 2 h of the calculated minimum time differences. We repeated this process 1000 times and obtained 1000 simulated data points for the number of detections. We carried out the first and second procedures for each season of the year and added the number of detections for each season. Finally, following the method reported by Cusack et al. (2017), we calculated the p value as the percentage of simulated data points where the observed number of detections was greater than or equal to the simulated number of detections obtained using random permutations. A p value < 0.05 (i.e., significantly fewer detections observed within 1 or 2 h than expected) represents behavioral avoidance from other species (Cusack et al. 2017; Balme et al. 2019). In a previous study that conducted a time-to-event analysis (e.g., Balme et al. 2019), consecutive detections of the same species less than 1 min apart were collapsed into a single sample. In this study, we set the interval time of each camera to 1 min; therefore, we used all detection data for this analysis.
Effects of weather conditions on activity of carnivores
To evaluate the effects of weather conditions on the activity of each target species, we constructed a generalized linear mixed model (GLMM). The presence/absence of detection (1/0) per night at each camera site was used as the response variable. We defined one night as the same night as the temporal co-occurrence analysis. The response variable was assumed to follow a binomial distribution and logit was used as the link function. Because temperature and precipitation increases energy costs for carnivores and affect their activities (e.g., Ables 1969; Kauhala et al. 2007), we used mean night-time temperature (°C) and hourly precipitation (mm) as explanatory variables. In the Kaminagawa area, we created these variables based on hourly temperature and precipitation data from meteorological observation data of the Kaminagawa Experimental Forest of Yamagata University, which is in this area. In the Nishiaraya area, we used hourly temperature and precipitation data from the closest meteorological station to this area, the Japan Meteorological Agency Tsuruoka Station (38°44′N, 139°49′E) (Japan Meteorological Agency 2021). Owing to the positive correlation between the mean night-time hourly precipitation and the increase in snow depth in winter (Pearson’s correlation coefficient r = 0.66), we considered that the precipitation in winter reflected the increase in snow depth (snowfall). Moonlight is another factor related to energy loss that affects the activity and interspecific relationships of carnivores as it increases the hunting efficiency of predators and its effects may spill over into the degree of niche partitioning (e.g., Penteriani et al. 2013; Prugh and Golden 2014; Leonard et al. 2020). Therefore, we included moonlight as a confounding factor among the explanatory variables. We created this variable from continuous value data on the age of the moon (National Astronomical Observatory of Japan 2021) and represented moonlight by the phases of the moon (e.g., Penteriani et al. 2013). Because the age of the moon is expressed as a continuous value in the range of approximately 0 (new moon) to 30 (new moon) with the full moon around 15 in the center, we adjusted the data as a continuous value in the range of 0 (new moon) to 15 (full moon) and used this data as the variable. We standardized the explanatory variables and assessed their influence by using the absolute values of the standardized regression coefficients. To consider multicollinearity in the model construction, we checked Pearson’s correlation coefficient (r) among the explanatory variables. As the absolute value of r was < 0.1 across all variables, we assumed that multicollinearity would not occur.
We used two random effects to construct the GLMM: random intercepts and random slopes. We used year–month (e.g., 2019–01), and camera-trap sites as random intercepts, and used area–season (e.g., Kaminagawa–winter) as random slopes. We evaluated the differences in the relative influence of variables by area and season using standardized regression coefficients of the fixed effect with random slopes. We obtained the regression coefficients and 95% CIs for each variable using 5000 iterations of the nonparametric bootstrap method. When the 95% CI of the coefficient did not overlap zero, the variable was considered statistically significant. The R package “glmmTMB” package (Magnusson et al. 2021) in R was used for the GLMM analysis.
Results
Camera-trap survey
The total duration of the camera-trap survey was 10,068 camera-trap days, excluding the number of days when the angle of the cameras was changed because of bear attacks or camera troubles. We detected 2519, 1810, and 1279 red foxes, raccoon dogs, and Japanese martens, respectively (Table 1). The dataset for the analyses of temporal niche partitioning and GLMM is included in the Supplementary Information (Table S11).
Temporal niche partitioning based on diel activity overlap
Kernel density estimations showed that all species were nocturnal in all areas and seasons (Fig. 2). For all species pairs and seasons, the mean Δ was higher than 0.5, indicating moderate or high diel activity overlap among the species (Fig. 2).

Diel activity patterns and the coefficient of temporal overlap (Δ) for each species-pair. RF, RD, and JM represent the red fox, raccoon dog, and Japanese marten, respectively. The mean Δ and the confidence interval are indicated in the upper right of each figure. The gray area indicates the overlap between diel activity patterns. The rug at the bottom of the plot indicates the original observations of activity time
For all species pairs and seasons, except for the foxes and martens in summer, the mean Δ was > 0.75 in both or one of the areas, indicating high diel activity overlap among the species (Fig. 2). In autumn in both areas, and in winter and spring in the Kaminagawa area, the mean Δ was relatively high for all species pairs (Fig. 2).
Temporal niche partitioning based on co-occurrence during one night
The results of the probabilistic co-occurrence analysis showed that the observed co-occurrence frequency was not significantly lower than the expected frequency for all seasons and species pairs in both areas (Table 2). Overall, the observed co-occurrence frequency was significantly greater than the expected frequency for multiple-species pairs, indicating co-occurrence per night (Table 2).
In winter, all species pairs showed significant co-occurrence in both areas (Table 2). Spring had the second highest number of species pairs, and both analyses showed a significant co-occurrence (Table 2). In summer, raccoon dogs and martens in both areas showed a significant co-occurrence (Table 2). In autumn, only the foxes and martens in the Nishiaraya area showed a significant co-occurrence (Table 2). Except for a few results, the value of the observed co-occurrence/expected co-occurrence was relatively higher in winter than in the other seasons in both areas (Table 2).
Temporal niche partitioning based on behavioral avoidance of fine scales
The results of the time-to-event analysis within 1 h showed that the observed number of detections was significantly lower than that expected for one species-pair (raccoon dog–marten in autumn in the Nishiaraya area); however, it was not significantly different for all other seasons and pairs (Fig. 3; Table S7). These results were the same for the time-to-event analysis within 2 h (Fig. 3; Table S7).

Frequency distribution of the 1000 simulated data of the number of detections and significant p-value estimated by time-to-event analysis within one or two hours. The dark gray of the distribution indicates the location of the observed number of detections. The vertical axis represents the frequency of simulated number of detections. The horizontal axis represents the number of detections. RF, RD, and JM represent the red fox, raccoon dog, and Japanese marten, respectively. RF-RD represents that raccoon dogs were detected after red foxes, while RD-RF represents that red foxes were detected after raccoon dogs (i.e., before-after). Asterisk* represents a p value < 0.05
Effects of weather conditions on activity of carnivores
The model showed that, in both areas, temperature had significant positive and negative effects on the nighttime detection of red foxes in winter and summer, respectively (Fig. 4). Precipitation (snowfall) had a significant negative effect in all areas and seasons, except in summer in the Nishiaraya area (Fig. 4). Although moonlight had a significant positive or negative effect in autumn, the effect was relatively small (Fig. 4). Overall, the effects of temperature and precipitation (snowfall) during winter were relatively large in both areas (Fig. 4).

Standardized regression coefficients and the 95% confidence intervals (CIs) of the fixed effect with random slopes per area–season in the generalized linear mixed model (GLMM) for the effects of weather conditions on species activity. Medians and 95% CIs for parameters were estimated by a nonparametric bootstrap method with 5000 iterations. RF, RD, and JM represent the red fox, raccoon dog, and Japanese marten, respectively. Asterisks* indicate significant differences (95% CIs did not exceed zero)
The model showed that temperature had a significant positive effect on the nighttime detection of raccoon dogs in winter in both areas (Fig. 4). Precipitation (snowfall) had a significant negative effect in winter and spring in the Kaminagawa area (Fig. 4). In the Nishiaraya area, precipitation had a significant negative effect in all seasons, except winter (Fig. 4). Overall, the effects of temperature in winter were the largest in both areas (Fig. 4), and those of precipitation (snowfall) in winter in the Kaminagawa area and spring in the Nishiaraya area were relatively large (Fig. 4).
The model showed that temperature had a significantly positive effect on the nighttime detection of Japanese martens in winter in both areas (Fig. 4). Precipitation (snowfall) had a significant negative effect in all areas and seasons, except in summer in the Nishiaraya area (Fig. 4). Moonlight had a significant negative effect in autumn in both areas (Fig. 4). Overall, compared to that of the nighttime detection of the other two species, temperature and precipitation had a similar degree of influence on the nighttime detection of martens in each season (Fig. 4).
Discussion
The present study showed a low degree of temporal niche partitioning between species at any scale, season, or area (Table 3) except for one species-pair at a fine scale (Fig. 3; Table S7). Multiple species pairs showed had high degree of temporal niche overlap (Table 3). In all seasons, many pairs showed a low degree of diel activity partitioning and a high degree of diel activity overlap (Fig. 2). In both areas, more pairs showed co-occurrence per night in winter than that in the other seasons (Table 2), and the degree of overlap (co-occurrence) was relatively higher in winter than in the other seasons, with a few exceptions. The possibility of finer scale partitioning at night in winter was rejected by the time-to-event analyses (Fig. 3; Table S7). These results indicate that the degree of temporal niche overlap in winter was higher than that in the other seasons. Low precipitation (snowfall) and high temperatures in winter commonly contributed to an increase in the activity of the three species, and the degree of their influence was relatively high (Fig. 4). These results suggest that the three species in this study are more concerned with responding to weather conditions than avoiding competition with other species. Therefore, our results indicate that the three species were more active under similar weather conditions during winter, resulting in a high degree of temporal niche overlap among the species in winter, supporting H2, but not H1. To understand carnivore community structure based on temporal niche partitioning, the effects of interspecific competition and weather conditions on the degree of temporal niche partitioning should be considered.
Previous studies have reported that raccoon dogs are active when the temperature is high during the winter (Kauhala et al. 2007; Seki and Koganezawa 2011; Zoller and Drygala 2013). Zalewski (2001) have reported that Martes sp. were less active during cold winter nights. In addition, deep and fluffy snow limits the travel of red foxes and raccoon dogs (Ables 1969; Kauhala et al. 2007; Pozzanghera et al. 2016) and reduces the foraging efficiency of red foxes and Martes sp. (e.g., Willebrand et al. 2017). These results suggest that red foxes, raccoon dogs, and Martes sp. are active at high temperatures and/or with little snowfall for energetically efficient travel, which is consistent with the results of this study. The reduced activity of the three target species during daytime (Fig. 2) may be due to the evolutionary fixation of their circadian rhythms as nocturnal, even though it is warmer during the day (Kronfeld-Schor and Dayan 2003). Thus, they may be active at night under weather conditions with less energy loss. Additionally, severe winter with snowfall greatly reduces the activity level (raccoon dogs, Kitao et al. 2009; Mustonen and Nieminen 2018) and survival rates (red foxes, Bartoń and Zalewski 2007) of carnivores. The relative detection frequency of raccoon dogs was the lowest in winter (Table 1), which might reflect a decreased activity level. As a survival strategy in winter for the three species in this study, coping with the weather may be more important than avoiding competition with other species.
While many previous studies have supported the niche-partitioning hypothesis (e.g., Barrull et al. 2014; Bu et al. 2016; Torretta et al. 2016; Tsunoda et al. 2020; Vilella et al. 2020; Rossa et al. 2021), our results showed a high degree of temporal niche overlap in winter. Since differences in body mass and population density among species were less related to the differences in these results (Tables S4, S5), winter environmental severity could be related to the differences in our results from the niche-partitioning hypothesis. This study was conducted in a heavy snowfall area with a maximum snow depth > 200 cm (Ministry of Land Infrastructure and Transport 2021; Watabe and Saito 2021b). The winter weather conditions in this area can be more severe than those in areas where previous studies have supported the niche-partitioning hypothesis because of the heavy snowfall. Based on these environmental differences, the degree of interspecific competition and the cost of adaptation to weather can vary depending on the extent of the weather impact. The survival strategy of carnivores (competition avoidance or weather adaptation) can shift depending on the extent of weather impact. The temporal niches of the three species in this study could have overlapped rather than partitioned, because the cost of adaptation to weather conditions in severe winters exceeded the risk of competition.
The diversity of active carnivorous species in winter may also be related to the differences in the results from the niche-partitioning hypothesis. In our study area, the activity levels of Japanese badgers and masked palm civets (medium-sized carnivores) substantially decreased during winter (Watabe and Saito 2022; MUS et al. unpublished data). In contrast, in southwestern Europe, Eurasian badgers are more active in winter than Japanese badgers living in our study area, and diverse species exist within the carnivore guild, including European wildcats, European genets (Genetta genetta), and top predators such as jackals and wolves (e.g., Torretta et al. 2016; Tsunoda et al. 2020; Vilella et al. 2020). Monterroso et al. (2014) suggested that increased diversity within carnivore guilds result in increased temporal niche partitioning between species. The high diversity of species that are more active in winter may be one of the factors that promote temporal niche partitioning. This suggests that multispecies interactions are intricately intertwined within carnivore guilds. Such interactions should be evaluated to assess the temporal niche partitioning among carnivores in the future.
A high degree of temporal niche overlap among the species was also observed in seasons other than winter (Table 3). The spring results in Kaminagawa might have shown similar trends to winter because of the remnants of the winter conditions. Because precipitation affects heat loss in wildlife (Hendrichsen and Tyler 2014), the fact that the three species commonly tended to be active at night when precipitation was low (Fig. 4) could be a factor in niche overlap. In addition, because carnivores synchronize their activity patterns with those of their prey species (Dias et al. 2019; Botts et al. 2020), the three species in this study might synchronize their activity patterns with those of their common prey, such as small mammals (e.g., Yamamoto 1994). As weather conditions influence the prey abundance and hunting efficiency of carnivores (Creel et al. 2016; Parsons et al. 2020), the weather conditions may indirectly influence the degree of niche partitioning among carnivores through prey species. In the future, it will be necessary to clarify the correlation between weather effects and activity levels at more finer scales and, the density and activity patterns of carnivore prey species. Thus, we can better understand the factors that contribute to the degree of temporal niche partitioning among carnivores.
This study showed a trend of toward a high degree of temporal niche overlap among three carnivorous species: red foxes, raccoon dogs, and Japanese martens. Our results indicated that the degree of temporal niche overlap among the three species was higher in winter than that in other seasons because the activity of the three species was greatly affected by winter weather conditions. As our results showed a common trend in the two different landscapes, we could have provided findings with some generality. Our results did not support the niche-partitioning hypothesis, suggesting that different degrees of weather effects can be an important factor in varying the temporal niche partitioning among carnivores. The influence of weather conditions on carnivores is known not only in heavy snowy environments, such as in this study area, but also in areas with little snowfall (e.g., Zalewski 2000; Zoller and Drygala 2013). Weather conditions may influence the degree of temporal niche partitioning among carnivores in areas where environmental changes associated with seasonal changes are observed, even in low snowfall environments. Furthermore, climate change could increase the degree of future weather variability. The World Meteorological Organization (2020) has reported that extreme changes in weather conditions, such as extremely high temperatures, blizzards, and heavy rains, have occurred worldwide in recent years. Such extreme weather conditions may significantly affect the activity level and foraging efficiency of carnivores and alter the degree of niche partitioning between species. Many carnivorous species are distributed in different environments worldwide (Davis et al. 2018). Thus, in various regions, it is important to clarify the temporal niche partitioning among carnivores in terms of interspecific relationships and weather conditions to understand carnivore community structures and their changes.
Data availability
The datasets used and/or analyzed during the current study are available from the supplementary information.
References
Ables ED (1969) Activity studies of red foxes in southern Wisconsin. J Wildlife Manage 33:145–153
Adachi T, Uehara A, Kuwahara Y, Takatsuki S (2016) Seasonal food habits of the Japanese marten (Martes melampus melampus) at Otome Highland, central Japan. Mamm Sci 56:17–25 (in Japanese with English summary)
Balme G, Rogan M, Thomas L et al (2019) Big cats at large: density, structure, and spatio-temporal patterns of a leopard population free of anthropogenic mortality. Popul Ecol 61:256–267. https://doi.org/10.1002/1438-390X.1023
Baltrūnaitė L (2006) Diet and winter habitat use of the red fox, pine marten and raccoon dog in Dzūkija National Park, Lithuania. Acta Zool Litu 16:46–53. https://doi.org/10.1080/13921657.2006.10512709
Barrull J, Mate I, Ruiz-Olmo J, Casanovas JG, Gosàlbez J, Salicrú M (2014) Factors and mechanisms that explain coexistence in a Mediterranean carnivore assemblage: an integrated study based on camera trapping and diet. Mamm Biol 79:123–131. https://doi.org/10.1016/j.mambio.2013.11.004
Bartoń KA, Zalewski A (2007) Winter severity limits red fox populations in Eurasia. Glob Ecol Biogeogr 16:281–289. https://doi.org/10.1111/j.1466-8238.2007.00299.x
Borcard D, Legendre P (2012) Is the Mantel correlogram powerful enough to be useful in ecological analysis? A simulation study. Ecology 93:1473–1481. https://doi.org/10.1890/11-1737.1
Botts RT, Eppert AA, Wiegman TJ et al (2020) Circadian activity patterns of mammalian predators and prey in Costa Rica. J Mammal 101:1313–1331. https://doi.org/10.1093/jmammal/gyaa103
Bu H, Wang F, McShea WJ, Lu Z, Wang D, Li S (2016) Spatial co-occurrence and activity patterns of mesocarnivores in the temperate forests of southwest China. PLoS ONE 11:e0164271. https://doi.org/10.1371/journal.pone.0164271
Burton AC, Neilson E, Moreira D, Ladle A, Steenweg R, Fisher JT, Bayne E, Boutin S (2015) Wildlife camera trapping: a review and recommendations for linking surveys to ecological processes. J Appl Ecol 52:675–685. https://doi.org/10.1111/1365-2664.12432
Carvalho JC, Gomes P (2004) Feeding resource partitioning among four sympatric carnivores in the Peneda-Gerês National Park (Portugal). J Zool 263:275–283. https://doi.org/10.1017/S0952836904005266
Biodiversity Center of Japan (2012) The 6–7th National Surveys on The Natural Environment, https://www.biodic.go.jp/kiso/fnd_list_h.html
Creel S, Creel NM, Creel AM, Creel BM (2016) Hunting on a hot day: effects of temperature on interactions between African wild dogs and their prey. Ecology 97:2910–2916. https://doi.org/10.1002/ecy.1568
Cusack JJ, Dickman AJ, Kalyahe M, Rowcliffe JM, Carbone C, MacDonald DW, Coulson T (2017) Revealing kleptoparasitic and predatory tendencies in an African mammal community using camera traps: a comparison of spatiotemporal approaches. Oikos 126:812–822. https://doi.org/10.1111/oik.03403
Davis CL, Rich LN, Farris ZJ et al (2018) Ecological correlates of the spatial co-occurrence of sympatric mammalian carnivores worldwide. Ecol Lett 21:1401–1412. https://doi.org/10.1111/ele.13124
Davis RS, Yarnell RW, Gentle LK et al (2021) Prey availability and intraguild competition regulate the spatiotemporal dynamics of a modified large carnivore guild. Ecol Evol 11:7890–7904. https://doi.org/10.1002/ece3.7620
Di Bitetti MS, Paviolo A, De Angelo C (2014) Camera trap photographic rates on roads vs. off roads: location does matter. Mastozool Neotrop 21:37–46
Dias DM, Massara RL, de Campos CB, Rodrigues FHG (2019) Feline predator–prey relationships in a semi-arid biome in Brazil. J Zool 307:282–291. https://doi.org/10.1111/jzo.12647
Donadio E, Buskirk SW (2006) Diet, morphology, and interspecific killing in carnivora. Am Nat 167:524–536. https://doi.org/10.1086/501033
Doncaster CP, Macdonald DW (1997) Activity patterns and interactions of red foxes (Vulpes vulpes) in Oxford city. J Zool 241:73–87. https://doi.org/10.1111/j.1469-7998.1997.tb05500.x
Durant SM (1998) Competition refuges and coexistence: an example from Serengeti carnivores. J Anim Ecol 67:370–386. https://doi.org/10.1046/j.1365-2656.1998.00202.x
Fedriani JM, Fuller TK, Sauvajot RM, York EC (2000) Competition and intraguild predation among three sympatric carnivores. Oecologia 125:258–270. https://doi.org/10.1007/s004420000448
Frey S, Fisher JT, Burton AC, Volpe JP (2017) Investigating animal activity patterns and temporal niche partitioning using camera-trap data: challenges and opportunities. Remote Sens Ecol Conserv 3:123–132. https://doi.org/10.1002/rse2.60
Furukawa G (2019) genkiFurukawa/rSetDayNightAttr documentation, https://rdrr.io/github/genkiFurukawa/rSetDayNightAttr/
Gómez-Ortiz Y, Monroy-Vilchis O, Castro-Arellano I (2019) Temporal coexistence in a carnivore assemblage from central Mexico: temporal-domain dependence. Mammal Res 64:333–342. https://doi.org/10.1007/s13364-019-00415-8
Griffith DM, Veech JA, Marsh CJ (2016) Cooccur: probabilistic species co-occurrence analysis in R. J Stat Softw 69:1–17. https://doi.org/10.18637/jss.v069.c02
Hendrichsen DK, Tyler NJC (2014) How the timing of weather events influences early development in a large mammal. Ecology 95:1737–1745. https://doi.org/10.1890/13-1032.1
Herfindal I, Lande US, Solberg EJ, Rolandsen CM, Roer O, Wam HK (2017) Weather affects temporal niche partitioning between moose and livestock. Wildlife Biol 2017:wlb.00275. https://doi.org/10.2981/wlb.00275
Hirasawa M, Kanda E, Takatsuki S (2006) Seasonal food habits of the raccoon dog at a western suburb of Tokyo. Mamm Study 31:9–14. https://doi.org/10.3106/1348-6160(2006)31[9:sfhotr]2.0.co;2
Hisano M, Hoshino L, Kamada S, Masuda R, Newman C, Kaneko Y (2017) A comparison of visual and genetic techniques for identifying Japanese marten scats enabling diet examination in relation to seasonal food availability in a sub-alpine area of Japan. Zool Sci 34:137–146
Hisano M, Ca M, Willcox D (2020) Toward a better understanding of the Japanese marten Martes melampus diet. Small Carniv Conserv 58:e58009
Hofmeester TR, Rowcliffe JM, Jansen PA (2017) A simple method for estimating the effective detection distance of camera traps. Remote Sens Ecol Conserv 3:81–89. https://doi.org/10.1002/rse2.25
Ikeda T, Uchida K, Matsuura Y, Takahashi H, Yoshida T, Kaji K, Koizumi I (2016) Seasonal and diel activity patterns of eight sympatric mammals in northern Japan revealed by an intensive camera-trap survey. PLoS ONE 11:e0163602. https://doi.org/10.1371/journal.pone.0163602
Japan Meteorological Agency (2021) Japan Meteorological Agency. Yearly climate data in Tsuruoka, https://www.data.jma.go.jp/gmd/risk/obsdl/index.php
Kamler JF, Ballard WB, Gilliland RL, Mote K (2003) Spatial relationships between swift foxes and coyotes in northwestern Texas. Can J Zool 81:168–172. https://doi.org/10.1139/z02-222
Kamler JF, Stenkewitz U, Klare U, Jacobsen NF, Macdonald DW (2012) Resource partitioning among cape foxes, bat-eared foxes, and black-backed jackals in South Africa. J Wildlife Manage 76:1241–1253. https://doi.org/10.1002/jwmg.354
Karanth KU, Srivathsa A, Vasudev D, Puri M, Parameshwaran R, Kumar NS (2017) Spatio-temporal interactions facilitate large carnivore sympatry across a resource gradient. Proc R Soc B 284:20161860. https://doi.org/10.1098/rspb.2016.1860
Kauhala K, Holmala K, Schregel J (2007) Seasonal activity patterns and movements of the raccoon dog, a vector of diseases and parasites, in southern Finland. Mamm Biol 72:342–353. https://doi.org/10.1016/j.mambio.2006.10.006
Kitao N, Fukui D, Hashimoto M, Osborne PG (2009) Overwintering strategy of wild free-ranging and enclosure-housed Japanese raccoon dogs (Nyctereutes procyonoides albus). Int J Biometeorol 53:159–165. https://doi.org/10.1007/s00484-008-0199-7
Korslund L, Steen H (2006) Small rodent winter survival: snow conditions limit access to food resources. J Anim Ecol 75:156–166. https://doi.org/10.1111/j.1365-2656.2005.01031.x
Kronfeld-Schor N, Dayan T (2003) Partitioning of time as an ecological resource. Annu Rev Ecol Evol S 34:153–181. https://doi.org/10.1146/132435
Leonard JP, Tewes ME, Lombardi JV, Wester DW, Campbell TA (2020) Effects of sun angle, lunar illumination, and diurnal temperature on temporal movement rates of sympatric ocelots and bobcats in South Texas. PLoS ONE 15:e0231732. https://doi.org/10.1371/journal.pone.0231732
Li J, Xue Y, Liao M, Dong W, Wu B, Li D (2022) Temporal and spatial activity patterns of sympatric wild ungulates in Qinling Mountains. China Animals 12:1666. https://doi.org/10.3390/ani12131666
Lindstrom ER, Brainerd SM, Helldin JO, Overskaug K (1995) Pine marten-red fox interactions: a case of intraguild predation? Ann Zool Fenn 32:123–130
Linnell JDC, Strand O (2000) Interference interactions, co-existence and conservation of mammalian carnivores. Divers Distrib 6:169–176. https://doi.org/10.1046/j.1472-4642.2000.00069.x
Magnusson A, Skaug H, Nielsen A, et al (2021) Generalized linear mixed models using template model builder, https://cran.r-project.org/web/packages/glmmTMB/index.html
Marinho PH, Fonseca CR, Sarmento P, Fonseca C, Venticinque EM (2020) Temporal niche overlap among mesocarnivores in a Caatinga dry forest. Eur J Wildlife Res 66:34. https://doi.org/10.1007/s10344-020-1371-6
Meredith M, Ridout M (2021) Overlap: estimates of coefficient of overlapping for animal activity patterns, https://cran.r-project.org/web/packages/overlap/index.html
Ministry of Land Infrastructure and Transport (2021) The act of special countermeasures for heavy snowfall area, https://www.mlit.go.jp/kokudoseisaku/chisei/crd_chisei_tk_000010.html
Misawa E (1979) Change in the food habits of the red fox, Vulpes vulpes schrencki Kishida according to habitat conditions. J Mammal Soc Japan 7:311–320 (in Japanese)
Monterroso P, Alves PC, Ferreras P (2014) Plasticity in circadian activity patterns of mesocarnivores in southwestern Europe: implications for species coexistence. Behav Ecol Sociobiol 68:1403–1417. https://doi.org/10.1007/s00265-014-1748-1
Monterroso P, Rebelo P, Alves PC, Ferreras P (2016) Niche partitioning at the edge of the range: a multidimensional analysis with sympatric martens. J Mammal 97:928–939. https://doi.org/10.1093/jmammal/gyw016
Murdoch JD, Munkhzul T, Buyandelger S, Reading RP, Sillero-Zubiri C (2010) Seasonal food habits of corsac and red foxes in Mongolia and the potential for competition. Mamm Biol 75:36–44. https://doi.org/10.1016/j.mambio.2008.12.003
Mustonen AM, Nieminen P (2018) A review of the physiology of a survival expert of big freeze, deep snow, and an empty stomach: the boreal raccoon dog (Nyctereutes procyonoides). J Comp Physiol B 188:15–25. https://doi.org/10.1007/s00360-017-1114-5
National Astronomical Observatory of Japan (2021) Ephemeris computation office NAOJ, https://eco.mtk.nao.ac.jp/koyomi/index.html
O’Brien TG, Kinnaird MF, Wibisono HT (2003) Crouching tigers, hidden prey: Sumatran tiger and prey populations in a tropical forest landscape. Anim Conserv 6:131–139. https://doi.org/10.1017/S1367943003003172
Ogurtsov SS, Zheltukhin AS, Kotlov IP (2018) Daily activity patterns of large and medium-sized mammals based on camera traps data in the Central Forest Nature Reserve, Valdai Upland, Russia. Nat Conserv Res 3:68–88. https://doi.org/10.24189/ncr.2018.031
Ohdachi SD, Ishibashi Y, Iwasa MA, Fukuki D, Saitoh T (2015) The wild mammals of Japan, 2nd edn. Shokadoh Book Seller, Kyoto
Otsu S (1972) Winter food of Japanese yellow marten, Martes melumpus melumpus (Temminck et Schelegel), in Yamagata Prefecture. Japn J Appl Entomol Zool 16:75–78 (in Japanese with English summary)
Padial JM, Âvila E, Sánchez JM (2002) Feeding habits and overlap among red fox (Vulpes vulpes) and stone marten (Martes foina) in two Mediterranean mountain habitats. Mamm Biol 67:137–146. https://doi.org/10.1078/1616-5047-00021
Palomares F, Caro TM (1999) Interspecific killing among mammalian carnivores. Am Nat 153:492–508. https://doi.org/10.1086/303189
Panzeri M, Mazza G, Bisi F, Mori E (2021) Patterns of spatiotemporal activity of an alien lagomorph inferred through camera-trapping. Mammal Res 66:281–288. https://doi.org/10.1007/s13364-021-00557-8
Parsons MA, Bridges AS, Biteman DS, Garcelon DK (2020) Precipitation and prey abundance influence food habits of an invasive carnivore. Anim Conserv 23:60–71. https://doi.org/10.1111/acv.12510
Penteriani V, Kuparinen A, del Mar DM et al (2013) Responses of a top and a meso predator and their prey to moon phases. Oecologia 173:753–766. https://doi.org/10.1007/s00442-013-2651-6
Petersen WJ, Savini T, Steinmetz R, Ngoprasert D (2019) Periodic resource scarcity and potential for interspecific competition influences distribution of small carnivores in a seasonally dry tropical forest fragment. Mamm Biol 95:112–122. https://doi.org/10.1016/j.mambio.2018.11.001
Pianka ER (1973) The structure of lizard communities. Annu Rev Ecol Syst 4:53–74. https://doi.org/10.1146/annurev.es.04.110173.000413
Pozzanghera CB, Sivy KJ, Lindberg MS, Prugh LR (2016) Variable effects of snow conditions across boreal mesocarnivore species. Can J Zool 94:697–705. https://doi.org/10.1139/cjz-2016-0050
Prugh LR, Golden CD (2014) Does moonlight increase predation risk? Meta-analysis reveals divergent responses of nocturnal mammals to lunar cycles. J Anim Ecol 83:504–514. https://doi.org/10.1111/1365-2656.12148
R Core Team (2021) R: a language environment for statistical computing. R foundation for statistical computing, Vienna, Austria, https://www.r-project.org/
Ridout MS, Linkie M (2009) Estimating overlap of daily activity patterns from camera trap data. J Agric Biol Environ Stat 14:322–337. https://doi.org/10.1198/jabes.2009.08038
Ripple WJ, Estes JA, Beschta RL et al (2014) Status and ecological effects of the world’s largest carnivores. Science 343:1241484. https://doi.org/10.1126/science.1241484
Roemer GW, Gompper ME, Van Valkenburgh B (2009) The ecological role of the mammalian mesocarnivore. Bioscience 59:165–173. https://doi.org/10.1525/bio.2009.59.2.9
Rossa M, Lovari S, Ferretti F (2021) Spatiotemporal patterns of wolf, mesocarnivores and prey in a Mediterranean area. Behav Ecol Sociobiol 75:32. https://doi.org/10.1007/s00265-020-02956-4
Saeki M, Johnson PJ, Macdonald DW (2007) Movements and habitat selection of raccoon dogs (Nyctereutes procyonoides) in a mosaic landscape. J Mammal 88:1098–1111. https://doi.org/10.1644/06-mamm-a-208r1.1
Seki Y, Koganezawa M (2011) Factors influencing winter home ranges and activity patterns of raccoon dogs Nyctereutes procyonoides in a high-altitude area of Japan. Acta Theriol 56:171–177. https://doi.org/10.1007/s13364-010-0020-y
Takeuchi M, Koganezawa M (1992) Home range and habitat utilisation of the red fox Vulpes vulpes in the Ashio Mountains, Central Japan. J Mammal Soc Japan 17:95–110. https://doi.org/10.11238/jmammsocjapan.17.95
Torretta E, Serafini M, Puopolo F, Schenone L (2016) Spatial and temporal adjustments allowing the coexistence among carnivores in Liguria (N-W Italy). Acta Ethol 19:123–132. https://doi.org/10.1007/s10211-015-0231-y
Tsukada H, Nonaka N (1996) Foraging behavior of red foxes Vulpes vulpes schrencki utilizing human food in the Shiretoko National Park, Hokkaido. Mamm Study 21:137–151
Tsunoda H, Newman C, Peeva S, Raichev E, Buesching CD, Kaneko Y (2020) Spatio-temporal partitioning facilitates mesocarnivore sympatry in the Stara Planina Mountains. Bulgaria Zoology 141:125801. https://doi.org/10.1016/j.zool.2020.125801
Vilella M, Ferrandiz-Rovira M, Sayol F (2020) Coexistence of predators in time: effects of season and prey availability on species activity within a Mediterranean carnivore guild. Ecol Evol 10:11408–11422. https://doi.org/10.1002/ece3.6778
Viviano A, Mori E, Fattorini N, Mazza G, Lazzeri L, Panichi A, Strianese L, Mohamed WF (2021) Spatiotemporal overlap between the European brown hare and its potential predators and competitors. Animals 11:562. https://doi.org/10.3390/ani11020562
Waggershauser CN, Ruffino L, Kortland K, Lambin X (2021) Lethal interactions among forest-grouse predators are numerous, motivated by hunger and carcasses, and their impacts determined by the demographic value of the victims. Ecol Evol 11:7164–7186. https://doi.org/10.1002/ece3.7574
Watabe R, Saito MU (2021a) Diel activity patterns of three sympatric medium-sized carnivores during winter and spring in a heavy snowfall area in northeastern Japan. Mamm Study 46:69–75. https://doi.org/10.3106/ms2020-0039
Watabe R, Saito MU (2021b) Effects of vehicle-passing frequency on forest roads on the activity patterns of carnivores. Landsc Ecol Eng 17:225–231. https://doi.org/10.1007/s11355-020-00434-7
Watabe R, Saito MU (2022) Diel activity pattern of a nonindigenous species, the masked palm civet in the Shonai region, Yamagata Prefecture, Japan. Tohoku J for Sci 27:11–17 (in Japanese)
Watabe R, Saito MU, Enari HS, Enari H (2020) Mammalian fauna of the Kaminagawa experimental forest of Yamagata University detected by camera traps. Tohoku J for Sci 25:37–40 (in Japanese)
Watabe R, Tsunoda H, Saito MU (2022) Evaluating the temporal and spatio-temporal niche partitioning between carnivores by different analytical method in northeastern Japan. Sci Rep 12:11987
Willebrand T, Willebrand S, Jahren T, Marcström V (2017) Snow tracking reveals different foraging patterns of red foxes and pine martens. Mammal Res 62:331–340. https://doi.org/10.1007/s13364-017-0332-2
World Meteorological Organization (2020) WMO provisional report on the state of the global climate 2020, https://reliefweb.int/report/world/wmo-provisional-report-state-global-climate-2020
Yamamoto Y (1994) Comparative analyses on food habits of Japanese marten, red fox, badger and raccoon dog in the Mt. Nyugasa, Nagano Prefecture. Japan Nat Environ Sci Res 7:45–52 (in Japanese with English summary)
Zalewska K, Waggershauser CN, Kortland K, Lambin X (2021) The best defence is not being there: avoidance of larger carnivores is not driven by risk intensity. J Zool 315:110–122. https://doi.org/10.1111/jzo.12910
Zalewski A (2000) Factors affecting the duration of activity by pine martens (Martes martes) in the Białowieża National Park, Poland. J Zool 251:439–447. https://doi.org/10.1017/S0952836900008037
Zalewski A (2001) Seasonal and sexual variation in diel activity rhythms of pine marten Martes martes in the Białowieża National Park (Poland). Acta Theriol 46:295–304. https://doi.org/10.1007/BF03192436
Zhou YB, Newman C, Xu WT, Buesching CD, Zalewski A, Kaneko Y, Macdonald DW, Xie ZQ (2011) Biogeographical variation in the diet of holarctic martens (genus Martes, Mammalia: Carnivora: Mustelidae): adaptive foraging in generalists. J Biogeogr 38:137–147. https://doi.org/10.1111/j.1365-2699.2010.02396.x
Zoller H, Drygala F (2013) Activity patterns of the invasive raccoon dog (Nyctereutes procyonoides) in North East Germany. Folia Zool 62:290–296
Acknowledgements
We thank the associates of the Yamagata University and the staff of Experimental Forest of the Yamagata University for supporting our field survey, and f the people in Kaminagawa and Nishiaraya communities in Tsuruoka City for permitting our field survey. We also thank Dr. H. Enari for providing helpful comments on this study and Dr. H. Tsunoda for providing helpful comments on data analysis and literature for this study. We are grateful to anonymous reviewers for improving an earlier draft of our manuscript.
Funding
This work was partly supported by YU-COE (M) grant from Yamagata University to MUS.
Author information
Authors and Affiliations
Contributions
RW and MUS conceived and designed the study, RW conducted the data collection and statistical analyses, and RW and MUS wrote the manuscript.
Corresponding author
Ethics declarations
Ethics approval
The procedures of this study were in accordance with the national laws of Japan. Ethical approval from ethics committee for involving animals was not required.
Competing interests
The authors declare no competing interests.
Additional information
Communicated by B. Voelkl
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Watabe, R., Saito, M.U. Winter weather conditions result in temporal niche overlap among three sympatric medium-sized carnivores in northeastern Japan. Behav Ecol Sociobiol 76, 164 (2022). https://doi.org/10.1007/s00265-022-03271-w
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-022-03271-w