
Abstract
Agonistic encounters among carnivores can be potentially avoided or minimized by shifts in daily activity patterns. Here, we investigated the temporal ecology of mesocarnivores which co-occur in a semiarid area where top predators are virtually absent. More specifically, we (i) describe the daily activity patterns of six mesocarnivore species, (ii) evaluate possible seasonal changes in their daily activity patterns, (iii) examine patterns of temporal overlap among mesocarnivores, and (iv) test the overlap in daily activity between mesocarnivores and their potential prey. Using camera-trapping data (13,976 camera-days) and circular and overlapping analyses, we studied six out of the seven mesocarnivore species recorded. Striped hog-nosed skunk and the crab-eating raccoon were nocturnal, crab-eating fox and northern tiger cat were nocturnal-crepuscular, ocelot was mainly nocturnal, and jaguarundi was diurnal. With the exception of jaguarundi, we fail to find strong temporal segregation among mesocarnivore and interspecific interactions did not vary seasonally, but we observed separation in their activity peaks and significant difference in their activity distributions. This partial temporal segregation can potentially contribute to interspecific coexistence, reducing the chances of interspecific killing, mainly in relation to the dominant species (ocelot). Mesocarnivores did not exhibit a significant synchrony of their activity with any of the preys evaluated, with the exception of jaguarundi, which significantly overlapped its distribution of activity with some preys. Temporal segregation contributes, but does not seem to be the only mechanism behind the coexistence of mesocarnivores in Caatinga dry forest; thus, other strategies such as spatial and dietary segregation should be considered.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Patterns of daily activity are a crucial component of mammalian ecology and behavior, through which species respond to variations in biotic, abiotic, and anthropogenic factors (Schoener 1974; Bennie et al. 2014; Gaynor et al. 2018). Many species avoid being active in periods of intense heat or cold to avoid hypo- or hyperthermia (Terrien et al. 2011), or during full moon nights, which can make predators and prey more exposed to mutual detection (Prugh and Golden 2014). Predators can adjust their daily activity in order to maximize the chances of prey encounter thus minimizing the energy expenditure, while pressing the prey to avoid encounters with potential predators (Foster et al. 2013; Monterroso et al. 2013). In addition, the negative effects of interspecific competition can also be minimized through temporal segregation, which is an important mechanism of coexistence among species with similar ecology and morphology (Lucherini et al. 2009; Di Bitetti et al. 2010; Monterroso et al. 2014). This strategy contributes to the avoidance of agonistic encounters which minimize interference competition levels and reduce the chances of intraguild predation and interspecific killing among mammalian carnivores (Carothers and Jaksic 1984; Polis et al. 1989; Palomares and Caro 1999).
Mesocarnivores (< 15 kg) usually occupy a trophic position just below the top predators (Roemer et al. 2009; Ritchie and Johnson 2009), being submitted to top-down control by apex predators while competing for habitat and food resources (Ritchie and Johnson 2009). So, the reduction in numbers or even the absence of top predators can contribute to increase the abundance of mesocarnivores, which can expand their ecological niche intensifying predation and intraguild competition effects (Crooks and Soulé 1999; Prugh et al. 2009). Under these circumstances, larger and generally dominant mesocarnivores (Oliveira and Pereira 2014) may emerge as apex predators affecting subordinate species (Prugh et al. 2009). However, there are only a few studies on the ecology of sympatric neotropical mesocarnivores in areas where top predators were locally extinct, which usually correspond to unprotected areas (Woodroffe and Ginsberg 1998). This type of knowledge is essential to the conservation of biological communities (Bu et al. 2016; Wang et al. 2015) in a world with increasingly anthropogenic disturbances.
The Caatinga, a seasonally dry tropical forest in northeastern Brazil, is a naturally stressful environment which has been suffering high anthropogenic impacts (Silva et al. 2017; Antongiovanni et al. 2018) that can be expected to modulate species ecology (Stoner and Timm 2011; Astete et al. 2017a, 2017b). In most of the Caatinga, top predators such as the jaguar Panthera onca Linnaeus, 1758 and the puma Puma concolor Linnaeus, 1771 are declining or absent (Azevedo et al. 2013; Feijó and Langguth 2013; Morato et al. 2013; Marinho et al. 2018a). In such sites, mesocarnivores coexist while being released from top-down regulation. In Caatinga, the mesocarnivore assemblage is most commonly formed by seven more widely distributed species (Feijó and Langguth 2013; Marinho et al. 2018a): three omnivorous species, represented by crab-eating raccoon Procyon cancrivorus (G.[Baron] Cuvier, 1798) (adult body mass = 5.4–8.8 kg), crab-eating fox Cerdocyon thous Linnaeus, 1766 (5.7–6.5 kg), and striped hog-nosed skunk Conepatus amazonicus Lichtenstein, 1838 (2.4 kg), and four species that are strictly carnivorous, represented by the felids ocelot Leopardus pardalis Linnaeus, 1758 (8–11 kg), jaguarundi Herpailurus yagouaroundi É, Geoffroy Saint-Hilare, 1803 (3–6 kg), and northern tiger cat Leopardus tigrinus Thomas, 1904 (1.5–3 kg), along with the mustelid lesser grison Galictis cuja Molina, 1782 (1–3 kg) (Paglia et al. 2012; Oliveira and Pereira 2014). This overlap in terms of ecological niche, associated with the virtual absence of top predators, can result in the increase of competition effects, which, in turn, can be lowered by deviations in the use of trophic, spatial, and temporal resources (Schoener 1974). For the symtopic mesocarnivores, especially with analogous morphology and hunting strategies, alterations in activity patterns can improve coexistence. In Brazilian semiarid regions, studies regarding mesocarnivore activity patterns and temporal interactions are focused on a single species or in a sub-set of the mesocarnivore guild, while they usually do not consider the seasonal effect (Dias and Bocchiglieri 2016; Dias 2017; Penido et al. 2017; Dias et al. 2018, 2019; Marinho et al. 2018b).
In this study, we use camera-trapping data to examine the temporal ecology of a syntopic mesocarnivore guild in a Caatinga dry forest area virtually free of top predators. Puma seems absent from the area for over 10 years (Marinho et al. 2018a), while no mention is provided on current or previous presence of the jaguar, the other Caatinga top predator. Our objectives were (i) to describe the daily activity patterns of the mesocarnivore species, (ii) to evaluate possible seasonal changes in their daily activity patterns, (iii) to examine patterns of segregation or temporal overlap among mesocarnivores, and (iv) to test the overlap in daily activity between mesocarnivores and their potential prey. According to previous information, we expected mesocarnivores to be nocturnal or mainly nocturnal (Dias 2017; Penido et al. 2017; Marinho et al. 2018b; Dias et al. 2018, 2019), except for jaguarundi, which has a well-documented diurnal habit throughout its distribution (Giordano 2016).
We expected that mesocarnivores with closely related ecology and morphology, particularly the felids, would exhibit greater temporal segregation, at least during their peaks of activity (Penido et al. 2017; Dias et al. 2019), to avoid aggression risk and competition for resources (exploitative competition), while omnivorous species would differentiate their activity mainly in relation to the ocelot, the largest mesocarnivore known in the area. An alternative hypothesis is that the activity patterns of the mesocarnivores are more synchronized with their preferential prey, especially to the hypercarnivorous felids (Linkie and Ridout 2011; Foster et al. 2013; Porfirio et al. 2016). Considering that the Caatinga dry forest is a highly seasonal environment, we hypothesized that temporal niche segregation would be larger during the dry season, when water and food resources are more scarce and concentrated and the visual contact among the mesocarnivores is potentially more common due to the lower understory leaf density, which may increase interference competition (Valeix et al. 2007; Vanak et al. 2013). In general, we expected a possible lower daytime activity in the dry season related to the avoidance of high daytime temperatures (Pita et al. 2011), considering the low humidity and the poor protection of vegetation cover in this period.
Methods
Study site
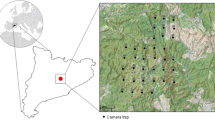
This study was developed in a seasonally dry tropical forest (Caatinga) area between the Feiticeiro and Bonfim mountains in Lajes municipality, Rio Grande do Norte state, northeastern Brazil (5° 44’–5° 51’ S latitude, 36° 11’–36° 06’ W longitude, Fig. 1). The region is considered a priority for the conservation of the Caatinga domain (MMA 2016; Fonseca et al. 2017). The climate is semiarid, with high temperatures and low rainfall. The higher precipitations usually occur between March and May, with average annual rainfall varying from 400 to 650 mm. The vegetation varies from open shrub formations, generally more disturbed and occurring at lower altitudes, to more forested and dense patches, mainly covering higher and steeper areas (Velloso et al. 2002).

Study area and distribution of the 50 camera-trap stations (A) used to investigate the temporal niche and coexistence of a mesocarnivore guild in a Caatinga dry forest in Lajes municipality, Rio Grande do Norte state (B), northeastern Brazil (C).
Despite being the largest continuous remnant of Caatinga vegetation in the Rio Grande do Norte state (Projeto Caatinga Potiguar 2015), the area has no legal protection and is under several anthropogenic disturbances developed in private properties, such as extensive livestock, cutting wood for charcoal production, and conversion of natural habitats to temporary agriculture. Poaching is another intense activity in the area (Marinho et al. 2018a). The last reliable reports of presence of puma in the region are more than 10 years old (Marinho et al. 2018a), being probably locally extinct (at least functionally) due to hunting and conflicts with cattle ranchers. Finally, the region is the target of mineral exploration, and is being prospected for installation of wind power plants.
Sampling design
Mesocarnivore occurrences were recorded through camera trapping in an area of approximately 70 km2, between May 2016 and February 2019, totaling a sampling effort of 13,976 camera-days divided into three sampling periods and along a total of 50 sampling points (Table S1). In the first survey (24 stations from May to June 2016), we used stations with double cameras aiming to estimate the density of the wild cats, while in the remaining two surveys we used stations with a single camera (43 stations from January to October 2017 and 37 stations from October 2018 to February 2019, Table S1). The sampling effort was divided in wet (7337 camera-days, January–June) and dry season (6639 camera-days, July–December) (adapted from Tomasella et al. 2018).
We used camera traps with heat and motion sensor (model Bushnell® Trophy Cam ™ HD). For the installation of the cameras, we prioritized trails used by people and/or livestock, dirt roads, and temporary streams. The distance between cameras was in average 929.5 m (SD = 262.1). We set the camera traps to record the date and time of the detections, with a minimum interval of 5 min between consecutive records, and we programmed them to take three photos per shot or one photo and one video of 10 s, remaining active 24 h every day. We considered the records of the same species at the same sampling point with more than 1 h apart as independent (Goulart et al. 2009). We did not use any bait to attract the animals.
Potential prey
The diet of mesocarnivores in the Caatinga is poorly known, but few existing studies can give an idea of their potential prey (Ximenez 1982; Olmos 1993; Dias and Bocchiglieri 2015, 2016). The striped hog-nosed skunk feeds mainly on arthropods and lizards preyed on burrows, and occasionally on fruits and small mammals such as marsupials (Olmos 1993). The crab-eating fox diet in the region is based mainly on arthropods, but also on fruits, lizards, birds, and some small rodents such as Spix’s cavy Galea spixii (Wagler, 1831) (Olmos 1993; Dias and Bocchiglieri 2016). The crab-eating raccoon can consume mainly arthropods, followed by fruits, and small vertebrates such as lizards, birds, and small rodents (Dias and Bocchiglieri 2015). In the Serra da Capivara National Park, northern tiger cat fed on large number of lizards such as Ameiva ameiva (Linnaeus, 1758) and Tropidurus hispidus (Spix, 1825), as well as on arthropods, birds, and a smaller proportion of unidentified small rodents (Olmos 1993). Ximenez (1982) also reported the presence of lizard scales in the stomach contents of the northern tiger cat. With respect to jaguarundi diet in Caatinga, there are reports of predation of common marmoset Callithrix jacchus (Linnaeus, 1758) (Ximenez 1982), rock cavy Kerodon rupestris F. Cuvier, 1825, Spix’s cavy, punaré rat Thrichomys laurentius Thomas, 1904, and other small rodents, as well as, birds, lizards, and arthropods (Olmos 1993; Dias and Bocchiglieri 2015). Finally, birds and lizards were more present in the diet of ocelot in Caatinga, although punaré rat and white-eared opossum Didelphis albiventris (Lund, 1840) also had been predated (Dias and Bocchiglieri 2015). However, due to its larger body size, the ocelot can also feed on larger prey such as armadillos (Wang 2002; Moreno et al. 2006).
Data analysis
We divided the independent records of each species in 1-h intervals over the 24 h circadian cycle. The uniformity of species records throughout the circadian cycle was tested using the Rayleigh test in the Oriana v.4 program (Kovach Commuting Services, Wales, UK). In order to describe the activity patterns, we classified the records in diurnal (between 1 h after sunrise and 1 h before sunset), nocturnal (between 1 h after sunset and 1 h before sunrise), and crepuscular (± 1 h of sunrise or sunset) (Porfirio et al. 2016). To define the exact time of sunrise and sunset, we used the software Tropsolar 5.0 (Cabús 2015). Since the samplings were carried out throughout the year, we estimated the monthly variations in the sunrise and sunset times from the 15th day of each month to classify the species records. For sunrise, the average value was 5:20 h, with an annual variation from 04:57 h to 05:35 h, while for sunset, the average value was 17:18 h, with a variation of 17:09 h to 17:40 h throughout the year. For these analyses, we used legal time as a reference, since annual variations in the daytime period are lower at low latitudes (Nouvellet et al. 2012).
We tested the existence of temporal segregation between the activity patterns of species pairs, within the mesocarnivore guild and between mesocarnivores and their potential prey species, with the Mardia-Watson Wheeler test (MWW test) in the software Oriana v.4. For each species, the same method was performed to test for shifts in daily activity patterns between the dry and wet season. In addition, we used the non-parametric Kernel density function to estimate the activity overlap coefficient (Δ) between mesocarnivores, as well as with their prey and between seasons (Ridout and Linkie 2009; Linkie and Ridout 2011). This coefficient, defined as the area under the curves formed by the two density functions in each time unit, ranges from 0 (no overlap) to 1 (total temporal overlap of activity) (Ridout and Linkie 2009). We obtained the 95% confidence intervals of the overlap coefficients estimated through 1000 bootstrap samples (Linkie and Ridout 2011; Meredith and Ridout 2018). According to Ridout and Linkie (2009), we used the coefficient Δ1 for small samples (< 75 records for at least one of the pairs compared) and the coefficient Δ4 for large samples (> 75 records). These analyses were done with the R package overlap (Meredith and Ridout 2018; R Development Core Team 2012). Finally, we classified the activity overlap between each comparison as follows: low overlap (Δ ≤ 0.5), moderate overlap (0.5 < Δ ≤ 0.75), and high overlap (Δ > 0.75) (Monterroso et al. 2014). We performed the analysis of activity overlap only with pairs of data (intra or between species) with more than 10 records in each pair evaluated.
Results
We recorded all seven species of mesocarnivores expected for the study area (Table 1). The number of records ranged from 1133 for the crab-eating fox to just five records for the lesser grison. Due to the low number of records of the lesser grison, this species was excluded from all posterior analyses.
The species activity patterns were not homogenous throughout the circadian cycle according to Rayleigh test (Fig. 2; Table 1). The striped hog-nosed skunk was nocturnal, starting its activities during dusk but avoided the dawn (Fig. 2a). The crab-eating fox was nocturnal-crepuscular, this canid started its activity at dusk, remained active throughout the night and reached the peak of activity at dawn, with some residual activity during the first part of the day (Fig. 2b). The crab-eating raccoon was nocturnal, but had a peak activity during dusk and another just before dawn (Fig. 2c). The ocelot was mostly nocturnal, but it initiated strongly its activities during dusk and slowed down continuously until the first hours of the morning (Fig. 2d). The northern tiger cat was nocturnal-crepuscular, but it was relatively generalist, performing a fair amount of its activities during day hours (Fig. 2e). Finally, the jaguarundi was diurnal, but it also presented a high activity during the dawn and dusk periods (Fig. 2f).

Daily activity patterns of mesocarnivores in a Caatinga dry forest, northeastern Brazil. Each circular histogram is divided into 24 intervals of 60 min, and their bars represent the percentage of the total number of camera-trap detections in each interval. Day (on average, 06:20 h–16:18 h), night (after 18:18 h–04:20 h), and twilight (± 1 before and after 5:20 h and 17:18 h) correspond to white, black, and gray, respectively. Species are (a) striped hog-nosed skunk, (b) crab-eating fox, (c) crab-eating raccoon, (d) ocelot, (e) northern tiger cat, and (f) jaguarundi. Origin of the specie’s images: De Angelo et al. (2015)
Daily activity distribution patterns were very similar between dry and wet periods for all mesocarnivore species; differences in activity distributions between seasons being non-significant (Fig. 3; Table 1). The crab-eating fox (Δ = 0.94 [0.90–0.97]) showed the highest activity overlap between seasons, while jaguarundi exhibited the lowest (Δ = 0.78 [0.63–0.90]), with some decrease in its sunset peak of activity during dawn in dry season (Fig. 3; Table 1).

Density estimates of daily activity patterns and extension of overlap within mesocarnivore species between dry and wet season in a Caatinga dry forest, northeastern Brazil. Overlap is represented by the shaded gray area. The dashed vertical lines represent the average legal time of sunrise [5:20 h] and sunset [17:18 h]) during the study period. The time of the records is shown as ticks in the bottom of the figures
Intraguild interactions
Overlap of daily activity patterns among mesocarnivore species varied widely, from 0.14 to 0.92 (Fig. 4). The activity distribution test indicated that there were significant differences in the activity patterns for 10 of the 15 mesocarnivore contrasts (Table 2). The lower overlap coefficients (0.14–0.48) appeared between the diurnal jaguarundi and all other species, showing a strong and significant temporal niche differentiation (Fig. 4; Table 2). The higher overlap coefficients occurred between crab-eating raccoon and all other mesocarnivores (0.76–0.83), except jaguarundi, and between crab-eating fox and northern tiger cat (0.92) (Fig. 4; Table 2), which indicate temporal niche similarity. The remaining species contrasts had moderate to high overlap coefficient (0.70–0.79), but always showing significant temporal niche differences through their activity distributions (Fig. 4; Table 2). It should be highlighted that we found no evidence of change in intraguild interactions between dry and wet seasons (Table S2).

Density estimates of daily activity patterns and extension of their overlap among pairs of mesocarnivores in a Caatinga dry forest, northeastern Brazil. Overlap is represented by the shaded gray area. The dashed vertical lines represent the average legal time of sunrise [5:20 h] and sunset [17:18 h]) during the study period. Significant differences in MWW test are indicated by an asterisk after overlap coefficient values (∆). The time of the records is shown as ticks in the bottom of the figures. Origin of the specie’s images: De Angelo et al. (2015)
Predator-prey temporal overlap
Camera trapping detected several mammal (N = 1184 records), bird (N = 1031), and lizard (N = 179) species that could be potential preys of mesocarnivores (Table S3). Among the mammals, we recorded Spix’s cavy, punaré rat, rock cavy, white-eared opossum, yellow armadillo Euphractus sexcinctus (Linnaeus, 1758), and nine-banded armadillo Dasypus novemcinctus (Linnaeus, 1758). In relation to the ground-dwelling and ground-foraging birds, we recorded several species of doves, including Columbina spp., Zenaida auriculata(Des Murs, 1847), and Leptotila verreauxi (Bonaparte, 1855), as well as tinamou species of the genera Crypturellus and Nothura. Among the lizards, we recorded A. ameiva, T. hispidus, Tropidurus semitaeniatus (Spix, 1825), and Ameivula ocellifera (Spix, 1825) (Table S3). Due to identification uncertainties, in the following analyses, doves, tinamous, and lizards were analyzed as groups (Table S3).
Most potential mammalian preys exhibited nocturnal or mainly nocturnal activity, with the exception of rock cavy and yellow armadillo which were cathemeral and crepuscular, respectively (Table S3; Fig. S1). For birds, doves were diurnal while tinamous were crepuscular (Table S3; Fig. S1). Finally, lizards were diurnal (Table S3; Fig. S1). Most preys had similar daily activity patterns throughout the year, with the exception of yellow armadillo and the doves, which exhibited significant difference between the dry and wet season (Table S3).
Among the omnivorous mesocarnivores (Table 3; Fig. S1), the daily distributions of records of the striped hog-nosed skunk differed significantly from the pattern exhibited by most potential preys, except from punaré rat and white-eared opossum (Table 3; Fig. S1). The distribution of activity of the crab-eating fox was significantly different from all potential preys, although it had a reasonable temporal overlap with some small mammals (Table 3; Fig. S1). Finally, the activity pattern of crab-eating raccoon was similar to Spix’s cavy and punaré rat (Table 3; Fig. S1), and to the white-eared opossum during dry season (Table S4), but differed from the other potential preys.
Regarding the felids (Table 3; Fig. S1), the ocelot and the northern tiger cat differed significantly their activity distributions from all potential preys, although they had a reasonably high activity overlap with some small mammal species such as Spix’s cavy and white-eared opossum (Table 3; Fig. S1). In fact, in the wet season, the distribution of activity of the ocelot did not differ from the white-eared opossum (Table S4). Finally, the activity pattern of jaguarundi was similar to that exhibited by rock cavy and tinamous in both seasons (Table S4), but differed from other potential prey (Table 3; Fig. S1). Except to the ocelot, predator-prey temporal interactions were extremely conserved across seasons (Table S4).
Discussion
This study provides a broad description of the temporal ecology of a complete guild of mesocarnivores, including intraguild and predator-prey temporal interactions, in an area of the Brazilian Caatinga where top predators are locally extinct or functionally absent. The observed activity patterns are qualitatively similar to those described in the literature: striped hog-nosed skunk is classified as nocturnal (Cavalcanti et al. 2014; Dias 2017), crab-eating fox as nocturnal-crepuscular (Bianchi et al. 2016; Dias and Bocchiglieri 2016; Penido et al. 2017), crab-eating raccoon as nocturnal (Gómez et al. 2005; Bianchi et al. 2016), ocelot as mainly nocturnal (Di Bitetti et al. 2010; Oliveira-Santos et al. 2012; Massara et al. 2016; Penido et al. 2017; Dias et al. 2018, 2019; Nagy-Reis et al. 2019), northern tiger cat as nocturnal-crepuscular (Penido et al. 2017; Marinho et al. 2018b; Dias et al. 2019), and jaguarundi as diurnal (Giordano 2016; Massara et al. 2016; Dias et al. 2019). In the case of the lesser grison, although the low number of records prevented any analysis, all five records were obtained between 06:15 and 07:15 h, suggesting a daytime activity as already reported in the literature (Kasper et al. 2013).
Potential competitor species tend to develop mechanisms to alleviate competition, especially in the case of morphologically similar and closely related species (Schoener 1974). In the case of carnivorous mammals, besides the exploitation competition, the risk of aggression (interference competition) and intraguild predation can induce submissive species to be active at hours with a lower probability of finding a dominant competitor (Polis et al. 1989). Thus, temporal avoidance is often the most important mechanism of coexistence (Bianchi et al. 2016; Carothers and Jaksic 1984). Our results provide partial support for the hypothesis that temporal segregation represents a mechanism that facilitates the coexistence of mesocarnivores in a semiarid region. Although most pairs of species exhibited a high or moderate activity overlap, almost all mesocarnivores segregated at least their activity peaks throughout the circadian cycle, suggesting a partial avoidance that may decrease competition as well as the risk of intraguild predation (Carothers and Jaksic 1984), especially in relation to the larger species in the area (i.e., the ocelot). For example, the ocelot has a relatively continuous higher activity between 18:00 p.m. and 2:00 a.m., followed by a decrease until approximately 5:00 a.m., while the other species exhibited a higher proportion of activity in the second part of the night, with two of them (crab-eating fox and northern tiger cat) displaying a larger peak near sunrise hours similar to diurnal jaguarundi. The studies have pointed out a high activity overlap between ocelot and omnivorous mesocarnivore species (Bianchi et al. 2016; Massara et al. 2016), and a low to moderate temporal overlap between ocelot and small sympatric felid species (Di Bitetti et al. 2010; Massara et al. 2016; Nagy-Reis et al. 2019).
Regarding generalist or omnivorous species, we found some evidence of temporal segregation between the striped hog-nosed skunk and the crab-eating fox, but no segregation between the crab-eating raccoon and the other two species. In the case of the crab-eating raccoon, its larger body size in relation to the other mesocarnivores prevents it from being attacked or predated (Oliveira and Pereira 2014). In addition, its preference for environments near water bodies (Cheida et al. 2013), as well as the consumption of aquatic and semi-aquatic prey, may decrease competition with other mesocarnivores through spatial and dietary segregation, respectively, at least when there are enough water bodies. However, considering the scarcity of water bodies in the Caatinga, especially in the dry season, it is possible that during most of the year, there is a considerable trophic niche overlap among all omnivorous mesocarnivores. In an environment with greater resource availability such as the Pantanal, temporal segregation was more important for generalist mesocarnivores, including crab-eating fox and crab-eating raccoon (Bianchi et al. 2016). Further studies are necessary for a better understanding of the ecology of the crab-eating raccoon in a semiarid and seasonal environment like the Brazilian Caatinga.
The three felid species, which due to their ecological similarities can be strong competitors, partially segregated their temporal activity. While the diurnal jaguarundi exhibited a daily activity pattern very different from the other two felids, the ocelot and the northern tiger cat separated their peaks of higher activity despite overlapping much of their daily activity. This pattern is similar to that found in other areas of the Caatinga where larger predators occur (Penido et al. 2017; Dias et al. 2019), but it is somewhat different from the pattern reported for a very closely related species, the southern tiger cat Leopardus gutullus Hensel, 1872 (Massara et al. 2016; Nagy-reis et al. 2019). In the mesic Atlantic Forest of southern and southeastern Brazil, the southern tiger cat exhibited a nocturnal behavior in the absence of the ocelot and the puma (Oliveira-Santos et al. 2012) compared to a cathemeral or mostly diurnal activity where larger felids are present with the overlap between them ranging from moderate to low (Oliveira-Santos et al. 2012; Massara et al. 2016; Nagy-Reis et al. 2019). Unlike what happens in these more humid environments, where temporal segregation seems to be stronger (Massara et al. 2016; Nagy-Reis et al. 2019), in the Brazilian semiarid, differentiation in spatial dimensions may be important to promote mesocarnivore coexistence (Dias et al. 2019). In this sense, future studies should investigate whether spatial segregation is acting in main or complementary way toward coexistence of this carnivore guild.
We found a high overlap in temporal activity patterns between northern tiger cat and crab-eating fox, which contrasts with the temporal segregation found in an area of Caatinga where large felids coexist with such mesocarnivores (Penido et al. 2017). Their coexistence can be facilitated by the fact that these two species partially differ in their diet, with crab-eating fox being omnivorous and northern tiger cat strictly carnivorous. The crab-eating fox, for instance, can be found near human habitations looking for food resources, while the northern tiger cat normally avoids such areas (Marinho et al. 2018b). These observations also suggest that an alternative mechanism of coexistence can be in the space use. Spatial segregation could alleviate possible agonistic interactions, despite the chances for intraguild predation are considered to be low (Oliveira and Pereira 2014). It should be notice that the temporal overlap between these two mesocarnivores may be forced by the avoidance of the peak activity of ocelot and hottest part of the day.
In addition to interspecific interactions, the environmental conditions may also be critical in shaping the species activity. In semiarid environments such as the Caatinga, where there are high solar radiation and high diurnal temperatures, both predators and prey are expected to be active during nocturnal and twilight periods to avoid overheating and water loss (Terrien et al. 2011; Penido et al. 2017). Thus, this should limit the temporal window within the circadian cycle available for species to adjust their activity in response to competition or predation risk. The considerable overlap in activity of even ecologically close species such as the ocelot and the northern tiger cat suggests this limitation. Thus, the differentiation in activity peaks may be a more efficient mechanism of coexistence.
In contrast to what one could expect, we did not detect important variations in the daily activity patterns of mesocarnivores between dry and wet periods. Since the Brazilian Caatinga experiences a marked seasonal variation in precipitation and resource availability along the year (Andrade et al. 2017), our results indicate that mesocarnivores are well adapted to such variable conditions. Possible behavioral mechanisms explain such resilience include diet and spatial shifts. For instance, during months of greater heat and water stress, the animals may be moving to more mesic locations such as mountains or to sites with water sources (Carmignotto and Astúa 2017).
Predators tend to manage the pressure for intraguild segregation, but they also need to synchronize their activity with their main prey to reduce energy expenditure on food hunting (Foster et al. 2013; Monterroso et al. 2013). According to our results, for most mesocarnivores, the activity pattern seems to reflect a balance between partial temporal segregation with competitors/predators and partial temporal overlap with as many potential prey as possible, since few species such as jaguarundi synchronized their activity with specific potential prey. All of the mesocarnivore species have a high or moderate activity overlap with at least one type of prey, although the peaks of activity were generally different. This may allow a sequential exploration of the resources while decreasing the chances of agnostic encounters (Monterroso et al. 2013). Indeed, the other mesocarnivores that exhibited high overlap and synchrony in their activity with potential prey are the omnivorous striped hog-nosed skunk and crab-eating raccoon, which should only occasionally feed on small mammals (Olmos 1993; Dias and Bochiglieri 2015).
In the case of the jaguarundi, the significant overlap in the distribution of activity with the rocky cavy and tinamous suggests that these must be important preys in the region, while the other wild felid species should exploit these preys in a complementary way. In other Caatinga areas, the rocky cavy has a more nocturnal activity which is more overlapped by the activity of the northern tiger cat (Penido et al. 2017; Dias et al. 2019) and the ocelot (Dias et al. 2018) than by the jaguarundi. Dias et al. (2019) also found a high and significant overlap of the ocelot activity with the nine-banded armadillo, a species apparently rare in our area, while the yellow armadillo is abundant but has a markedly distinct activity compared to the ocelot.
However, it is important to highlight that the temporal overlap alone does not define the vulnerability of the prey or the preference of the predator, since there must also be spatial overlap, not investigated here. In addition, some prey can be captured while resting or taking refuge in their dens (Emsens et al. 2013), such as suggested to crab-eating fox and hog-nosed skunk in Caatinga, which seem to prey on lizards removing them from their dens (Olmos 1993). Therefore, studies on spatial interactions and diet of mesocarnivores in this semiarid region are important to elucidate predator-prey relationships and the level of feeding overlap between mesocarnivores.
Finally, our results suggest that the mesocarnivore species have generally a higher proportion of nighttime activity, and that there is a separation in their activity peaks rather than a stronger temporal segregation which remains throughout the seasons, possibly as a trade-off between avoiding the aggressive encounters with competitors/predators and hotter periods of the circadian cycle. Although mesocarnivores presented a high overlap with at least one prey, species such as the crab-eating fox, the northern tiger cat, and the ocelot did not exhibit a strong synchrony of their activity with any of the preys evaluated, suggesting a more generalist behavior that could contribute to mediate intraguild interactions. Our results contribute to the understanding of the ecology of mesocarnivores, intraguild interactions, and predator-prey relationships in semiarid environments in a scenario of potential disturbance caused by the eradication of top predators. It is important to keep in mind that high overlap of daily activity between species does not necessarily determine a high potential of encounter if these species segregate spatially; so, further studies should seek to understand the role of the spatial and trophic dimensions of the ecological niche in intraguild interactions, and ideally to compare the ecology of species in areas with different degrees of carnivore guild integrity.
References
Andrade EM, Aquino DN, Chaves LCG, Lopes FB (2017) Water as capital and its uses in the Caatinga. In: Silva JC, Leal I, Tabarelli M (eds) Caatinga: the largest tropical dry forest region in South America. Springer, Cham, pp 281–302
Antongiovanni M, Venticinque EM, Fonseca CR (2018) Fragmentation patterns of the Caatinga drylands. Landsc Ecol 33:1353–1367. https://doi.org/10.1007/s10980-018-0672-6
Astete S, Marinho-Filho J, Kajin M, Penido G, Zimbres B, Sollmann R, Jácomo ATA, Tôrres NM, Silveira L (2017a) Forced neighbours: coexistence between jaguars and pumas in a harsh environment. J Arid Environ 146:27–34. https://doi.org/10.1016/j.jaridenv.2017.07.005
Astete S, Marinho-Filho J, Machado RB, Zimbres B, Jácomo ATA, Sollmann R, Tôrres NM, Silveira L (2017b) Living in extreme environments: modeling habitat suitability for jaguars, pumas, and their prey in a semiarid habitat. J Mammal 98:464–474. https://doi.org/10.1093/jmammal/gyw184
Azevedo FC, Lemos FG, Almeida LB, Campos CB, Beisiegel BDM, Paula RC, Crawshaw Junior PG, Ferraz KMP, Barros M, De Oliveira TG (2013) Avaliação do risco de extinção da onça-parda Puma concolor (Linnaeus, 1771) no Brasil. Biodiv Brasil 3:107–121
Bennie JJ, Duffy JP, Inger R, Gaston KJ (2014) Biogeography of time partitioning in mammals. Proc Natl A Sci 111:13727–13732. https://doi.org/10.1073/pnas.1216063110
Bianchi RC, Olifiers N, Gompper ME, Mourão G (2016) Niche partitioning among pesocarnivores in a Brazilian Wetland. PLoS One 11:e0162893. https://doi.org/10.1371/journal.pone.0162893
Bu H, Wang F, McShea WJ, Lu Z, Wang D, Li S (2016) Spatial co-occurrence and activity patterns of mesocarnivores in the temperate forests of Southwest China. PLoS One 11:e0164271. https://doi.org/10.1371/journal.pone.0164271
Cabús R (2015) Tropsolar 5.0. Grupo de Pesquisa em Iluminação (GRILU). http://www.ctec.ufal.br/grupopesquisa/gril. Accessed 15 April 2019
Carmignotto AP, Astúa D (2017) Mammals of the Caatinga: diversity, ecology, biogeography, and conservation. In: Silva JC, Leal I, Tabarelli M (eds) Caatinga: the largest tropical dry forest region in South America. Springer, Cham, pp 211–254
Carothers JH, Jaksic FM (1984) Time as a niche difference: the role of interference competition. Oikos 42:403–406. https://doi.org/10.2307/3544413
Cavalcanti GN, Alfaro-Alvarado LD, Rodrigues FHG (2014) Home range and activity of Conepatus semistriatus (Carnivora, Mephitidae) in Emas National Park, Brazil. Anim Biol 64:151–162. https://doi.org/10.1163/15707563-00002436
Cheida CC, Rodrigues FHG, Beisiegel BM (2013) Avaliação do risco de extinção do Guaxinim Procyon cancrivorus (Cuvier, 1798) no Brasil. Biodiv Brasil 3:283–290
Crooks KR, Soulé ME (1999) Mesopredator release and avifaunal extinctions in a fragmented system. Nature 400:563–566. https://doi.org/10.1038/23028
De Angelo C, Paviolo A, Di Blanco Y, Di Bitetti M, Chiappe A (2015) Guía de huellas de los mamíferos de Misiones y otras áreas del subtrópico de Argentina. Ediciones del Subtrópico, Tucumán
Di Bitetti MS, De Angelo CD, Di Blanco YE, Paviolo A (2010) Niche partitioning and species coexistence in a Neotropical felid assemblage. Acta Oecol 36:403e412. https://doi.org/10.1016/j.actao.2010.04.001
Dias DM, Bocchiglieri A (2015) Dieta de carnívoros (Mammalia, Carnivora) em um remanescente de Caatinga, Nordeste do Brasil. Bioikos 29:13–19
Dias DM, Bocchiglieri A (2016) Trophic and spatio-temporal niche of the crab-eating fox, Cerdocyon thous (Linnaeus, 1766) (Carnivora: Canidae), in a remnant of the Caatinga in northeastern Brazil. Mammalia 80:281–291. https://doi.org/10.1515/mammalia-2014-0108
Dias DM (2017) Spatiotemporal ecology of the striped hog-nosed skunk Conepatus semistriatus (Carnivora, Mephitidae) in a seasonally dry forest of northeastern Brazil. Anim Biol 2:119–131. https://doi.org/10.1163/15707563-00002525
Dias DM, Campos CB, Rodrigues FHG (2018) Behavioural ecology in a predator-prey system. Mamm Biol 92:30–36. https://doi.org/10.1016/j.mambio.2018.04.005
Dias DM, Massara RL, Campos CB, Rodrigues FHG (2019) Feline predator-prey relationships in a semi-arid biome in Brazil. J Zool 307:282–291. https://doi.org/10.1111/jzo.12647
Emsens WJ, Hirsch BT, Kays R, Jansen PA (2013) Prey refuges as predator hotspots: ocelot (Leopardus pardalis) attraction to agouti (Dasyprocta punctata) dens. Acta Theriol 59:257–262. https://doi.org/10.1007/s13364-013-0159-4
Feijó A, Langguth A (2013) Mamíferos de médio e grande porte do Nordeste do Brasil: distribuição e taxonomia, com descrição de novas espécies. Rev Nord Biol 22:3–225
Fonseca CR, Antongiovanni M, Matsumoto M, Bernard E, Venticinque EM (2017) Conservation opportunities in the Caatinga. In: Silva JC, Leal I, Tabarelli M (eds) Caatinga: the largest tropical dry forest region in South America. Springer, Cham, pp 429–443
Foster VC, Sarmento P, Sollmann R, Tôrres N, Jácomo ATA, Negrões N, Fonseca C, Silveira L (2013) Jaguar and puma activity patterns and predator-prey interactions in four Brazilian biomes. Biotropica 45:373–379. https://doi.org/10.1111/btp.12021
Gaynor KM, Hojnowski CE, Cater NH, Bashares JS (2018) The influence of human disturbance on wildlife nocturnality. Science 360:1232–1123. https://doi.org/10.1126/science.aar7121
Giordano AJ (2016) Ecology and status of the jaguarundi Puma yagouaroundi: a synthesis of existing knowledge. Mammal Rev 46:30–43. https://doi.org/10.1111/mam.12051
Gómez H, Wallace RB, Ayala G, Tejada R (2005) Dry season activity periods of some Amazonian mammals. Stud Neotropical Fauna Environ 40:91–95. https://doi.org/10.1080/01650520500129638
Goulart FVB, Cáceres NC, Graipel ME, Tortato MA, Ghizoni IR Jr, Oliveiras-Santos LGR (2009) Habitat selection by large mammals in a southern Brazilian Atlantic Forest. Mamm Biol 74:182–190. https://doi.org/10.1016/j.mambio.2009.02.006
Kasper CB, Leuchtenberger C, Bornholdt R, Pontes ARM, Beisiegel BM (2013) Avaliação do risco de extinção do furão Galictis cuja (Molina, 1782) no Brasil. Biodiv Brasil 3:203–210
Linkie M, Ridout MS (2011) Assessing tiger-prey interactions in Sumatran rainforests. J Zool 284:224–229. https://doi.org/10.1111/j.1469-7998.2011.00801.x
Lucherini M, Reppucci JI, Walker RS, Villalba ML, Wurstten A, Gallardo G, Iriarte A, Villalobos R, Perovic P (2009) Activity pattern segregation of carnivores in the high Andes. J Mammal 90:1404–1409. https://doi.org/10.1644/09-MAMM-A-002R.1
Marinho PH, Bezerra D, Antongiovanni M, Fonseca CR, Venticinque EM (2018a) Mamíferos de médio e grande porte da Caatinga do Rio Grande do Norte, nordeste do Brasil. Mastozool Neotrop 25:345–362. https://doi.org/10.31687/saremMN.18.25.2.0.15
Marinho PH, Bezerra D, Antongiovanni M, Fonseca CR, Venticinque EM (2018b) Activity patterns of the threatened northern tiger cat Leopardus tigrinus and its potential prey in a Brazilian dry tropical forest. Mamm Biol 89:30–36. https://doi.org/10.1016/j.mambio.2017.12.004
Massara RL, Paschoal AMO, Bailey LL, Doherty PF Jr, Chiarello AG (2016) Ecological interactions between ocelots and sympatric mesocarnivores in protected areas of the Atlantic Forest, southeastern Brazil. J Mammal 97:1634–1644. https://doi.org/10.1093/jmammal/gyw129
Meredith M, Ridout M (2018) Overview of the overlap package. R Project. 1–9. https://cran.r project.org/web/packages/overlap/vignettes/overlap.pdf. Accessed 04 February 2019
MMA (2016) Áreas Prioritárias para conservação, uso sustentável e repartição de benefícios da biodiversidade do bioma Caatinga – 2ª atualização - Portaria N°223, de 21 de junho de 2016 Ministério do Meio Ambiente http://www.mma.gov.br/areas-protegidas/cadastro-nacional-de-ucs/item/10724-resultados-da-2%C2%B0-atualiza%C3%A7%C3%A3o-das-%C3%A1reas-priorit%C3%A1rias.html Accessed 20 December 2018
Monterroso P, Alves PC, Ferreras P (2013) Catch me if you can: diel activity patterns of mammalian prey and predators. Ethology 119:1044–1056. https://doi.org/10.1111/eth.12156
Monterroso P, Aves PC, Ferreras P (2014) Plasticity in circadian activity patterns of mesocarnivores daily activity coexistence. Behav Ecol Sociobiol 68:1403–1417. https://doi.org/10.1007/s00265-014-1748-1
Morato RG, Beisiegel BM, Ramalho EE, Campos CB, Boulhosa RLP (2013) Avaliação do risco de extinção da onça-pintada Panthera onca (Linnaeus, 1758) no Brasil. Biodiv Brasil 3:122–132
Moreno RS, Kays RW, Samudio R Jr (2006) Competitive release in diets of ocelot (Leopardus pardalis) and puma (Puma concolor) after jaguar (Panthera onca) decline. J Mammal 87:808–816. https://doi.org/10.1644/05-MAMM-A-360R2.1
Nagy-Reis MB, Iwakami VHS, Estevo CA, Setz EZF (2019) Temporal and dietary segregation in a Neotropical small-felid assemblage and its relation to prey activity. Mamm Biol 95:1–8. https://doi.org/10.1016/j.mambio.2018.12.005
Nouvellet P, Rasmussen GSA, MacDonald DW, Courchamp F (2012) Noisy clocks and silent sunrises: measurement methods of daily activity pattern. J Zool 286:179–184. https://doi.org/10.1111/j.1469-7998.2011.00864.x
Oliveira TG, Pereira JA (2014) Intraguild predation and interspecific killing as structuring forces of carnivoran communities in South America. J Mamm Evol 21:427–436. https://doi.org/10.1007/s10914-013-9251-4
Oliveira-Santos LGR, Graipel ME, Tortato MA, Zucco CA, Cáceres NC, Goulart FVB (2012) Abundance changes and activity flexibility of the northern tiger cat, Leopardus tigrinus (Carnivora: Felidae), appear to reflect avoidance of conflict. Zoologia 29:115–120. https://doi.org/10.1590/S1984-46702012000200003
Olmos F (1993) Notes on the food habitats of Brazilian “Caatinga” carnivores. Mammalia 57:126–130
Pita R, Mira A, Beja P (2011) Circadian activity rhythms in relation to season, sex and interspecific interactions in two Mediterranean voles. Anim Behav 81:1023–1030. https://doi.org/10.1016/j.anbehav.2011.02.007
Paglia AP, Fonseca GAB, Rylands AB, Herrmann G, Aguiar LMS, Chiarello AG, Leite YLR, Costa LP, Siciliano S, Kierulff MCM, Mendes SL, Tavares VC, Mittermeier RA, Patton JL (2012) Annotated checklist of Brazilian mammals. 2nd edition. Occas Pap Conserv Biol 6:1–76
Palomares F, Caro TM (1999) Interspecific killing among mammalian carnivores. Am Nat 153:492–508. https://doi.org/10.1086/303189
Penido G, Astete S, Jácomo ATA, Sollmann R, Tôrres N, Silveira L, Marinho Filho J (2017) Mesocarnivore activity patterns in the semiarid Caatinga: limited by the harsh environment or affected by interspecifc interactions? J Mammal 98:1732–1740. https://doi.org/10.1093/jmammal/gyx119
Polis GA, Myers CA, Holt RD (1989) The ecology and evolution of intraguild predation: potential competitors that eat each other. Annu Rev Ecol Syst 20:297–330. https://doi.org/10.1146/annurev.es.20.110189.001501
Porfirio G, Foster VC, Fonseca C, Sarmento P (2016) Activity patterns of ocelots and their potential prey in the Brazilian Pantanal. Mamm Biol 81:511–517. https://doi.org/10.1016/j.mambio.2016.06.006
Projeto Caatinga Potiguar (2015) Projeto Caatinga Potiguar – Cartograma. WCS-Brazil/UFRN. http://programs.wcs.org/brazil/Lugares-naturais/Projeto-Caatinga.aspx. Accessed 02 November 2017
Prugh LR, Golden CD (2014) Does moonlight increase predation risk? Meta-analysis reveals divergent responses of nocturnal mammals to lunar cycles. J Anim Ecol 83:504–514. https://doi.org/10.1111/1365-2656.12148
Prugh LR, Stoner CJ, Epps CW, Bean WT, Ripple WJ, Laliberte AS, Bashares JS (2009) The rise of the mesopredator. Bioscience 59:779–791. https://doi.org/10.1525/bio.2009.59.9.9
R Development Core Team (2012) R: a language and environment for statistical computing. R Foundation for Statistical Computing. http://www.R-project.org/ Accessed 30 April 2018
Ritchie EG, Johnson CN (2009) Predator interactions, mesopredator release and biodiversity conservation. Ecol Lett 12:982–998. https://doi.org/10.1111/j.1461-0248.2009.01347.x
Roemer GW, Gompper ME, Valkenburgh BV (2009) The ecological role of the mammalian mesocarnivore. BioScience 59:165–173. https://doi.org/10.1525/bio.2009.59.2.9
Ridout MS, Linkie M (2009) Estimating overlap of daily activity patterns from camera trap data. J Agr Biol Envir St 14:322–337. https://doi.org/10.1198/jabes.2009.08038
Schoener TW (1974) Resource partitioning in ecological communities. Science 185:27–39. https://doi.org/10.1126/science.185.4145.27
Silva JMC, Barbosa LCF, Leal I, Tabarelli M (2017) The Caatinga: understanding the challenges. In: Silva JC, Leal I, Tabarelli M (eds) Caatinga: the largest tropical dry forest region in South America. Springer, Cham, pp 3–19
Stoner K, Timm RM (2011) Seasonally dry tropical forest mammals: adaptations and seasonal patterns. In: Dirzo R, Young HS, Mooney HA, Ceballos G (eds) Seasonally dry tropical forests: ecology and conservation. Island Press, Washington, pp 85–106
Terrien J, Perret M, Aujard F (2011) Behavioral thermoregulation in mammals: a review. Front Biosci 16:1428–1444. https://doi.org/10.2741/3797
Tomasella J, Silva RMPV, Barbosa AA, Rodriguez DA, Oliveira MS, Sestini MF (2018) Desertification trends in the Northeast of Brazil over the period 2000–2016. Int J Appl Earth Obs 73:197–206. https://doi.org/10.1016/j.jag.2018.06.012
Vanak AT, Fortin D, Thaker M, Ogden M, Owen C, Greatwood S, Slotow R (2013) Moving to stay in place: behavioral mechanisms for coexistence of African large carnivores. Ecology 94:2619–2631. https://doi.org/10.1890/13-0217.1
Velloso AL, Sampaio EVSB, Pareyn FGC (2002) Ecorregiões propostas para o Bioma Caatinga. Associação Plantas do Nordeste, Instituto de Conservação Ambiental, The Nature Conservancy do Brasil, Recife
Valeix M, Chamaillé-Jammes S, Fritz H (2007) interference competition and temporal niche shifts – elephants and herbivore communities at waterholes. Oecologia 153:739–748. https://doi.org/10.1007/s00442-007-0764-5
Wang E (2002) Diets of Ocelots (Leopardus pardalis), Margays (L. wiedii), and Oncillas (L. tigrinus) in the Atlantic rainforest in Southeast Brazil. Stud Neotrop Fauna E 37:207–212. https://doi.org/10.1076/snfe.37.3.207.8564
Wang Y, Allen ML, Wilmers CC (2015) Mesopredator spatial and temporal responses to large predators and human development in the Santa Cruz Mountains of California. Biol Conserv 190:23–33. https://doi.org/10.1016/j.biocon.2015.05.007
Woodroffe R, Ginsberg JR (1998) Edge effects and the extinction of populations inside protected areas. Science 280:2126–2128. https://doi.org/10.1126/science.280.5372.2126
Ximenez A (1982) Notas sobre felidos neotropicales viii observaciones sobre el contenido estamacal y el comportamiento alimentar de diversas espécies de felinos. Rev Nord Biol 5:89–91
Acknowledgments
We are grateful to João B. de Lima (Seu João) and Joana Darque for the field assistance and hospitality, and to Eugenia C. Schmidt, Juan C. V. Mena, Felipe Marinho, Raul dos Santos, Maria L. Falcão, V. Paixão, and T. Oliveira for field or analysis assistance. We would like to thank two anonymous reviewers who helped to improve the work.
Funding
This study was partially supported by Restaurante Camarões and The Mohamed bin Zayed Species Conservation Fund (#172516360). PHM, PS, and CF would like to thank the University of Aveiro and FCT/MEC for the financial support to CESAM RU (UID/AMB/50017) through national funds and co-financed by the FEDER, within the PT2020 Partnership Agreement. EMV (308040/2017-1) and CRF (305304/2013-5; 306812/2017-7) were supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and PHM was supported by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES; financing code 001) and Santander Universities (Santander Mundi scholarship).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
(DOCX 769 kb)
Rights and permissions
About this article

Cite this article
Marinho, P.H., Fonseca, C.R., Sarmento, P. et al. Temporal niche overlap among mesocarnivores in a Caatinga dry forest. Eur J Wildl Res 66, 34 (2020). https://doi.org/10.1007/s10344-020-1371-6
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10344-020-1371-6