
Abstract
The enzymatic hydrolysis of the nitrile group of different 2-acetoxynitriles was investigated in order to obtain catalysts that chemoselectively hydrolyse nitriles in the presence of ester groups. The biotransformation of four 2-acetoxynitriles [2-acetoxybutenenitrile (ABN), 2-acetoxyheptanenitrile (AHN), 2-acetoxy-2-(2-furyl)acetonitrile (AFN), and 2-acetoxy-2,3,3-trimethylbutanenitrile (ATMB)] by different bacterial strains that synthesise nitrilases or nitrile hydratases was studied. ABN, AHN and AFN were converted by various microorganisms belonging to different bacterial genera (e.g. Pseudomonas or Rhodococcus) expressing either nitrilase or nitrile hydratase activities. In contrast, no metabolism of the sterically hindered substrate ATMB was observed. All wild-type strains investigated formed considerable amounts of cyanide and aldehydes from the 2-acetoxynitriles. This indicated the presence of esterases converting the 2-acetoxynitriles to 2-hydroxynitriles, which then spontaneously decomposed to the corresponding aldehydes and cyanide. In order to suppress unwanted side-reactions, biotransformations were performed with recombinant Escherichia coli strains that heterologously expressed nitrilase activities originating from Pseudomonas, Rhodococcus, or Synechocystis strains. The attempted conversion of the 2-acetoxynitriles to almost stoichiometric amounts of the corresponding 2-acetoxycarboxylic acids was finally achieved by using either a recombinant E. coli strain that highly overexpressed the nitrilase gene from the pseudomonad or the purified enzyme derived from this strain.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The hydrolysis of nitriles is of synthetic importance for the production of amides or carboxylic acids. The traditionally applied chemical reactions usually require rather harsh conditions, which may result in unwanted side reactions if the nitriles carry additional functional groups. One possible strategy for a more chemoselective conversion of additionally functionalised nitriles to amides and acids is the utilisation of nitrile-converting enzymes. Two different enzyme systems for the hydrolysis of nitriles are known: (1) direct hydrolysis of a nitrile to the carboxylic acid by nitrilases, or (2) a two-step reaction via amide by means of nitrile hydratases and amidases. Thus, these reactions enable production of either the corresponding amide or acid (Bunch 1998; Kobayashi and Shimizu 1994; Martínková and Křen 2002; Nagasawa and Yamada 1995; Schulze 2002).
One possible application for nitrile hydrolysing bacterial enzymes is the conversion of chiral α-hydroxynitriles, or derivatives thereof, which are accessible by the addition of HCN to aldehydes or ketones using R- or S-specific oxynitrilases of plant origin (Effenberger et al. 2000; Griengl et al. 2000). 2-Hydroxynitriles (cyanohydrins) generally exhibit low stability in aqueous solutions, and chemically decompose to the corresponding aldehyde or ketone plus cyanide. One possibility to stabilise the new chiral centre formed by the oxynitrilase reaction is acetylation of the 2-hydroxy-group, which prevents the spontaneous decomposition of the chiral product. The subsequent chemical hydrolysis of the nitrile group of these 2-acetoxynitriles to the corresponding 2-acetoxyamides or 2-acetoxycarboxylic acids is prone to the simultaneous deacetylation of the substrates (or products). Therefore, the hydration or hydrolysis of some aliphatic α-acetoxynitriles was previously attempted using the "nitrilase complex" SP409 from Novo Industri (Bagsvaerd, Denmark). However, it was found that fast deacetylation occurred during the reaction of the SP409 catalyst with 2-acetoxyheptanenitrile (AHN) and it was concluded that the commercial preparation contained significant esterase activities (de Raadt et al. 1992) and therefore could not be used for the hydrolysis of 2-acetoxynitriles.
We have previously described the isolation of various bacterial strains with the ability to convert a wide range of nitriles to the corresponding amides or carboxylic acids (Layh et al. 1997). In the present publication, we attempted to identify, from this strain collection and some genetically modified organisms, suitable biocatalysts with the ability to hydrolyse AHN and other aliphatic 2-acetoxynitriles to the corresponding 2-acetoxycarboxylic acids without hydrolysing the ester bonds of the 2-acetoxynitriles.
Materials and methods
Bacterial strains and cultivation of bacteria
The wild-type bacterial strains investigated were isolated previously from enrichment cultures with various nitriles as sole sources of nitrogen (Layh et al. 1992, 1997). Most of the strains are available from the DSMZ (Deutsche Sammlung für Mikroorganismen und Zellkulturen, Braunschweig, Germany) (see Table 1). The bacteria were routinely grown at 30°C with succinate (10 mM) as carbon source and the same nitriles as nitrogen sources (0.5–2 mM each) that had been used originally for the enrichment of the organisms.
The recombinant Escherichia coli strains used heterologously expressed nitrilases originating from the cyanobacterium Synechocystis spp. 6803, Rhodococcus rhodochrous NCIMB11216, or Pseudomonas fluorescens EBC191. The genes were amplified by PCR from the relevant genomic DNA using the published nucleotide sequences and cloned into a rhamnose-dependent expression system (pJOE2702) as described previously (Heinemann et al. 2003; Kiziak 1998; Ress-Löschke et al. 2000; Stumpp et al. 2000; Trott et al. 2001).
To induce nitrilase production, the recombinant E. coli strains were cultivated in Luria-Bertani broth (LB) plus ampicillin (100 μg/ml) at 30°C until the bacterial cultures reached an optical density at 546 nm (OD546) of approximately 0.02–0.2; 0.2% (w/v) l-rhamnose was then added and the cells were harvested after further growth for 6–13 h at 30°C.
Preparation of cell extracts
Cell suspensions in Tris/HCl buffer (50 mM, pH 7.5) were disrupted with a French Press (Aminco, Silver Springs, Md.) at 80 MPa. Cell debris was removed by centrifugation at 80,000 g and 4°C for 30 min. Protein was determined by the method of Bradford (1976) with bovine serum albumin as standard.
Purification of the His-tagged nitrilase
Cell extracts of E. coli JM109 (pDC12) were prepared in Tris/HCl buffer (50 mM, pH 7.5) as described above. Ni-NTA agarose (Qiagen, Hilden, Germany) was suspended in Tris/HCl (20 mM, pH 7.5) and transferred (2 ml each) to an empty 10 ml-polypropylene column. The filled column was equilibrated with a buffer system (pH 7.5) consisting of Tris/HCl (50 mM) and NaCl (100 mM). The cell extracts (50–70 mg protein) were applied to the column and the protein was eluted with subsequent steps of the Tris/HCl/NaCl buffer (2–4 ml each) with increasing imidazole concentrations (10–150 mM). The fractions with nitrilase activity eluted at an imidazole concentration of about 50 mM. This active protein fraction was finally dialysed for 60 min at 8°C against a 500-fold surplus of Tris/HCl buffer (20 mM, pH 7.5).
Analytical methods
The conversions of 2-acetoxy-3-butenenitrile (ABN) and 2-acetoxy-2-(2-furyl)acetonitrile (AFN) were analysed in most experiments by HPLC (HPLC Millenium Chromatography Manager 2.0, equipped with a programmable multi-wavelength detector model 486 and HPLC pump 510; Waters, Milford, Mass.). The solvent system consisted of 25 or 35% (v/v) methanol (for analysis of ABN or AFN turnover, respectively) in H2O with 0.3% (v/v) H3PO4, and a flow rate of 0.7 ml min−1 was applied. The separation of the individual compounds was achieved using a reversed-phase column (150×4 mm; Grom-Sil C8; Grom, Herrenberg, Germany). The compounds were detected using a UV/Vis detector at a fixed wavelength of 210 nm.
Conversion of AHN (and, in certain experiments, also of AFN and ABN) was analysed by gas chromatography (GC) using a Chrompack CP9001 gas chromatograph (Chrompack, Frankfurt, Germany). The individual compounds were separated on a cyclodextrin matrix as stationary phase (FS-Hydrodex β-PM, 45 m ×0.25 mm; Macherey & Nagel, Düren, Germany) with nitrogen gas as mobile phase (0.8 ml/min) and detected using a flame ionisation detector (FID) (250°C) (= GC system 1). The injector temperature was set at 220°C. For the separation of nitriles, amides and carboxylic acids, a linear temperature gradient from 110°C to 220°C was applied.
In certain experiments for the simultaneous analysis of the acetoxy-compounds and their deacetylated products, a Hewlett Packard Model 5890 GC equipped with a capillary column HP 5 (30 m ×0.32 mm ×0.25 μm) and a split/splitless injector plus FID was used (= GC system 2). The injector temperature was fixed at 250°C and the detector temperature at 280°C. A temperature program was used that started at 40°C for 3 min and a linear temperature gradient was applied with a heating rate of 10°C per minute reaching a final temperature of 250°C. The carrier gas was nitrogen and a flow rate of 1.94 ml/min was used.
The concentrations of ammonia and cyanide were determined spectrophotometrically using commercial test kits (Spectroquant; Merck, Darmstadt, Germany).
Standard assay for the hydrolysis of nitriles with resting cells
In the standard experiments, resting cells (approximately 2–4 mg bacterial dry weight in 10 ml) were incubated in 50 mM Na/K-phosphate buffer, pH 7.4, with the relevant nitriles (5 mM each) at 30°C on a rotary shaker. After different time intervals (about 2–20 min), aliquots (500 μl) were taken and the cells removed by centrifugation in an Eppendorf centrifuge at 16,000 g. The supernatants were acidified with HCl to approximately pH 2 and samples were either analysed directly by HPLC or prepared for GC analysis by extraction of the substrates and products into an equal volume of dichloromethane.
In certain experiments, the reactions were scaled up and performed in a total volume of 20 ml and aliquots of 1.5 ml were taken. The reactions were stopped by centrifugation, and the substrates and products extracted from these samples twice into 1.5 ml ethylacetate. The organic phase was separated from the aqueous phase, dried with Na2SO4 and the volume of the sample reduced by evaporation to 0.1 ml.
Chemicals
Racemic ABN, AHN, and AFN were synthesised according to the literature (Gassman and Talley 1978) employing trimethylsilyl cyanide followed by acidic hydrolysis of the silyl ether and acetylation. The free cyanohydrins were hydrolysed with concentrated HCl at ambient temperature to give the corresponding amides; the analogous hydroxy acids were obtained at reflux temperature and the amides and acids were finally acetylated. All other chemicals, media, and procedures were as previously described (Bauer et al. 1994; Layh et al. 1992, 1994).
Results
Conversion of ABN, AHN, and AFN by various bacteria with nitrilase or nitrile-hydratase/amidase activities
We have previously described the isolation of various bacterial strains with the ability to utilise aromatic nitriles such as 2-phenylpropionitrile, 2-(2-methoxyphenyl)propionitrile, ibuprofen nitrile, 2-phenylbutyronitrile, naproxen nitrile, ketoprofen nitrile, benzonitrile, or naphthalene-2-carbonitrile as sole source of nitrogen (Layh et al. 1997). In the present study, these isolates were also tested for their ability to convert aliphatic α-acetoxynitriles. For this purpose, the bacteria were cultivated with the respective aromatic nitriles that had been used for their enrichment and resting cells were incubated with ABN, AHN, AFN, or 2-acetoxy-2,3,3-trimethylbutanenitrile (ATMB) (for structural formulas see Fig. 1). Analysis of the reactions by GC demonstrated that many of the isolates were able to convert ABN, AHN, and AFN (Table 1). In contrast, no significant turnover of ATMB was observed with any of the strains. GC analysis clearly demonstrated that the bacterial strains did not convert the acetoxynitriles to stoichiometric amounts of the desired products but formed significant amounts of side-products. This suggested that the cells also expressed esterase activities, as also observed by de Raadt et al. (1992) with the nitrile-hydrolysing whole cell catalyst SP409 previously distributed by Novo Industri. Deacetylation by esterases should result in the formation of unstable 2-hydroxynitriles, which were expected to spontaneously decompose in the aqueous medium to the corresponding aldehydes and cyanide (Fig. 2). Indeed, HPLC and GC analysis, and also a spectrophotometric cyanide assay, proved that all strains investigated (with nitrilase and also with nitrile hydratase/amidase-systems) formed significant amounts of cyanide and the corresponding aldehydes from all three 2-acetoxynitriles.

Chemical structures of the 2-acetoxynitriles studied. A 2-acetoxybutenenitrile (ABN), B 2-acetoxyheptanenitrile (AHN), C 2-acetoxy-2-(2-furyl)acetonitrile (AFN), D 2-acetoxy-2,3,3-trimethylbutanenitrile (ATMB)

Schematic representation of different enzymatic and chemical reactions involved in the conversion of 2-acetoxynitriles
Formation of 2-acetoxyheptaneamide by strains with nitrile hydratase activity
Our previous study (Layh et al. 1997) suggested that certain of the isolates synthesised nitrilases and others converted nitriles via nitrile-hydratase/amidase systems (Table 1). This observation was confirmed when the conversion of 2-acetoxyheptanenitrile was analysed in greater detail (using GC system 2, see Materials and methods). Thus it was found that bacteria harbouring nitrilases converted AHN to 2-acetoxyheptanoic acid. In contrast, strains that synthesised a nitrile-hydratase/amidase system formed only small amounts of 2-acetoxyheptanoic acid and a second product identified as 2-acetoxyheptanoic acid amide by comparison with a chemically synthesised reference compound.
Influence of phenylmethylsulfonylfluoride on the esterase activity in P. fluorescens EBC191
The results described above clearly demonstrated that all strains investigated possessed pronounced esterase activities that would preclude utilisation of these biocatalysts for chemoselective hydrolysis of the nitrile group of acetoxynitriles. Therefore, a model system was used to test if these esterase activities could be inactivated by using the esterase inhibitor phenylmethylsulfonylfluoride (PMSF) (Zollner 1989). Cell extracts from the nitrilase-synthesising strain P. fluorescens EBC191 were incubated with different concentrations of PMSF and the turnover of AFN was analysed (Fig. 3). The addition of PMSF indeed reduced the amount of deacetylated products, but the turnover of the nitrile also decreased in the presence of PMSF. Furthermore, it was observed that the cell extracts produced small amounts of the 2-acetoxyamide from AFN in either the presence or absence of PMSF (see Fig. 3). The formation of some amides from nitriles by the action of nitrilases has been previously reported (Effenberger and Oßwald 2001; Piotrowski et al. 2001; Stevenson et al. 1992).

Influence of phenylmethylsulfonylfluoride (PMSF) on product formation during the hydrolysis of AFN by cell-free extracts from Pseudomonas fluorescens EBC191. The reaction mixtures contained Na/K phosphate buffer (50 mM, pH 7.5), cell-free extract (0.32 mg protein), 5 mM AFN, and 0 mM (A), 1 mM (B), or 5 mM (C) PMSF in a total volume of 3.5 ml. The reaction mixtures were incubated at 30°C in a thermomixer (300 strokes/min). Aliquots (200 μl) were taken after different time intervals and the reactions terminated by the addition of 10% (v/v) 1 M HCl. The concentrations of AFN (●), furfural (■), and 2-acetoxy-2-(2-furyl)acetic acid (◊) were analysed by HPLC. The concentration of cyanide (□) was quantified using a commercially available cyanide test system
Conversion of 2-acetoxynitriles by E. coli strains heterologously expressing nitrilases from Synechocystis spp. PCC6803, R. rhodochrous NCIMB 11216, and P. fluorescens EBC191
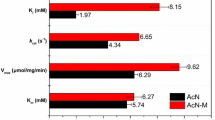
The results described above suggested that the desired stoichiometric conversion of 2-acetoxynitriles to the corresponding 2-acetoxycarboxylic acids was not possible with the bacterial isolates available to us due to the ubiquitous presence of esterase activities in these bacteria. It was therefore decided to perform experiments with recombinant E. coli strains heterologously expressing nitrilase genes from different sources. These nitrilase genes were under the control of a strong promoter that is induced by the addition of rhamnose (Stumpp et al. 2000). It was expected that these constructs would express a much higher level of nitrilase activity compared to the wild-type strains. The nitrilases investigated were originally obtained from P. fluorescens EBC191 (Layh et al. 1992; Kiziak 1998), Synechocystis spp. PCC6803 (Heinemann et al. 2003), and R. rhodochrous NCIMB 11216 (Harper 1977; Ress-Löschke et al. 2000). All three recombinant E. coli strains were able to convert ABN and AFN. The recombinant E. coli strains producing the nitrilases from P. fluorescens EBC191 and R. rhodochrous NCIMB 11216 produced nitrilase activities about 13- to 17-times higher than those of the respective wild-type strains. (A similar comparison could not be performed for the Synechocystis nitrilase, because it is not known under which conditions the nitrilase activity is induced in Synechocystis spp. PCC6803; Heinemann et al. 2003). E. coli strains expressing recombinant nitrilase from P. fluorescens EBC191 showed significantly higher activities for both substrates than strains expressing the Rhodococcus or Synechocystis nitrilases (Table 2).
A comparison of the substrate conversion rates of ABN and AFN by the nitrilase-synthesising wild-type strain of P. fluorescens EBC191 and the recombinant E. coli strain synthesising the Pseudomonas nitrilase clearly demonstrated that the recombinant organism produced significantly more of the desired 2-acetoxycarboxylic acids compared to the wild-type strain (Fig. 4). The resting cells of the recombinant organism showed specific activities with AFN and ABN of 1.0 and 0.4 U/mg protein, respectively, and did not produce detectable amounts of the corresponding aldehydes. In contrast, traces of the aldehydes could be detected with E. coli strains synthesising Rhodococcus or Synechocystis nitrilases. These recombinant strains produced significantly lower nitrilase activities compared to the Pseudomonas nitrilase with the 2-acetoxynitriles tested although also in these strains the nitrilases represented more than 10% of the soluble proteins. A comparison of the turnover rates of the 2-acetoxynitriles in comparison to the preferred substrates of the nitrilases (benzonitrile for the Rhodococcus nitrilase and fumarodinitrile for the Synechocystis nitrilase) suggested that for both enzymes the relative conversion rate of all acetoxynitriles was less than 5% of the activity found with the best substrates.

Comparison of the hydrolysis of ABN and AFN by P. fluorescens EBC 191 and Escherichia coli (pDC12) heterologously expressing the nitrilase from P. fluorescens EBC191. A liquid culture of P. fluorescens EBC191 was grown with succinate as source of carbon and energy and ethylbenzylcyanide as nitrogen source. E. coli (pDC12) was cultivated in LB medium plus ampicillin (100 μg/ml) and nitrilase activity was induced by the addition of rhamnose (0.2% w/v) as described in Materials and methods. The cells were harvested by centrifugation and resuspended in Na/K-phosphate buffer (50 mM, pH 7.4) to optical densities at 546 nm (OD546) of 1 (E. coli) or 2.5 (P. fluorescens EBC 191). AFN or ABN were added to initial concentrations of 4 mM and the cells incubated in a thermomixer at 30°C (300 strokes/min). Aliquots were removed after different time intervals, cells precipitated by centrifugation, and the supernatants acidified with 10% (v/v) 1 M HCl. The concentrations of the respective nitrile (●), 2-acetoxy acid (▢), 2-acetoxy amide (◇), and the corresponding aldehyde (■) were determined by HPLC and the concentration of cyanide (□) using the Spectroquant test
The formation of traces of the aldehydes by E. coli strains synthesising the Rhodococcus or Synechocystis nitrilases suggested that some relevant esterase activities were also present in E. coli, but that these activities were negligible if the cells expressed high nitrilase activities.
The recombinant E. coli strain synthesising the nitrilase from P. fluorescens EBC191 produced small amounts of the 2-acetoxyamides from AFN and ABN as had been observed during the conversion of AHN by the wild-type strain of P. fluorescens EBC191.
Conversion of 2-acetoxynitriles by E. coli JM109
The observation that the E. coli whole cell catalysts still produced some deacetylated products suggested that the E. coli strain used also possessed some esterase activities for the hydrolysis of 2-acetoxynitriles. Therefore, E. coli JM109 (without the recombinant nitrilase genes) was incubated with the relevant 2-acetoxynitriles (5 mM each) and specific activities of about 0.004–0.03 U/mg protein were determined. GC analysis confirmed that E. coli JM109 indeed formed 2-hydroxyheptanenitrile from AHN and 2-hydroxy-2-(2-furyl)acetonitrile from AFN. In contrast, no turnover of 2-methylbutyronitrile or 2-methylcapronitrile was found with the control strain.
Conversion of ABN and AFN by the purified nitrilase from P. fluorescens EBC191
The results described above indicate that esterase activities that hydrolyse the ester bonds of compounds such as ABN and AFN are almost ubiquitous in bacteria and that minor esterase activities are also present in E. coli. It was therefore decided to perform the attempted biotransformations using the purified nitrilase from P. fluorescens EBC191. Under these conditions, the conversion of ABN and AFN was also achieved without the formation of detectable amounts of deacetylated side products (Fig. 5).

Hydrolysis of ABN and AFN by the purified nitrilase from P. fluorescens EBC191. The His-tagged nitrilase was purified on a Ni-NTA column. The reaction mixtures contained 20 mM Na/K-phosphate buffer, the purified nitrilase (40 μg/ml) and the respective acetoxynitriles (20 mM each) in a total volume of 250 μl. The samples were incubated in a thermomixer (30°C, 300 strokes/min). Aliquots (50 μl) were taken at different time intervals. The reactions were stopped by the addition of 5 μl 1 M HCl and precipitated protein removed by centrifugation (16,000 g, 3 min). The concentrations of the 2-acetoxynitriles (●), 2-acetoxy amides (◇), 2-acetoxy acids (○), and the corresponding aldehydes (■) were determined by HPLC
Discussion
There are only very few publications that describe the hydrolysis of aliphatic 2-hydroxynitriles or 2-acetoxynitriles by microorganisms. Thus, it has been shown that the yeast Torulopsis candida GN405 converted dl-2-hydroxyisovaleronitrile and dl-2-hydroxyisocapronitrile preferentially to the corresponding l-acids (Fukuda et al. 1973). Furthermore, different Rhodococcus strains (R. butanica ATCC 21197 and the SP409 catalyst previously distributed by Novo Industri) were shown to hydrolyse 2-hydroxydodecanoic acid nitrile and its acetoxy-derivative as well as 2-hydroxy-2,3-dimethylbutyronitril, 2-hydroxycyclohexanenitrile and 2-acetoxyheptanenitrile (de Raadt et al. 1992; Kakeya et al. 1991). Also, some aromatic 2-hydroxynitriles and 2-acetoxynitriles were successfully—even enantioselectively—hydrolysed using bacterial nitrilases. For instance, a preference for the formation of R-O-acetylmandelic acid from racemic (R,S)-O-acetylmandelonitrile was demonstrated for the nitrilase from P. fluorescens EBC191 (Layh et al. 1992). Likewise, the enantioselective hydrolysis of racemic (R,S)-mandelonitrile to R-mandelic acid was shown for a nitrilase from Alcaligenes faecalis ATCC 8750 (Yamamoto et al. 1992).
It has previously been suggested that nitrilases be grouped according to their substrate preference as "aromatic nitrilases", "aliphatic nitrilases", or "arylacetonitrilases" (Kobayashi et al. 1990; Nagasawa et al. 1990). It was therefore a surprising observation that all organisms and enzymes tested in the present study were able to hydrolyse aliphatic O-acetoxynitriles provided that the chiral carbon atom carried at least one hydrogen substituent. Furthermore, it became evident that the nitrilase from P. fluorescens EBC191, which, according to its substrate preference, is a typical "arylacetonitrilase" (Layh et al., 1992), demonstrated much higher specific activities with the aliphatic O-acetoxynitrile ABN compared to the nitrilase from Synechocystis spp. 6803, which has been shown to convert several aliphatic nitriles (Heinemann et al. 2003). This biochemical evidence, together with some recently performed sequence alignments of different nitrilases (Heinemann et al. 2003), suggested that for a valid classification of nitrilases it is necessary to perform a thorough comparison of the substrate specificities of the enzymes with a wide range of different substrates.
The results obtained in the present study with different bacterial strains suggest that esterases with the ability to hydrolyse aliphatic acetoxynitriles are almost ubiquitously distributed among bacteria of different taxonomic groups. It therefore appears rather unlikely that a wild-type bacterium can be obtained that performs a stoichiometric conversion of 2-acetoxynitriles to the corresponding 2-acetoxy acids. Unfortunately, utilisation of E. coli cells heterologously overexpressing nitrilases from different genetic backgrounds also resulted in some ester hydrolysis when only low nitrilase activities were present in the recombinant organisms. Therefore, it can be deduced that cells of E. coli also synthesise some relevant esterase activities. The existence of several esterases in the latter organism has been previously demonstrated biochemically (Goullet 1973; Goullet et al. 1984; Haruki et al. 1999; Peist et al. 1997), and can be deduced from the genome sequence, but these enzymes have never been tested with substrates that resemble those analysed in the present study. Therefore, it will be necessary to perform biotransformations of 2-acetoxynitriles using either E. coli whole cell catalysts that demonstrate rather high specific activities for conversion of the nitrile groups, or purified nitrilase.
References
Bauer R, Hirrlinger B, Layh N, Stolz A, Knackmuss H-J (1994) Enantioselective hydrolysis of racemic 2-phenylpropionitrile and other (R,S)-2-arylpropionitriles by a new bacterial isolate, Agrobacterium tumefaciens strain d3. Appl Microbiol Biotechnol 42:1–7
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal Biochem 72:248–254
Bunch AW (1998) Nitriles. In: Rehm HJ, Reed G (eds) Biotechnology, vol 8a. Biotransformations I. Wiley-VCH Weinheim, pp 277–324
Effenberger F, Oßwald S (2001) Enantioselective hydrolysis of (RS)-2-fluoroarylacetonitriles using nitrilase from Arabidopsis thaliana. Tetrahedron Asymmetry 12:279–285
Effenberger F, Förster S, Wajant H (2000) Hydroxynitrile lyases in stereoselective catalysis. Curr Opin Biotechnol 11:532–539
Fukuda Y, Harada T, Izumi Y (1973) Formation of l-α-hydroxyacids from d,l-α-hydroxynitriles by Torulopsis candida GN405. J Ferment Technol 51:393–397
Gassman PG, Talley JJ (1978) Cyanohydrins—a general synthesis. Tetrahedron Lett 40:3773–3776
Goullet P (1973) An esterase zymogram of Escherichia coli. J Gen Microbiol 77:27–35
Goullet P, Picard B, Laget PF (1984) Purification and properties of carboxylesterase B of Escherichia coli. Ann Microbiol (Paris) 135A:375–387
Griengl H, Schwab H, Fechter M (2000) The synthesis of chiral cyanohydrins by oxynitrilases. Trends Biotechnol 18:252–256
Harper DB (1977) Microbial metabolism of aromatic nitriles. Enzymology of the C-N cleavage by Nocardia sp. (Rhodochrous group) N.C.I.B. 11216. Biochem J 165:309–319
Haruki M, Oohashi Y, Mizuguchi S, Matsuo Y, Morikawa M, Kanaya S. (1999) Identification of catalytically essential residues in Escherichia coli esterase by site-directed mutagenesis. FEBS Lett 454:262–266
Heinemann U, Engels D, Kiziak C, Mattes R, Stolz A (2003) Cloning, heterologous expression and enzymatic characterization of a nitrilase from the cyanobacterium Synechocystis spp. PCC6803. Appl Environ Microbiol 69: (in press)
Kakeya H, Sakai N, Sugai T, Ohta H (1991) Preparation of optically active α-hydroxy acid derivatives by microbial hydrolysis of cyanhydrins and its application to the synthesis of (R)-4-dodecanolide. Agric Biol Chem 55:1877–1881
Kiziak C (1998) Heterologe Expression der Nitrilase aus Pseudomonas fluorescens EBC191 und chimärer Enzymvarianten in E. coli. Reinigung, Stabilisierung und biochemische Charakterisierung der Enzyme. Diplomarbeit, Universität Stuttgart
Kobayashi M, Shimizu S (1994) Versatile nitrilases: nitrile-hydrolysing enzymes. FEMS Microbiol Lett 120:217–224
Kobayashi M, Yanaka N, Nagasawa T, Yamada H (1990) Purification and characterization of a novel nitrilase of Rhodococcus rhodochrous K22 that acts on aliphatic nitriles. J Bacteriol 172:4807–4815
Layh N, Stolz A, Förster S, Effenberger F, Knackmuss H-J (1992) Enantioselective hydrolysis of O-acetylmandelonitrile to O-acetylmandelic acid by bacterial nitrilases. Arch Microbiol 158:405–411
Layh N, Stolz A, Böhme J, Effenberger F, Knackmuss H-J (1994) Enantioselective hydrolysis of racemic naproxen nitrile and naproxen amide to S-naproxen by new bacterial isolates. J Biotechnol 33:175–182
Layh N, Hirrlinger B, Stolz A, Knackmuss H-J (1997) Enrichment strategies for nitriles hydrolysing bacteria. Appl Microbiol Biotechnol 47:668–674
Martínková L, Křen V (2002) Nitrile- and amide-converting microbial enzymes: stereo-, regio- and chemoselectivity. Biocatal Biotransform 20:79–93
Nagasawa T, Yamada H (1995) Microbial production of commodity chemicals. Pure Appl Chem 67:1241–1256
Nagasawa T, Mauger J, Yamada H (1990) A novel nitrilase, arylacetonitrilase, of Alcaligenes faecalis JM3. Eur J Biochem 194:765–772
Peist R, Koch A, Bolek P, Sewitz S, Kolbus T, Boos W (1997) Characterization of the aes gene of Escherichia coli encoding an enzyme with esterase activity. J Bacteriol 179:7679–7686
Piotrowski M, Schönfelder S, Weiler EW (2001) The Arabidopsis thaliana isogene NIT4 and its orthologs in tobacco encode β-cyano-l-alanine hydratase/nitrilase. J Biol Chem 276:2616–2621
Raadt A de, Klempier N, Faber K, Griengl H (1992) Chemoselective enzymatic hydrolysis of aliphatic and alicyclic nitriles. J Chem Soc Perkin Trans I:137–140
Ress-Löschke M, Hauer B, Mattes R, Engels D (2000) Nitrilase aus Rhodococcus rhodochrous NCIMB 11216. German patent application No DE 10010149A1
Schulze B (2002) Hydrolysis and formation of C-N bonds. In: Drauz K, Waldmann H (eds) Enzyme catalysis in organic synthesis, vol II. Wiley-VCH, Weinheim, pp 699–715
Stevenson DE, Feng R, Dumas F, Groleau D, Mihoc A, Storer AC (1992) Mechanistic and structural studies on Rhodococcus ATCC 39484 nitrilase. Biotechnol Appl Biochem 15:283–302
Stumpp T, Wilms B, Altenbuchner J (2000) Ein neues, l-Rhamnose-induzierbares Expressionssystem für Escherichia coli. Biospektrum 6:33–36
Trott S, Bauer R, Knackmuss H-J, Stolz A (2001) Genetic and biochemical characterization of an enantioselective amidase from Agrobacterium tumefaciens d3. Microbiology 147:1815–1824
Yamamoto K, Fujimatsu I, Komatsu K-I (1992) Purification and characterization of the nitrilase from Alcaligenes faecalis ATCC 8750 responsible for enantioselective hydrolysis of mandelonitrile. J Ferment Bioeng 73:425–430
Zollner H (1989) Handbook of enzyme inhibitors. VCH, Weinheim, Germany
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Heinemann, U., Kiziak, C., Zibek, S. et al. Conversion of aliphatic 2-acetoxynitriles by nitrile-hydrolysing bacteria. Appl Microbiol Biotechnol 63, 274–281 (2003). https://doi.org/10.1007/s00253-003-1382-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-003-1382-8