
Abstract
The invasion of native riparian forests by exotic tree species can lead to profound changes in the ecological integrity of freshwater ecosystems. We assessed litter decomposition of native (Alnus glutinosa and Quercus robur) and invasive (Acacia melanoxylon and Acacia dealbata) tree species, and associated microbial activity and community structure, after being immersed for conditioning in 3 reference and 3 “invaded” streams in Serra da Lousã (central Portugal) and used in microcosms simulating stream conditions. Litter decomposition differed among species, in the order: Al. glutinosa > Q. robur > (Ac. melanoxylon ~ Ac. dealbata). Alnus glutinosa litter decomposed faster probably because it was soft and had high nitrogen concentration for decomposers. Quercus robur litter decomposed slower most likely because it was tough and had high polyphenol and low nitrogen concentrations. Acacia melanoxylon litter was the toughest and had a thick cuticle that likely acted as a physical barrier for microbial colonization. In Ac. dealbata, the small-sized leaflets and high lignin concentration may have limited microbial litter decomposition. Litter decomposition was faster in “invaded” streams, probably because they were N-limited and increases in nitrogen concentration in water, promoted by Acacia species invasion, stimulated microbial activity on litter. The aquatic hyphomycete community structure differed among litter species and between stream types, further suggesting that microbes were sensitive to litter characteristics and water nutrient concentrations. Overall, the invasion of native riparian forests by Acacia species may affect microbial decomposer activity, thus altering important stream ecosystem processes, such as litter decomposition and nutrient cycles.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The invasion of native forests by exotic plant species is occurring worldwide and is expected to increase due to human activities (e.g., agriculture, urbanization) and climate change (e.g., increased temperature, altered precipitation) [1, 2]. Plant invasions are expected to change native forest composition and alter the diversity of plant functional traits [2]. Therefore, plant invasions can have profound effects on detrital ecosystems, which depend on plant matter inputs [2,3,4].
In forest streams, the plant matter supplied by the riparian vegetation (e.g., leaves, wood) represents a major source of organic carbon for aquatic food webs [5]. In these ecosystems, plant litter decomposition is a fundamental ecosystem process mediated by the activities of microbes, namely fungi and bacteria, and invertebrate detritivores [6]. Fungi, particularly aquatic hyphomycetes, and bacteria produce a large variety of extracellular enzymes that promote leaf maceration and increase leaf palatability to invertebrate detritivores [6, 7]. Aquatic hyphomycetes are key decomposers of litter particularly in the initial stages of the process, while bacteria only increase their contribution when litter has been partially broken down [8]. Altogether, aquatic decomposers mineralize the organic carbon from litter, convert it into biomass, and promote the release of dissolved and fine organic particles that cause leaf mass loss [7]. Microbial decomposers respond to changes in litter characteristics and nutrient concentrations in stream water. Microbes generally colonize litter of high nutritional quality (i.e., soft litter, with high nutrient concentration and low concentrations of structural and secondary compounds) and decompose it faster than more recalcitrant litter [9,10,11,12], and have higher activity under nutrient-rich conditions [10, 13, 14]. Additionally, aquatic hyphomycete community structure is sensitive to resource diversity (e.g., leaf litter), being more diverse when resource diversity (i.e., riparian vegetation and litter inputs) is higher [15,16,17]. Also, some aquatic hyphomycete species can occur as endophytes, and thus, increases in riparian diversity may promote an increase in aquatic hyphomycete diversity due to host specificity between endophytes and trees [18].
Over the past 250 years, many Australian Acacia species have been introduced into several regions of the world [19], mainly for wood and timber production, to stabilize dunes and to prevent soil erosion [20]. Specific traits such as resistant seeds that quickly germinate, capacity to fix atmospheric nitrogen (N), high capacity to adapt to different environmental conditions, rapid growth rate, and high accumulation of biomass are responsible for their successful invasive behavior [20, 21]. In Portugal, Acacia species cover at least ~ 17,000 ha [22], and it is forbidden to further introduce and use the entire Acacia genus for afforestation [23]. Acacia species are among the 10 most problematic invasive exotic species in Portugal and among the 30 most problematic in Spain and France [24]. Invasion of native forests by Acacia species is expected to affect the characteristics of litter inputs to streams as diverse deciduous forests with low abundances of N-fixers are being replaced by monospecific stands of evergreen N-fixing species [20]. Moreover, N concentrations in stream water were expected to increase as found for streams with riparian areas invaded by other exotic N-fixing species [25,26,27,28,29]. Given the sensitivity of microbial decomposers to litter characteristics and dissolved nutrient concentrations, changes in these parameters can have strong effects on litter decomposition in “invaded” streams.
Currently, in central Portugal, many native riparian deciduous forests dominated by Castanea sativa Mill. and Quercus robur L. are being invaded by exotic N-fixing species (mostly Acacia dealbata Link. and to a lesser extent Acacia melanoxylon R. Br.). Here, we examine the effects of litter identity and quality (native species: Alnus glutinosa (L.) Gaertn and Q. robur; invasive species: Ac. melanoxylon and Ac. dealbata) and dissolved nutrient concentrations (nutrient levels from reference and “invaded” streams) on litter decomposition and associated microbial decomposers in laboratory microcosms simulating stream conditions. We predict that (i) litter with high nutritional quality (soft, high initial N concentration) will have higher microbial activity and faster litter decomposition; (ii) microbial activity will be higher, and litter decomposition will be faster in “invaded” than reference streams due to a stimulation of microbial activity by increased N concentrations in stream water; and (iii) litter in reference streams will have higher aquatic hyphomycete species richness due to a higher inoculum potential as a result of higher resource diversity.
Material and Methods
Study Region
Six small forest streams (< 4 m wide), located in Serra da Lousã (central Portugal), were selected for this experiment (Table 1). Reference streams (Cerdeira, Candal, and Maior) run through native forests mainly composed of C. sativa and Q. robur trees, while “invaded” streams (Sotão, Fiscal, and Piedade) run through forests heavily invaded by Acacia species (94–100% cover in the riparian area defined as 50 m to each side of the stream and 250 m long is composed mostly by Ac. dealbata (> 90%) and to a lesser extent by Ac. melanoxylon (< 10%)). All streams run on schist bedrock. Stream basins have low human occupation, and agricultural activity is nearly absent (< 0.5% of basin area is occupied by agriculture).
Litter Species and Initial Chemical Characteristics
Litter of two native species, Al. glutinosa and Q. robur, and of two invasive species, Ac. melanoxylon and Ac. dealbata, commonly found in the riparian vegetation of streams flowing through native and “invaded” forests of central Portugal, respectively, were selected for this study. The native species were selected among the dominant riparian species to represent extremes in terms of leaf litter N concentration [30]. Alnus glutinosa is an actinorhizal species that establishes symbiosis with the N-fixing actinobacteria Frankia alni, thus acting as an N-fixing tree species, while Q. robur is not an N-fixer. Acacia species are N-fixers, but while Ac. melanoxylon has phyllodes (i.e., leaf-like structures derived from modified petioles), Ac. dealbata has bipinnate leaves with a large number of small leaflets (2–5 mm long and 0.4–0.7 mm wide).
Litter was collected in Serra da Lousã, central Portugal, in autumn 2016. Litter of Al. glutinosa, Q. robur, and Ac. melanoxylon were collected just after abscission from the ground in small tree stands dominated by the target species, where trees were exposed to similar environmental and edaphic conditions and appeared to have similar age and vigor. Acacia dealbata litter was collected directly from a few trees of similar age and vigor in an Ac. dealbata dominated stand. Litter of each species was pooled together, air-dried at room temperature, and stored in the dark until used.
Three sets of air-dried litter from each species (randomly selected from each species litter pool) were oven-dried (105 °C, ± 48 h) and ground to a fine powder (< 0.5 mm size; Retsch, ZM 100 Ultra Centrifugal Mill, Haan, Germany). Then, the powder was oven-dried (105 °C, ± 48 h) and analyzed to determine the initial polyphenols [31], lignin [32], phosphorus [33], carbon (C), and nitrogen (N) (Thermo Fisher Scientific Inc., CNH auto analyzer IRMS Thermo Delta V advantage with a Flash EA, 1112 series, Waltham, USA) concentrations of each species prior to the beginning of the experiment. Results were expressed as percentage of dry mass (DM).
Microbial Colonization of Litter
Air-dried litter of Al. glutinosa, Q. robur, Ac. melanoxylon, and Ac. dealbata were enclosed individually in fine-mesh bags (0.5 mm mesh size; 5 litter bags per litter species) ad libitum, and litter bags were immersed in the three reference and the three “invaded” streams on March 2017 to allow microbial colonization. On the same occasion, conductivity, water temperature, pH, and dissolved O2 concentration were measured in situ with field probes (WTW, Weilheim, Germany). Stream water (± 8 L) was also collected in acid-washed plastic bottles from each stream, transported in a cooler to the laboratory, filtered through glass microfiber filters (47 mm diameter, 0.7 μm pore size; Whatman GF/F, GE Healthcare UK Limited, Little Chalfont, UK), and frozen at − 20 °C until needed. Stream water was used for the microcosm experiment and for determination of nutrient concentrations. Nitrite, nitrate, and ammonium concentrations were determined by the colorimetric method (AA3 Bran + Luebbe autoanalyzer; SEAL Analytical, Norderstedt, Germany).
Microcosm Setup
After 1 week of colonization in the streams, litter bags were collected, placed individually in zip-lock bags, and transported in a cooler to the laboratory. In the laboratory, litter was rinsed with distilled water. Discs from Al. glutinosa, Q. robur, and Ac. melanoxylon litter were cut with a cork borer (12-mm diameter; avoiding the main vein for Al. glutinosa and Q. robur), and leaflets from Ac. dealbata were detached from the secondary veins. Litter discs (20 per microcosm) and portions of leaflets were wet-weighed (± 0.1 mg) and transferred to 100-mL Erlenmeyer flasks containing 40 mL of filtered water from the stream of origin. Microcosms were stored on an orbital shaker (100 rotations minute−1; GFL 3017, Burgwedel, Germany) at 18 °C, under a 12-h light and 12-h dark regime. Each stream within each stream type acted as a replicate, i.e., Cerdeira, Maior, and Candal were the three replicate streams for the reference stream type and Sotão, Fiscal, and Piedade were the three replicate streams for the “invaded” stream type (4 litter species × 2 stream types × 3 replicate streams × 4 sampling dates = 96 microcosms) (Fig. 1). The water in the microcosms was renewed twice a week using filtered water from the stream of origin. After 14, 31, 49, and 70 days of incubation in the laboratory, three microcosms per treatment (i.e., one flask per litter species per stream) were sacrificed for the determination of microbial respiration, fungal biomass, conidia production by aquatic hyphomycetes, litter toughness, and litter mass loss (see below).

Experimental design showing that litter of Alnus glutinosa, Quercus robur, Acacia melanoxylon, and Acacia dealbata was enclosed in fine-mesh bags and immersed for conditioning during 7 days in three reference (Cerdeira, Candal and Maior) and three “invaded” streams (Sotão, Fiscal, and Piedade), in Serra da Lousã (central Portugal). Sets of 20 discs were cut from Al. glutinosa, Q. robur, and Ac. melanoxylon litter and portions of leaflets were detached from Ac. dealbata litter and incubated in flasks containing water from the stream of origin. Three microcosms per treatment were sacrificed after 14, 31, 49, and 70 days of incubation for mass remaining and microbial determinations
Microbial Respiration
On each sampling date, a subset of five litter discs or a portion of leaflets from each sacrificed microcosm was used to determine the microbial oxygen consumption rates using a closed six-channel dissolved oxygen measuring system (Strathkelvin 929 System, North Lanarkshire, UK). A saturated solution of sodium sulfite in 0.01 M sodium borate (0% O2; prepared immediately before use) and an O2-saturated water sample from the stream of origin (100% O2, kept at 18 °C) were used to calibrate the oxygen electrodes. Then, litter discs and portions of leaflets were incubated in 3-mL chambers with the corresponding saturated stream water kept at 18 °C. Chambers without plant material were used as controls. The system was left running for ± 45 min. Then, litter discs and leaflets were removed from the chambers and used to determine litter toughness (see below) and litter discs DM. Oxygen consumption rates were determined by the difference in the O2 concentration in the sample and the control, at the beginning and the end of a 15-min interval during which O2 consumption over time was linear (taken between 10 and 40 min of incubation). Results were expressed as mg O2 g−1 DM h−1.
Fungal Biomass
Another subset of five litter discs and a portion of leaflets were frozen at − 20 °C, lyophilized overnight, weighed (± 0.1 mg) to determine DM and used for ergosterol extraction [31, 34]. Litter discs and leaflets were placed in tightly closed tubes with 10 mL of KOH in methanol (8 g L−1). Lipids were extracted in an 80 °C water bath for 30 min, purified by solid phase extraction (Waters Sep-Pak Vac RC tC18 cartridges; Waters Corp., Milford, USA) and eluted with isopropanol. Ergosterol was quantified by high-performance liquid chromatography (HPLC; Dionex DX-120, Sunnyvale, CA, USA) using a Thermo Scientific Syncronis C18 column (250 × 4 mm, 5 μm particle size; Thermo, Waltham, MA, U.S.A.) and HPLC-grade methanol as the mobile phase (1.4 mL min−1 at 33 °C). Ergosterol was detected at 282 nm, quantified based on a standard curve of ergosterol in isopropanol, and converted into mycelial biomass assuming 5.5 μg ergosterol mg−1 fungal DM [34]. Results were expressed as mg fungal DM g−1 litter DM.
Conidia Production by Aquatic Hyphomycetes
On each sampling date, the conidial suspension from each sacrificed microcosm was saved into 50-mL Falcon tubes, fixed with 2 mL of 37% formalin, and stored in the dark until processed. Suspensions were then mixed with 100 μL of 0.5% Triton X-100, and appropriate volumes of each sample were filtered through cellulose nitrate filters (25 mm diameter, 5 μm pore size; Sartorius Stedim Biotech GmbH, Goettingen, Germany). Filters were stained with 0.05% cotton blue in 60% lactic acid, and conidia were counted and identified under a microscope (200× magnification; Leica, DM1000, Wetzlar, Germany) [31]. Sporulation rates were expressed as the number of conidia released mg−1 litter DM day−1 and species richness as the number of species sample−1.
Litter Toughness
Litter toughness was determined for Al. glutinosa, Q. robur, and Ac. melanoxylon litter discs using a penetrometer [31]. Toughness was measured as the mass (± 0.01 g) required for a pin, from a 1.55-mm diameter iron rod, to penetrate through the litter, avoiding major veins [31]. Toughness was not determined for Ac. dealbata litter since it was not possible to apply this method to the small leaflets. Litter toughness was used as a surrogate for enzymatic maceration of litter and expressed as percentage of initial litter toughness. Initial litter toughness was determined applying the same method to litter discs extracted after the 1-week colonization period.
Litter Decomposition
The remaining 10 litter discs and portions of leaflets from each sacrificed microcosm were oven-dried at 105 °C for 24 h and weighed (± 0.1 mg). This DM was added to that of discs and portions of leaflets used for the determination of microbial respiration and fungal biomass to determine DM remaining. Litter DM remaining comprised the biomass of associated fungi. Results were expressed as percentage of initial DM. Initial DM was estimated by multiplying the initial wet mass by a conversion factor derived from an additional group of wet litter discs and leaflets from each stream. These samples were oven-dried at 105 °C for 24 h, weighed (± 0.1 mg) to determine the litter discs and leaflets initial DM, and the conversion factor was calculated as the ratio between the initial DM and the initial wet mass. Litter was incubated in the field for a short period, and it was carefully rinsed with distilled water before discs were cut or leaflets detached, and therefore, adherent sediments were not a concern and there was no need to estimate litter ash fraction.
Data Analysis
Data normality was checked by D’Agostino-Pearson or Kolmogorov-Smirnov test and homoscedasticity was checked by Bartlett or Levene’s test. When data failed to meet one of these assumptions, they were transformed (e.g., fungal biomass) before analysis.
Water characteristics were compared between stream types, and litter initial chemical characteristics were compared among species by one-way analysis of variance (ANOVA), followed by Tukey’s Honest Significant Difference (HSD) test when needed.
Decomposition rates (k, d−1) were calculated assuming an exponential decay model, by linear regression of the ln-transformed fraction of DM remaining against time (days), and a linear decay model, by linear regression of the fraction of DM remaining against time (days). Since none of these models properly fits the data in all treatments, the effects of litter species and stream type on litter decomposition, given by the DM remaining at the last sampling date, were tested by two-way ANOVA, followed by Tukey’s HSD test.
Fungal biomass (Box-Cox transformed to meet the assumptions of ANOVA), aquatic hyphomycete species richness and litter toughness were compared among litter species, stream type, and time (days) by three-way ANOVA, followed by Tukey’s test. Microbial respiration rates and fungal sporulation rates were compared among litter species, stream type, and time (days) by three-way univariate permutational analysis of variance (PERMANOVA), since data failed to meet the ANOVA assumptions of homoscedasticity even after transformation. Univariate PERMANOVAs were run on Euclidean distance [35], and residuals were permutated under a reduced model of 9999 permutations, followed by pair-wise comparison tests [35]. Univariate PERMANOVA is equivalent to a standard ANOVA and can be used when data fail to meet the assumptions of ANOVA [35]. For each of the measured parameters, the square of the correlation ratio (eta2), i.e., the percentage of the variance accounted for each of the main effects and interactions, was calculated as SSeffect/SStotal × 100 [36].
The structure of aquatic hyphomycete communities was assessed by cluster analysis, based on the Bray-Curtis similarity matrix of conidial production by aquatic hyphomycetes (log (x + 1) transformed). Aquatic hyphomycete communities were compared among litter species and between stream types by two-way multivariate PERMANOVA, followed by pair-wise comparison tests.
Cluster analysis, PERMANOVA and pair-wise tests were performed on PRIMER 6 (v6.1.16) and PERMANOVA+ (v1.0.6; Primer-E Ltd., Plymouth, UK). All other statistical analyses were performed on the STATISTICA 8 software (StatSoft Inc., Tulsa, Oklahoma, USA).
Results
Stream Water
During leaf colonization, stream water was cool, circumneutral, well-oxygenated, and had low conductivity and low nutrient concentrations (Table 1). Water characteristics were similar between stream types, except for conductivity and NO3−-N, which were significantly higher in “invaded” than in reference streams (one-way ANOVA, p < 0.001 and p = 0.003, respectively) (Table 1).
Litter Chemistry
Litter species differed in initial chemical characteristics (one-way ANOVA, p < 0.05), except for carbon concentration (Table 2). Quercus robur litter had the highest polyphenols and phosphorus concentrations and C:N ratio, and the lowest N concentration (Table 2). Acacia dealbata litter had the highest N concentration, higher lignin concentration than Q. robur, and the lowest polyphenol concentration and C:N ratio together with Al. glutinosa. Acacia melanoxylon litter had the lowest phosphorus concentration (Table 2).
Litter Decomposition
Litter mass decreased during the incubation period for all species (Fig. 2a–d). After 70 days, litter mass remaining varied between 52 and 50% for Al. glutinosa, 69 and 64% for Q. robur, 79 and 77% for Ac. melanoxylon, and 82 and 77% for Ac. dealbata, in reference and “invaded” streams, respectively. Litter mass remaining differed significantly among litter species and between stream types (two-way ANOVA, p < 0.001 and p = 0.021, respectively), with no significant interaction between factors (p = 0.631) (Table S1). Litter mass remaining was significantly higher in reference than in “invaded” streams (two-way ANOVA, p = 0.021) and was in the order: Al. glutinosa < Q. robur < (Ac. melanoxylon ~ Ac. dealbata) (Tukey’s test, p < 0.001). Litter species accounted for a disproportionally higher variance in litter mass remaining (eta2 = 92%) when compared with stream type (eta2 = 2%) (Table S1).

Dry mass remaining, litter toughness remaining and microbial respiration rate on Alnus glutinosa (a, e, h), Quercus robur (b, f, i), Acacia melanoxylon (c, g, j) litter discs and Acacia dealbata (d, k) leaflets incubated over 70 days in microcosms simulating conditions of reference and “invaded” streams. Values are means ± SE (n = 3)
Litter Toughness
Litter toughness remaining decreased mostly during the first 14 days of incubation for all species and after 70 days litter toughness varied between 25 and 21% for Al. glutinosa, 48 and 42% for Q. robur, and 55 and 60% for A. melanoxylon, in reference and “invaded” streams, respectively (Fig. 2e–g). Litter toughness differed significantly among litter species (three-way ANOVA, p < 0.001) (Table S2), with lower toughness for Al. glutinosa, followed by Q. robur and Ac. melanoxylon (Tukey’s test, p < 0.001). Litter species accounted for 59% of the variance in litter toughness (Table S2). Litter toughness did neither significantly differ between stream types nor was there a significant interaction between litter species and stream type (three-way ANOVA, p = 0.380 and p = 0.222, respectively) (Table S2).
Microbial Respiration Rate
After 70 days of incubation, microbial respiration rates varied between 0.40 and 0.39 mg O2 g−1 DM h−1 for Al. glutinosa litter, 0.16 and 0.26 mg O2 g−1 DM h−1 for Q. robur, 0.12 and 0.15 mg O2 g−1 DM h−1 for Ac. melanoxylon, and 0.48 and 0.55 mg O2 g−1 DM h−1 for Ac. dealbata, in reference and “invaded” streams, respectively (Fig. 2h–k). Microbial respiration rates differed significantly among litter species (three-way univariate PERMANOVA, p < 0.001) and were significantly affected by the interaction between litter species and stream type (p = 0.004) (Table S3). Microbial respiration was higher in Al. glutinosa litter, followed by Ac. dealbata and Q. robur, and lower in Ac. melanoxylon (pair-wise test, p < 0.002). Quercus robur, Ac. melanoxylon, and Ac. dealbata litter supported higher microbial respiration in “invaded” than in reference streams (pair-wise test, p = 0.013, p < 0.001, and p = 0.001, respectively). Litter species accounted for a higher variance in microbial respiration (eta2 = 46%) than the interaction between litter species and stream type (eta2 = 4%) (Table S3).
Fungal Biomass
After 70 days of incubation, fungal biomass varied between 32 and 30 mg g−1 DM for Al. glutinosa litter, 57 and 51 mg g−1 DM for Q. robur, 23 and 24 mg g−1 DM for Ac. melanoxylon, and 42 and 33 mg g−1 DM for Ac. dealbata, in reference and “invaded” streams, respectively (Fig. 3a–d). Fungal biomass differed significantly among litter species (three-way ANOVA, p < 0.001) and was significantly affected by the interaction between litter species and stream type (p < 0.001) (Table S2). Quercus robur and Ac. dealbata litter had higher fungal biomass than Al. glutinosa and A. melanoxylon (Tukey’s test, p < 0.001). Alnus glutinosa and Ac. dealbata litter had higher fungal biomass in reference streams, while Q. robur and Ac. melanoxylon had higher fungal biomass in “invaded” streams (Tukey’s test, p < 0.001). Litter species accounted for a higher variance in fungal biomass (eta2 = 54%) than the interaction between litter species and stream type (eta2 = 4%) (Table S2).

Fungal biomass, fungal sporulation rate and aquatic hyphomycete (AH) species richness associated with Alnus glutinosa (a, e, i), Quercus robur (b, f, j), Acacia melanoxylon (c, g, k) litter discs and Acacia dealbata (d, h, l) leaflets incubated over 70 days in microcosms simulating conditions of reference and “invaded” streams. Values are means ± SE (n = 3)
Fungal Sporulation Rate
Maximum sporulation rates by aquatic hyphomycetes varied between 517 and 378 conidia mg−1 DM day−1 for Al. glutinosa litter, 191 and 235 conidia mg−1 DM day−1 for Q. robur, 7 and 100 conidia mg−1 DM day−1 for Ac. melanoxylon, and 239 and 284 conidia mg−1 DM day−1 for Ac. dealbata, in reference and “invaded” streams, respectively (Fig. 3e–h). Sporulation rates differed significantly among litter species (three-way univariate PERMANOVA, p < 0.001) (Table S3). Generally, Al. glutinosa, Q. robur, and Ac. dealbata litter had higher fungal sporulation rates than Ac. melanoxylon, in both reference and “invaded” streams (pair-wise test, p < 0.050). Litter species accounted for 18% of the variance in sporulation rates (Table S3). Sporulation rates did neither differ significantly between stream types nor there was a significant interaction between litter species and stream type (three-way univariate PERMANOVA, p < 0.159 and p < 0.152, respectively) (Table S3).
Aquatic Hyphomycete Communities
Maximum aquatic hyphomycete species richness per date varied between 13 and 10 species for Al. glutinosa litter, 11 and 10 species for Q. robur, 8 and 5 species for Ac. melanoxylon, and 13 and 10 for Ac. dealbata, in reference and “invaded” streams, respectively (Fig. 3i–l). Species richness differed significantly among litter species and between stream types (three-way ANOVA, p < 0.001), with no significant interaction between factors (p = 0.658) (Table S2). Aquatic hyphomycete species richness over time was higher in reference than in “invaded” streams and higher for Al. glutinosa, Q. robur, and Ac. dealbata than for Ac. melanoxylon litter (Tukey’s test, p < 0.010). Litter species accounted for a higher variance in species richness (eta2 = 30%) than stream type (eta2 = 6%) (Table S2).
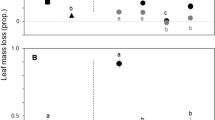
Over the entire incubation period, aquatic hyphomycete species richness varied between 23 and 16 species for Al. glutinosa litter, 20 and 21 species for Q. robur, 17 and 11 species for Ac. melanoxylon, and 22 and 20 for Ac. dealbata, in reference and “invaded” streams, respectively (Table S4). The aquatic hyphomycete community structure differed significantly among litter species and between stream types (two-way multivariate PERMANOVA, p < 0.001). Articulospora tetracladia Ingold and Triscelophorus acuminatus Nawawi were the most abundant species in Al. glutinosa, Q. robur, and Ac. dealbata litter, in both reference and “invaded” streams (Fig. 4a, b). In Ac. melanoxylon litter, Lunulospora curvula Ingold was abundant in both stream types, while A. tetracladia, Alatospora pulchella Marvavová, and Heliscus lugdunensis Sacc. & Terry were more abundant in reference streams and Dimorphospora foliicola Tubaki was more abundant in “invaded” streams (Fig. 4a, b). Therefore, aquatic hyphomycete communities were distributed in three groups: (i) Ac. melanoxylon litter in reference and “invaded” streams, (ii) Al. glutinosa, Q. robur, and Ac. dealbata litter in reference streams, and (iii) Al. glutinosa, Q. robur, and Ac. dealbata litter in “invaded” streams (pair-wise test, p < 0.050) (Fig. 5).

Relative contribution (%, based on conidial production) of aquatic hyphomycetes (AH) species associated with Al. glutinosa (Ag), Q. robur (Qr), and Ac. melanoxylon (Am) litter discs and Ac. dealbata (Ad) leaflets incubated over 70 days in microcosms simulating conditions of reference (a) and “invaded” (b) streams

Cluster dendrogram of aquatic hyphomycete communities associated with Alnus glutinosa (Ag), Quercus robur (Qr), and Acacia melanoxylon (Am) litter discs and Ac. dealbata (Ad) leaflets incubated over 70 days in microcosms simulating conditions of reference (R) and “invaded” (I) streams
Discussion
Invasion of native forests by N-fixing Acacia species occurs worldwide [19]. However, information about their effects on the structure and function of freshwater ecosystems is very limited (but see [37,38,39] for the effects of the invasion of the Fynbos biome by Acacia mearnsii De Wild in South African streams). In this study, litter decomposition, microbial decomposer activity, and aquatic hyphomycete species richness and community structure differed among litter species, probably due to differences in litter nutritional characteristics, and between stream types most likely due to differences in N concentration in stream water. Our results suggest that invasion of native riparian forests by Acacia species can affect litter decomposition and the activity, diversity, and structure of associated microbial decomposers in streams.
Decomposition of Acacia Litter Is Slower than that of Native Species
In this study, litter decomposition differed between native and invasive species. Litter decomposition was faster for Al. glutinosa, followed by Q. robur (native species) and slower for Ac. melanoxylon and Ac. dealbata (invasive species). This was expected since litter decomposition and microbial decomposer activity strongly depend on the structural and chemical characteristics of litter that fall into streams [40, 41]. Previous studies have shown that soft litter with high nutrient concentrations (e.g., N, P) and low concentrations of structural and secondary compounds (e.g., lignin, polyphenols) generally have higher microbial activity and faster decomposition than litter with high concentrations of recalcitrant compounds [9, 14, 42]. In this study, Al. glutinosa litter decomposed faster probably because it was soft, and had a low C:N ratio and a high N concentration, which facilitated microbial colonization and activity. Indeed, the activity of microbial decomposers was higher in Al. glutinosa litter when compared with the other species. On the other hand, in Q. robur litter, the high polyphenols and low N concentrations were most likely responsible for the limited microbial colonization and activity, and slower decomposition when compared with Al. glutinosa. Nevertheless, it was interesting to note that even though Q. robur litter was nutrient-poor for microbial decomposers, it decomposed faster than litter of invasive Acacia species probably because it was softer than Ac. melanoxylon phyllodes and had lower lignin concentration than Ac. dealbata leaflets. Litter of invasive N-fixing species, Ac. melanoxylon and Ac. dealbata, also had high N concentrations like Al. glutinosa; however, the higher concentration of recalcitrant structural compounds likely contributed to their decreased decomposition. Acacia melanoxylon litter decomposed slowly and had lower microbial activity, probably because it had a high lignin concentration and a tough and thick cuticle that acted as a physical barrier for microbial colonization [42]. Previous studies have shown that lignin and toughness are important factors limiting microbial decomposer activity and litter decomposition in streams [11, 42,43,44]. Therefore, Ac. melanoxylon can be considered a poor resource for aquatic microbial decomposers [42]. For Ac. dealbata, the reason for slow decomposition was not as clear as leaflets were chemically similar to Al. glutinosa litter and were fully colonized by microbes. One possible explanation is that leaflets probably had a tough and resistant cuticle that limited aquatic hyphomycete enzymatic activity on plant litter tissue, preventing the access of microbes to litter N. Therefore, the higher structural stability of leaflets given by their small size and higher lignin concentration probably allowed fungi to use the inorganic nutrients (e.g., N or P) directly from the stream water to grow mycelia in their surface and sporulate [45]. Indeed, previous studies have shown that higher microbial activity may occur in slow-decomposing substrates [10, 14, 46], probably due to their high structural stability for microbial growth. Additionally, the large surface area:volume ratio of Ac. dealbata leaflets may have facilitated its colonization by bacteria, which could have resulted in increased microbial activity but reduced litter decomposition due to the reduced capacity of bacteria to decompose litter [8, 47, 48].
Litter Decomposition Is Faster in “Invaded” than in Reference Streams
In this study, litter decomposition also differed between stream types. Litter decomposition was faster in “invaded” than in reference streams, probably due to the higher NO3−-N concentrations found in “invaded” stream water. Previous studies have shown that increases in N concentrations in water can accelerate litter decomposition in streams due to a stimulation of microbial decomposer activity [10, 13, 49, 50]. In our case, differences in decomposer activity between stream types were observed for microbial respiration (Q. robur, Ac. melanoxylon, and Ac. dealbata) and fungal biomass (Q. robur and Ac. melanoxylon), with higher values in “invaded” than in reference streams. This stimulation of microbial activity in “invaded” streams, although expected, is, nevertheless, impressive given that NO3−-N concentrations in “invaded” stream water were still in the oligotrophic range (< 30 μg L−1), despite being ~ 5× higher than in reference streams. Although we did not specifically measure the N concentrations in “invaded” stream water through time, the absence of agricultural activity in the catchment area of “invaded” streams and the absence of other sources of N enrichment suggest that the increased N concentration in “invaded” stream water was most likely due to N leaching from N-rich soils and the decomposition of N-rich litter that falls into streams [25, 28]. Indeed, increased N concentration has also been observed in streams running through forests invaded by other exotic N-fixing species [25,26,27,28,29]. We can therefore anticipate that the invasion of native forests by N-fixing Acacia species will most likely increase N concentration in stream water, and that this increase, even if small, will stimulate microbial decomposer activity [10, 49, 51, 52], especially in streams where background N is limiting, and thus, accelerate litter decomposition [10, 53, 54].
Aquatic Hyphomycete Community Structure Differed Among Litter Species and Between Stream Types
Aquatic hyphomycete species richness and community structure differed among litter species, given the differences in litter characteristics [55,56,57]. Aquatic hyphomycete communities on Ac. melanoxylon litter (17 and 11 species in reference and “invaded” streams, respectively) were highly distinct from those on Al. glutinosa, Q. robur, and Ac. dealbata, which were more diverse (20–23 and 16–21 in reference and “invaded” streams, respectively). It was also interesting to note that aquatic hyphomycete species richness and community structure differed between both Acacia species. Acacia dealbata litter supported higher species richness than Ac. melanoxylon and had a community structure very similar to that present on native Al. glutinosa and Q. robur. Thus, different Acacia species can have different effects on aquatic microbial decomposer communities. The invasion of native forests by species with characteristics similar to those of native species may have a weaker effect on stream decomposer communities than the invasion of native forests by species with very different characteristics [2]. However, the replacement of a diverse forest by a monospecific stand may still have strong negative impacts mediated by a reduction in resource diversity [15].
The species richness and structure of aquatic hyphomycete communities also differed between stream types. The total number of aquatic hyphomycete species (across litter species and sampling dates) recorded for reference and “invaded” streams was the same (24 species each), and very similar to the total number of species recorded in this study (25 species). However, the high difference between total and mean aquatic hyphomycete species richness per sampling date suggests high heterogeneity among streams; “invaded” streams had higher heterogeneity than reference streams (i.e., “invaded” streams have different aquatic hyphomycete communities with low number of species, while reference streams have similar communities with high numbers of species resulting in similar overall species richness per stream type). Aquatic hyphomycete communities from reference streams were more diverse than those from “invaded” streams on each sampling date, probably because the first had higher substrate diversity since they flow through diverse deciduous forests, while “invaded” streams flow through forests dominated by Acacia species, in some cases forming (almost) monospecific stands [15, 16, 58]. Higher aquatic hyphomycete species richness is expected to occur in streams flowing through diverse forests, due to higher substrate heterogeneity for aquatic hyphomycetes [15, 56]. Nevertheless, aquatic hyphomycete communities in A. glutinosa, Q. robur, and Ac. dealbata litter, from reference and “invaded” streams, were mostly dominated by A. tetracladia and T. acuminatus. Articulospora tetracladia is a species known to have a worldwide distribution [59] and can be found in streams with low and intermediate levels of nutrient concentrations [50]. Additionally, it was also interesting to note that invasive Ac. melanoxylon litter from reference streams supported a higher fungal species richness than litter from “invaded” streams, suggesting that the more diverse communities in reference streams may harbor fungal species with superior enzymatic capabilities (sampling effect mechanism, i.e., higher chance of having specific traits due to higher species richness [60]). In “invaded” streams, the aquatic hyphomycete community from A. melanoxylon litter was dominated by D. foliicola, a species known to be very abundant in streams with high levels of nutrient concentrations [50, 53]. Although our “invaded” streams were still in the oligotrophic range, the higher NO3−-N concentrations in water combined with the recalcitrant characteristics of A. melanoxylon stimulated D. foliicola conidia production.
Conclusion
Our results suggest that the invasion of native forests by exotic N-fixing Acacia species will affect litter decomposition and the activity and structure of associated microbial decomposers. However, the magnitude of the effects may vary with the identity of the Acacia species. Acacia melanoxylon and Ac. dealbata litter decomposed slower than the litter of the native species. However, although decomposer activity and aquatic hyphomycete species richness on Ac. melanoxylon litter were lower than on the native species, those on Ac. dealbata generally had values in the range of the native species. Also, differences in the structure of aquatic hyphomycete communities were found between native species and Ac. melanoxylon, but not Ac. dealbata. This suggests that the effects of tree species invasion on stream litter decomposition and microbial communities depend more on the litter characteristics of the species than on the species origin (native/exotic).
Additionally, the differences found between reference and “invaded” streams further suggest that effects of invasion by exotic N-fixing tree species on stream detrital pathways are mediated also by changes in water characteristics (e.g., increase in N concentration), although to a lower extent than by changes in litter characteristics. Overall, invasion of diverse deciduous forests by N-fixing Acacia species may affect litter decomposition via multiple pathways (e.g., changes in litter input characteristics, changes in water quality), with the magnitude of the effects depending on the dissimilarity between native and invasive species.
References
Millennium Ecosystem Assessment (2005) Ecosystems and human well-being: synthesis. Island Press, Washington DC
Kominoski JS, Shah JJF, Canhoto C, Fischer DG, Giling DP, González E, Griffiths NA, Larrañaga A, LeRoy CJ, Mineau MM, McElarney YR, Shirley SM, Swan CM, Tiegs SD (2013) Forecasting functional implications of global changes in riparian plant communities. Front Ecol Environ 11(8):423–432. https://doi.org/10.1890/120056
Castro-Díez P, Pauchard A, Traveset A, Vilà M (2016) Linking the impacts of plant invasion on community functional structure and ecosystem properties. J Veg Sci 27:1233–1242. https://doi.org/10.1111/jvs.12429
Ferreira V, Koricheva J, Pozo J, Graça MAS (2016) A meta-analysis on the effects of changes in the composition of native forests on litter decomposition in streams. For Ecol Manag 364:27–38. https://doi.org/10.1016/j.foreco.2016.01.002
Wallace JB, Eggert SL, Meyer JL, Webster JR (1997) Multiple trophic levels of a forest stream linked to terrestrial litter inputs. Science 277(5322):102–104. https://doi.org/10.1126/science.277.5322.102
Graça MAS, Canhoto C (2006) Leaf litter processing in low order streams. Limnetica 25(1–2):1–10
Gessner MO, Chauvet E, Dobson M (1999) A perspective on leaf litter breakdown in streams. Oikos 85:377–384. https://doi.org/10.2307/3546505
Baldy V, Gessner MO, Chauvet E (1995) Bacteria, fungi and the breakdown of leaf litter in a large river. Oikos 74:93–102. https://doi.org/10.2307/3545678
Gessner MO, Chauvet E (1994) Importance of stream microfungi in controlling breakdown rates of leaf litter. Ecology 75(6):1807–1817. https://doi.org/10.2307/1939639
Ferreira V, Gulis V, Graça MAS (2006) Whole-stream nitrate addition affects litter decomposition and associated fungi but not invertebrates. Oecologia 149:718–729. https://doi.org/10.1007/s00442-006-0478-0
Schindler MH, Gessner MO (2009) Functional leaf traits and biodiversity effects on litter decomposition in a stream. Ecology 90(6):1641–1649. https://doi.org/10.1890/08-1597.1
Ferreira V, Encalada AC, Graça MAS (2012) Effects of litter diversity on decomposition and biological colonization of submerged litter in temperate and tropical streams. Freshw Sci 31(3):945–962. https://doi.org/10.1899/11-062.1
Gulis V, Suberkropp K (2003) Effect of inorganic nutrients on relative contributions of fungi and bacteria to carbon flow from submerged decomposing leaf litter. Microb Ecol 45:11–19. https://doi.org/10.1007/s00248-002-1032-1
Pereira A, Geraldes P, Lima-Fernandes E, Fernandes I, Cássio F, Pascoal C (2016) Structural and functional measures of leaf-associated invertebrates and fungi as predictors of stream eutrophication. Ecol Indic 69:648–656. https://doi.org/10.1016/j.ecolind.2016.05.017
Ferreira V, Castela J, Rosa P, Tonin AM, Boyero L, Graça MAS (2016) Aquatic hyphomycetes, benthic macroinvertebrates and leaf litter decomposition in streams naturally differing in riparian vegetation. Aquat Ecol 50:711–725. https://doi.org/10.1007/s10452-016-9588-x
Laitung B, Chauvet E (2005) Vegetation diversity increases species richness of leaf-decaying fungal communities in woodland streams. Arch Hydrobiol 164(2):217–235. https://doi.org/10.1127/0003-9136/2005/0164-0217
Lecerf A, Dobson M, Dang CK, Chauvet E (2005) Riparian plant species loss alters trophic dynamics in detritus-based stream ecosystems. Oecologia 146:432–442. https://doi.org/10.1007/s00442-005-0212-3
Ghate SD, Sridhar KR (2017) Endophytic aquatic hyphomycetes in roots of riparian tree species of two Western Ghat streams. Symbiosis 71:233–240. https://doi.org/10.1007/s13199-016-0435-6
Richardson DM, Rejmánek M (2011) Trees and shrubs as invasive alien species – a global review. Divers Distrib 17:788–809. https://doi.org/10.1111/j.1472-4642.2011.00782.x
Souza-Alonso P, Rodríguez J, González L, Lorenzo P (2017) Here to stay. Recent advances and perspectives about Acacia invasion in Mediterranean areas. Ann For Sci 74:55. https://doi.org/10.1007/s13595-017-0651-0
Le Maitre DC, Gaertner M, Marchante E et al (2011) Impacts of invasive Australian acacias: implications for management and restoration. Divers Distrib 17:1015–1029. https://doi.org/10.1111/j.1472-4642.2011.00816.x
ICNF (2019) IFN6 – Principais resultados – relatório sumário. Instituto da Conservação da Natureza e das Florestas, Lisboa
Presidência do Conselho de Ministros (2019) Decreto-Lei n.° 92/2019. Diário da República 130:3428–3442
Sheppard AW, Shaw RH, Sforza R (2006) Top 20 environmental weeds for classical biological control in Europe: a review of opportunities, regulations and other barriers to adoption. Weed Res 46:93–117. https://doi.org/10.1111/j.1365-3180.2006.00497.x
Goldstein CL, Williard KWJ, Schoonover JE (2009) Impact of an invasive exotic species on stream nitrogen levels in southern Illinois. J Am Water Resour Assoc 45:3–672. https://doi.org/10.1111/j.1752-1688.2009.00314.x
Atwood TB, Wiegner TN, Turner JP, MacKenzie RA (2010) Potential effects of an invasive nitrogen-fixing tree on a Hawaiian stream food web. Pac Sci 64(3):367–379. https://doi.org/10.2984/64.3.367
Mineau MM, Baxter CV, Marcarelli AM (2011) A non-native riparian tree (Elaeagnus angustifolia) changes nutrient dynamics in streams. Ecosystems 14:353–365. https://doi.org/10.1007/s10021-011-9415-0
Wiegner TN, Hughes F, Shizuma LM, Bishaw DK, Manuel ME (2013) Impacts of an invasive N2-fixing tree on Hawaiian stream water quality. Biotropica 45(4):409–418. https://doi.org/10.1111/btp.12024
Stewart SD, Young MB, Harding JS, Horton TW (2019) Invasive nitrogen-fixing plant amplifies terrestrial–aquatic nutrient flow and alters ecosystem function. Ecosystems 22:587–601. https://doi.org/10.1007/s10021-018-0289-2
Graça MAS, Poquet JM (2014) Do climate and soil influence phenotypic variability in leaf litter, microbial decomposition and shredder consumption? Oecologia 174:1021–1032. https://doi.org/10.1007/s00442-013-2825-2
Graça MAS, Bärlocher F, Gessner MO (2005) Methods to study litter decomposition. A Practical Guide, Springer, Dordrecht
Goering HK, Van Soest PJ (1970) Forage fiber analyses (apparatus, reagents, procedures, and some applications). Agricultural Research Service, Washington DC
APHA (1995) Standard methods for the examination of water and watershed. American Public Health Association, Washington DC
Gessner MO, Chauvet E (1993) Ergosterol-to-biomass conversion factors for aquatic hyphomycetes. Appl Environ Microbiol 59(2):502–507
Anderson MJ, Gorley RN, Clarke KR (2008) PERMANOVA+ for PRIMER: guide to software and statistical methods. PRIMER-E, Plymouth
Richardson JTE (2011) Eta squared and partial eta squared as measures of effect size in educational research. Educ Res Rev 6:135–147. https://doi.org/10.1016/j.edurev.2010.12.001
Lowe SR, Woodford DJ, Impson DN, Day JA (2008) The impact of invasive fish and invasive riparian plants on the invertebrate fauna of the Rondegat River, Cape Floristic Region, South Africa. Afr J Aquat Sci 33(1):51–62. https://doi.org/10.2989/AJAS.2007.33.1.6.390
Railoun MZ (2018) Impacts of the invasive tree Acacia mearnsii on riparian and instream aquatic environments in the Cape Floristic Region, South Africa. Dissertation, Stellenbosch University
Wiener KD (2018) Quantifying the impacts of invasive N2-fixing plants on relative sediment loads and nutrient concentrations in mountain streams of the Cape Floristic Region, South Africa. Dissertations, Stellenbosch University
Gessner MO, Swan CM, Dang CK, McKie BG, Bardgett RD, Wall DH, Hättenschwiler S (2010) Diversity meets decomposition. Trends Ecol Evol 25(6):372–380. https://doi.org/10.1016/j.tree.2010.01.010
Zhang M, Cheng X, Geng Q, Shi Z, Luo Y, Xu X (2019) Leaf litter traits predominantly control litter decomposition in streams worldwide. Glob Ecol Biogeogr 28:1469–1486. https://doi.org/10.1111/geb.12966
Ferreira V, Raposeiro PM, Pereira A, Cruz AM, Costa AC, Graça MAS, Gonçalves V (2016) Leaf litter decomposition in remote oceanic island streams is driven by microbes and depends on litter quality and environmental conditions. Freshw Biol 61:783–799. https://doi.org/10.1111/fwb.12749
Li AOY, Ng LCY, Dudgeon D (2009) Effects of leaf toughness and nitrogen content on litter breakdown and macroinvertebrates in a tropical stream. Aquat Sci 71:80–93. https://doi.org/10.1007/s00027-008-8117-y
Jabiol J, Lecerf A, Lamothe S, Gessner MO, Chauvet E (2019) Litter quality modulates effects of dissolved nitrogen on leaf decomposition by stream microbial communities. Microb Ecol 77:959–966. https://doi.org/10.1007/s00248-019-01353-3
Suberkropp K, Chauvet E (1995) Regulation of leaf breakdown by fungi in streams: influences of water chemistry. Ecology 76(5):1433–1445. https://doi.org/10.2307/1938146
Arroita M, Aristi I, Flores L, Larrañaga A, Díez J, Mora J, Romaní AM, Elosegi A (2012) The use of wooden sticks to assess stream ecosystem functioning: comparison with leaf breakdown rates. Sci Total Environ 440:115–122. https://doi.org/10.1016/j.scitotenv.2012.07.090
Gulis V, Suberkropp K (2003) Interactions between stream fungi and bacteria associated with decomposing leaf litter at different levels of nutrient availability. Aquat Microb Ecol 30:149–157
Duarte S, Pascoal C, Alves A, Correia A, Cássio F (2010) Assessing the dynamic of microbial communities during leaf decomposition in a low-order stream by microscopic and molecular techniques. Microbiol Res 165:351–362. https://doi.org/10.1016/j.micres.2009.06.002
Gulis V, Ferreira V, Graça MAS (2006) Stimulation of leaf litter decomposition and associated fungi and invertebrates by moderate eutrophication: implications for stream assessment. Freshw Biol 51:1655–1669. https://doi.org/10.1111/j.1365-2427.2006.01615.x
Pereira A, Trabulo J, Fernandes I, Pascoal C, Cássio F, Duarte S (2017) Spring stimulates leaf decomposition in moderately eutrophic streams. Aquat Sci 79:197–207. https://doi.org/10.1007/s00027-016-0490-3
Huryn AD, Butz Huryn VM, Arbuckle CJ, Tsomides L (2002) Catchment land-use, macroinvertebrates and detritus processing in headwater streams: taxonomic richness versus function. Freshw Biol 47:401–415. https://doi.org/10.1046/j.1365-2427.2002.00812.x
Fernandes I, Seena S, Pascoal C, Cássio F (2014) Elevated temperature may intensify the positive effects of nutrients on microbial decomposition in streams. Freshw Biol 59:2390–2399. https://doi.org/10.1111/fwb.12445
Gulis V, Suberkropp K (2003) Leaf litter decomposition and microbial activity in nutrient-enriched and unaltered reaches of a headwater stream. Freshw Biol 48:123–134. https://doi.org/10.1046/j.1365-2427.2003.00985.x
Rosemond AD, Benstead JP, Bumpers PM, Gulis V, Kominoski JS, Manning DWP, Suberkropp K, Wallace JB (2015) Experimental nutrient additions accelerate terrestrial carbon loss from stream ecosystems. Science 347:1142–1145. https://doi.org/10.1126/science.aaa1958
Canhoto C, Graça MAS (1996) Decomposition of Eucalyptus globulus leaves and three native leaf species (Alnus glutinosa, Castanea sativa and Quercus faginea) in a Portuguese low order stream. Hydrobiologia 333:79–85. https://doi.org/10.1007/BF00017570
Gulis V (2001) Are there any substrate preferences in aquatic hyphomycetes? Mycol Res 105:1088–1093. https://doi.org/10.1016/S0953-7562(08)61971-1
Ferreira V, Elosegi A, Gulis V, Pozo J, Graça MAS (2006) Eucalyptus plantations affect fungal communities associated with leaf-litter decomposition in Iberian streams. Arch Hydrobiol 166(4):467–490. https://doi.org/10.1127/0003-9136/2006/0166-0467
Laitung B, Pretty JL, Chauvet E, Dobson M (2002) Response of aquatic hyphomycete communities to enhanced stream retention in areas impacted by commercial forestry. Freshw Biol 47:313–323. https://doi.org/10.1046/j.1365-2427.2002.00801.x
Duarte S, Bärlocher F, Pascoal C, Cássio F (2016) Biogeography of aquatic hyphomycetes: current knowledge and future perspectives. Fungal Ecol 19:169–181. https://doi.org/10.1016/j.funeco.2015.06.002
Dang CK, Chauvet E, Gessner MO (2005) Magnitude and variability of process rates in fungal diversity-litter decomposition relationships. Ecol Lett 8:1129–1137. https://doi.org/10.1111/j.1461-0248.2005.00815.x
Acknowledgments
We thank Olímpia Sobral (CEF, University of Coimbra) for all the help in the field and laboratory, and Teresa Gonçalves and Daniela Antunes (CNC, University of Coimbra) for the use of the lyophilizer. Water nutrient analyses were performed by CACTI - Centro de Apoio Científico-Tecnolóxico á Investigación from the University of Vigo, Spain.
Funding
This study was financed by the Portuguese Foundation for Science and Technology (FCT) through the strategic project UID/MAR/04292/2019 granted to MARE and through the exploratory project IF/00129/2014 granted to Verónica Ferreira. Ana Pereira received a doctoral fellowship from FCT (SFRH/BD/118069/2016), financed by the European Social Fund through the Human Capital Operational Program (POCH) and by National Funds from MCTES.
Author information
Authors and Affiliations
Corresponding author
Electronic Supplementary Material
ESM 1
(PDF 245 kb)
Rights and permissions
About this article

Cite this article
Pereira, A., Ferreira, V. Invasion of Native Riparian Forests by Acacia Species Affects In-Stream Litter Decomposition and Associated Microbial Decomposers. Microb Ecol 81, 14–25 (2021). https://doi.org/10.1007/s00248-020-01552-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00248-020-01552-3