
Abstract
The quest for novel natural products has recently focused on the marine environment as a source for novel microorganisms. Although isolation of marine-derived actinomycete strains is now common, understanding their distribution in the oceans and their adaptation to this environment can be helpful in the selection of isolates for further novel secondary metabolite discovery. This study explores the taxonomic diversity of marine-derived actinomycetes from distinct environments in the coastal areas of the Yucatan Peninsula and their adaptation to the marine environment as a first step towards novel natural product discovery. The use of simple ecological principles, for example, phylogenetic relatedness to previously characterized actinomycetes or seawater requirements for growth, to recognize isolates with adaptations to the ocean in an effort to select for marine-derived actinomycete to be used for further chemical studies. Marine microbial environments are an important source of novel bioactive natural products and, together with methods such as genome mining for detection of strains with biotechnological potential, ecological strategies can bring useful insights in the selection and identification of marine-derived actinomycetes for novel natural product discovery.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In recent years, the search for bioactive secondary metabolites has turned its focus to the exploration of microbial populations from ecologically diverse environments that harbor microorganisms with specialized adaptations. This has yielded many novel natural products with diverse types of bioactivity, including novel diketopiperazine dimers and a new iso-naseseazine B, novel isocoumarins, streptorubin B, sporolides, and marinomycins [1,2,3]. The marine environment has become an important source of income for this industry with estimations on the global market for marine-derived drugs reaching up to $ 8.6 billion a year [4, 5]. Marine-derived Actinobacteria, found mainly in marine sediments, are still the leading source of novel compounds and have been responsible for compounds such as the abyssomicins, salinosporamide A, and arenicolides [6,7,8].
Even with improved cultivation techniques, only less than 20% of microbial diversity can generally be accessed under laboratory conditions [9, 10] and only a few of the cultivated taxa (i.e., 7000) have been formerly described with a type strain [10, 11]. The opportunities of finding new microbes by modifying current cultivation techniques are vast. One way to search for uncultured diversity may be the exploration of environments with distinct environmental pressures, such as the ocean floor. The ocean sediments cover three fourths of our planet and are subjected to high pressures, varying oxygen conditions, and the influence of ions present in seawater. These conditions could provide distinct selective pressures to microorganisms resulting in the production of new molecules [12, 13]. In fact, over the past 12 years, reports on the discovery of marine natural products with pharmaceutical significance have continued to increase [5, 14] and new technologies that allow for genome mining to explore cryptic biosynthetic gene clusters have become a very promising area for natural product discovery [15]. However, exploration of marine bacterial diversity often requires cultivation strategies for specific bacteria with potential for secondary metabolite production that are of utmost importance in natural product discovery. Actinomycetes are widespread in the ocean environment with more than 17 genera isolated from diverse sediments and up to 10% of marine snow microbial populations represented by this class of bacteria [13, 16,17,18]. Actinomycetes isolated from the Sea of Cortez to the deep trenches in the middle of the Atlantic suggest they have managed to adapt and survive under diverse and harsh conditions [16, 17, 19, 20]. The habitats in which marine-derived actinomycetes thrive suggest they have developed a distinct set of chemical repertoires for defense and microbial communication [21, 22].
Understanding the taxonomic distribution and marine adaptations of these microorganisms in easily accessible coastal areas could help lead to the identification of locations with taxonomically diverse populations of marine-adapted actinomycetes, therefore increasing the chances of finding novel compounds. In the recent past, exploration of the marine environments as a source of actinomycetes has resulted in the discovery of bioactive metabolites including meroterpenoids, like azamerone, sesquiterpenes like neomarinone, polyketides like saliniketal and abyssomicin C, and peptides like thiocoraline and cyclomarin A [23]. The exploration of overlooked marine environments, including coastal sediments of the Yucatan Peninsula where karstic sediments and bottom water spurts from the underground aquifer join [24]], could lead to the discovery of new microbial taxa capable of producing novel compounds. In fact, recent studies have shown that exploration of coastal zones is still a promising source of bioactive actinomycetes [16, 25,26,27,28,29].
Even though the use of novel sequencing technologies as a way to mine for specialized metabolite biosynthetic pathways in Actinobacteria has recently bolted [30], random selection of locations for cultivation of actinomycetes from the marine environment has still resulted in successful achievements in the discovery of sources for bioactive compounds [16]. However, this randomness can be reduced by applying ecological strategies towards the selection of locations with increased probability of harboring microbial taxa that produce novel metabolites [22]. The present study was designed to explore the diversity of marine-derived actinomycetes from sites along the Yucatan coast and use ecological and phylogenetic strategies to support the hypothesis that distinct phylogeny and marine adaptations can be used as ecological indicators of strains that produce bioactive secondary metabolites.
Materials and Methods
One hundred six marine sediment samples were collected between 2 and 30 m depth by scuba diving and using 50-mL Whirl Pack sterile bags from 12 sites located in the Yucatan Peninsula from January 2012 to March 2013. Samples were kept at 4 °C until processed.
Marine sediment samples (5 g) were dried in a laminar flow hood (24 to 36 h); once completely dried, the sediment was stamped in agar plates using an autoclaved foam plug in a clockwise direction to create a serial dilution effect as described previously by Jensen et al. [31]. Agar plates contained A1 media with peptone (4 g), starch (10 g), yeast extract (2 g), cycloheximide (100 μg mL−1) to prevent fungal contamination, and either rifamycin or gentamicin (10 μg mL−1 and 5 μg mL−1, respectively) to reduce the growth of gram-negative bacteria. Plates were incubated at 28 °C for 1 to 3 weeks. Colonies were selected based on morphology and pigmentation as mentioned in Bergey’s Manual of Systematic Bacteriology [32]. Strains were subcultured using A1 media until pure and tested using a non-staining reaction with KOH [33]. Gram-positive isolates were then inoculated in 2-L flasks of A1 liquid medium in order to obtain cell mass.
Molecular Methods
Genomic DNA from the isolates was extracted using the in situ lysis method described by Rojas and collaborators [34]. Presence of DNA was confirmed by electrophoresis on an agarose gel (1%). The 16S rDNA gene was amplified using the actinomycete-specific primers FC27 (5′ TACGGCTACCTTGTTACGACTT 3′) and RC1492 (5′ AGAGTTTGATCCTGGCTCAG 3′) and the program suggested by Mincer and collaborators [20] was followed: 10 min at 95 °C, followed by 30 cycles of 45 s at 94 °C, 45 s at 65 °C for annealing and 1 min at 72 °C for extension, and a final extension for 10 min at 72 °C as suggested. PCR product amplification was confirmed by agarose gel electrophoresis.
PCR products were sent to Macrogen enterprise (Korea) for purification and sequencing. All sequences were deposited in the GenBank data base under accession numbers KT214470–KT214553.
Sequence Processing
Reverse and forward sequences were aligned and trimmed (900 bp) using Geneious 7.1.2 (http://www.geneious.com) and nearest neighbors were obtained by blasting against the EZTaxon database [35]. All further sequence alignments were performed in SILVA ( www.arb-silva.de/act ) and operational taxonomic units (OTUs) at 97% sequence identity were created using the cluster command and average neighbor method on MOTHUR [36, 37]. Using the otu.rep command in MOTHUR, one representative sequence per OTU was used to create an alignment. This was used to create a maximum likelihood phylogenetic tree with 1000 bootstrap randomizations on MEGA 7 [38, 39]. A representative OTU tree was used as an input for the weighted.unifrac command in MOTHUR to obtain a dissimilarity dendogram and the UniFrac values among locations. The richness estimator Chao 1 [40] was calculated using the rearefaction.single command in MOTHUR. All graphics were developed on the R project software [41, 42] using vegan. All edits on names for the phylogenetic tree were done using InkScape (http://www.inkscape.org).
Effect of Distilled Water (DW) on Growth
To test the requirement of seawater for growth, all strains were inoculated on A1 media. Once growth was observed, strains were inoculated at the same time on plates containing A1 medium prepared with seawater and in plates with the same medium using distilled water. Plates were incubated at 28 °C for 1 to 3 weeks. Full growth was considered when the actinomycete strain grew equally well in both media after the time period evaluated. Isolated growth was determined when the actinomycete strain showed random growth of colonies on DI water medium while the area of growth was full of biomass on the seawater water medium after the time period evaluated. Null growth was considered when no colonies where observed on the DI water medium while the area of growth was full of biomass on the seawater medium after the time period evaluated.
Cultivation and Extraction of Marine Actinomycetes
The strains NCA002, NCA004, NCA005, NCA006, NCA008, NCA012, NCA024, NCA049, and NCA093 were grown for 10–12 days in Erlenmeyer flasks (two 2 L) containing seawater A1 medium (1 L) and placed on a rotary shaker (120 rpm, 27 °C). Following fermentation, each strain culture was centrifuged (5000 rpm, 30 min) and the cell-free supernatant was harvested and mixed twice with ethyl acetate (600 mL). The organic layer was dried using anhydrous sodium sulfate and the solvent was evaporated under reduced pressure. The cell pellets were washed with distilled water and extracted with acetone (500 mL) two times at room temperature. The acetone was evaporated under reduced pressure.
Thin-Layer Chromatography
Analytical thin-layer chromatography (TLC) was carried out using aluminum-backed silica gel (60F254) plates (E.M. Merck, 0.2 mm thickness); the plates were first examined under UV light (254 and 366 nm) and visualized by dipping the plates in a solution of phosphomolybdic acid (20 g) and ceric sulfate (2.5 g) in 500 mL of sulfuric acid (5%), followed by drying and gentle heating. The metabolites present in the ethyl acetate extracts were then visualized using dichloromethane/methanol 95:5. The acetone extracts were visualized using dichloromethane/methanol 6:4.
Gas Chromatography-Mass Spectrometry
The gas chromatography-mass spectrometry (GC-MS) analysis was carried out using a Trace GC Ultra Series GC System (ThermoScientific Inc., Waltham, MA, USA) coupling with a mass detector triple quadrupole model TSQ Quantum XLS (ThermoScientific Inc.). A ThermoScientific TR-5MS column (30 m, × 0.25 mm i.d., × 0.25 μm film thickness) was used with helium as a carrier gas (1.2 mL min−1). The GC oven temperature was kept at 50 °C for 1 min and programmed to 300 °C at a rate of 7 °C min−1 for 2 min. The injector temperature was at 50 °C.
Liquid Chromatography Analysis
The crude extracts were analyzed by ultra-performance liquid chromatography tandem mass spectrometry (UPLC-MS). The UPLC system, a Waters Acquity UPLC-H Class (Waters, Milford, MA, USA) equipped with a quaternary pump, sample manager, column oven, and photodiode array detector (PDA), was coupled to a SQD2 single-quadrupole mass spectrometer with an electrospray ionization (ESI) source. The analysis were performed using a reversed-phase C18 column (Waters BEH C18 column, 50 × 2.1 mm i.d., 1.7  m) and a gradient solvent system from 20:80 to 100:0 CH3CN-H2O (0.1% formic acid) in 12 min, at a flow rate of 0.3 mL min−1. PDA detector was set from 190 to 500 nm and the mass spectrometer parameters were the following: cone and capillary voltage, 35.0 V and 3.0 kV, respectively; source and desolvation temperature, 350 and 450 °C, respectively; collision gas, N2; and mass range, m/z 50–2000 (scan duration of 0.5 s). All samples were analyzed in both positive and negative modes.
m) and a gradient solvent system from 20:80 to 100:0 CH3CN-H2O (0.1% formic acid) in 12 min, at a flow rate of 0.3 mL min−1. PDA detector was set from 190 to 500 nm and the mass spectrometer parameters were the following: cone and capillary voltage, 35.0 V and 3.0 kV, respectively; source and desolvation temperature, 350 and 450 °C, respectively; collision gas, N2; and mass range, m/z 50–2000 (scan duration of 0.5 s). All samples were analyzed in both positive and negative modes.
Purification of NCA004 and NCA008 and the Isolation of Resistomycin
The crude ethyl acetate extract of NCA004 (200 mg) was subjected to gravity column chromatography purification (2.5 cm diameter, 20 cm high) with Hx/CHCl3/AcOEt/MeOH 7:3:2:1 to produce five main fractions (A–E). Fraction C was determined by TLC as a pure metabolite 1 (2 mg) yellow oil soluble in CHCl3, retention factor (Rf) 0.63 in Hx/AcOEt 95:5, and identified by high-performance liquid chromatography-mass spectrometry (HPLC-MS) as resistomycin. Additionally, sonication of the crude acetone extract of strain NCA004 (2.05 g) with 100 mL of CH2Cl2 for 30 min (2x) yielded 460.3 mg of a medium-to-low polarity fraction, which was then subjected to successive vacuum liquid chromatography (VLC) (CHCl3/MeOH), gravity column chromatography (Hx/An 8.2, 7:3, and 1:1), and gel permeation column chromatography, Sephadex LH-20 (CH2Cl2/MeOH 1:1), that resulted in seven semi-purified fractions. The sonication of the crude acetone extract of strain NCA008 (2.56 g) with 100 mL of CH2Cl2 for 30 min (2x) yielded 693.7 mg of a medium-to-low polarity fraction, which was then subjected to a gel permeation column chromatography, Sephadex LH-20 (CH2Cl2/MeOH 1:1), and resulted in two main semi-purified fractions.
Antimicrobial Activity of Crude Fermentation Extracts
Antimicrobial activity from the ethyl acetate and acetone extracts from nine marine actinomycete strains was determined by a triplicate bioautography assay (planar chromatographic analysis-biologial detection method [43] with the pathogenic bacterial strains Staphylococcus aureus (ATCC 25923) and Pseudomonas aeruginosa (ATCC 27853)). This method is a fast and simple technique which combines the advantages of TLC separation of a crude extract and antimicrobial detection [44, 45]. Activity of the spots with different Rf from crude extracts was assessed by adding 0.5 mg of crude extracts (or semi-purified fraction extracts) on the TLC plates and eluted using a dichloromethane/methanol 95:5 mixture. After the elution, each TLC plate was set in 10 mL TSB medium containing 1 × 106 UFC mL−1 of bacteria. TLC plates were incubated at 37 °C for 24 h. To aid inhibition zone visualization, dehydrogenase activity was detected with tetrazolium salt [3-(4, 5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, MTT] reagents (1 mg mL−1) since metabolically active bacteria convert MTT into formazan dye [43]. A strong bioactivity was considered when halos of inhibition around the spots in the TLC covered more than 50% of the growth in the plate. A medium bioactivity was considered when halos of inhibition around the spots in the TLC covered between 10 and 50% of the growth in the plate. A weak bioactivity was considered when halos of inhibition around the spots in the TLC were small and covered less than 10% of the plate.
Results
Molecular Identification and Operational Taxonomic Unit Distribution of Marine Sediment-Associated Actinomycetes
From a total of 106 sediment samples, 85 actinomycete strains were isolated and taxonomically identified. Thirty-two operational taxonomic units (OTUs) were formed based on 97% similarity within these sequences. The OTUs were distributed among nine different genera: Streptomyces, Saccharomonospora, Dietzia, Nocardiopsis, Pseudonocardia, Verrucosispora, Brachybacterium, Jiangella, and Salinispora.
Most (61%) of OTUs belonged to the Streptomyces genus which were also the most abundant of the genera identified (Table 1). Salinispora was the second most abundant genus with four OTUs followed by Jiangella, with three OTUs. The rest of the genera had one or two OTUs formed by single strains (e.g., Saccharomonospora, Dietzia, and Brachybacterium) or by multiple strains in the same OTUs (Nocardiopsis, Pseudonocardia, and Verrucosispora).
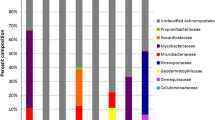
From all 12 locations covering the western [Gulf of Mexico (Celestún)], northern [Yucatan current (La Bocana, Chelem, Dzilam, Rio Lagartos, and Holbox)], and eastern [Caribbean (Cancun, Puerto Morelos, Cozumel, Akumal, Mahahual, and Banco Chinchorro)] coasts of the Yucatan Peninsula, the highest number of genera and OTUs was observed in Cancun (Fig. 1), while in Akumal and La Bocana, only Streptomyces and Salinispora were identified. The only location where no representative of the Streptomyces genus was isolated was Banco Chinchorro which is an island in the Caribbean seas (Fig. 1).

Number of OTUs and genera from sampled locations
The phylogenetic diversity of marine-derived actinomycetes isolated from the Yucatan shows that strains in OTUs belonging to the genera Dietzia, Pseudonocardia, Nocardiopsis, Verrucosispora, and Saccharomonospora are similar to their nearest type strain, while many Streptomyces strains branch out from the type strain “leaf” (Fig. 2, Supplementary Figure 1). In an effort to explore if these branching Streptomyces OTUs showed an adaptation to life in the ocean, we ran growth tests in deionized water media. Out of 69 Streptomyces strains tested, 21 showed a requirement for growth. Figure 2 shows OTUs that contain seawater requiring strains in bold. When their 16S rRNA gene sequence was compared to the GenBank database, all strains shared more than 98% of their nucleotide sequence with their nearest neighbor; therefore, we did not consider any of them as novel actinomycete strains. However, all of these strains’ nearest neighbors were reported as isolated from marine environments (data not shown); therefore, all of our isolates can still be considered as marine-derived.

Maximum likelihood tree with 1000 bootstrap randomizations of partial 16S rDNA gene sequences (900 bp) from OTUs isolated from marine sediments of the Yucatan Peninsula. Nodes above 50% bootstrap values are shown. OTU representative strains display collection number, Genbank accession number, and number of sequences per OTU in parenthesis. OTUs with strains that were unable to grow in distilled water are shown in bold. Black circles indicate OTUs with a bioactive strain (see results below). Type strains show the ID numbers found on EZ-Taxon (ezbiocloud.net)
Alpha Diversity of Marine Sediment-Derived Cultivable Actinomycetes in Yucatan
A rarefaction curve for the count of the actinomycete OTUs in Yucatan, together with a richness estimator, shows the number of OTUs does not reach a plateau, suggesting our sampling effort was not enough to reach the highest number of culturable actinomycetes from sampled sediments. Therefore, further processing and cultivation methods, or a greater sampling effort in these locations, is recommended (Fig. 3). A more detailed analysis shows that no location was sampled or processed to its full potential for actinomycete cultivation (Supplementary Figure S2).

Rarefaction curves for richness (sobs) and estimated richness (chao) using the statistical estimator Chao 1 [46] of cultivable actinomycetes. Data shows a plateau was not reached, suggesting further sampling or processing of samples is needed to achieve a higher coverage of the cultivable actinomycete richness from sediments in Yucatan
Effect of Seawater on Growth of Marine Sediment-Associated Actinomycetes
A growth test where the seawater in the medium was replaced with deionized water assessed adaptation to growth in the marine environment. Our results suggest that marine adaptation is a common trait among coastal streptomycetes since 30% of strains isolated from these areas required seawater for growth. Furthermore, the acquisition of this trait seems to be common (40%) among diverse Streptomyces from coastal sediments from the Yucatan (Fig. 2). On the other hand, non-Streptomyces strains isolated from these same sediments did not require seawater for growth, except for the obligate marine genus Salinispora. This result suggests that Streptomyces have either been in the coastal area for longer times allowing them to adapt to these conditions or that members of these genus have a greater genomic plasticity than other actinomycetes which enables them to easily adapt to stressing environmental conditions [47]. The two locations with the highest number of marine-adapted Streptomyces were Chelem and Mahahual, while none of the Streptomyces strains isolated from Celestún, Dzilam, and Cancun showed any effects when seawater was removed from the growth medium (Fig. 4) highlighting the former two locations as key sites for future sampling efforts.

Distribution of Streptomyces strains according to their growth on DW among sampled locations (full growth: no changes in growth on both media; isolated growth: random growth in DW medium; null growth: no growth on DW medium)
Beta Diversity of Marine Sediment-Derived Cultivable Actinomycetes in Yucatan
To explore the phylogenetic differences between cultivable actinomycete diversity in marine sediments from different locations of the Yucatan, we performed a UniFrac statistical test which uses phylogenetic information (i.e., evolutionary distances) to compare microbial communities [48]. Two distinct clusters were observed: the first cluster contained locations where a majority of the cultivable strains belonged to the family Micromonosporacea (Rio Lagartos, Cancun, Dzilam, Akumal, and Banco Chinchorro); and the second cluster was formed by locations where most strains belonged to the family Streptomycetacea (Mahahual, Holbox, Chelem, La Bocana, Cozumel, and Celestún) (Fig. 5). Internal nodes in these clusters (sub-clusters) suggest that geographic vicinity is not a determining factor for phylogenetic similarity among cultivable actinomycete diversity. That is, locations as far apart as Rio Lagartos (northern Yucatan—Yucatan current) and Cancun (eastern Yucatan—Caribbean Sea), or Dzilam and Akumal, clade closer together than to any adjacent location. No trends were observed according to sample locations, except for Banco Chinchorro and Celestún that did not form a cluster with any other location. The former is an island in the Caribbean eastern side of the peninsula, while the latter is a coastal lagoon on the western side of the peninsula.

Weighted UniFrac dissimilarity dendogram shows a clear separation between Micromonosporacea-dominated locations (top clade) and those locations dominated by Streptomycetacea (bottom clade) in the Yucatan Peninsula. Banco Chinchorro (island in the eastern Caribbean) and Celestún (coastal lagoon in the western Gulf of Mexico side of the peninsula) do not group with any other location
Antimicrobial Activity of Fermentation Extracts and Compound Isolation
Based on the phylogeny, location, and seawater requirement analysis, eight streptomycetes were selected (Table 2). The ethyl acetate and acetone crude extracts of eight strains were tested for antimicrobial activity against Staphylococcus aureus and Pseudomonas aeruginosa by a bioautography assay in order to analyze the activity of TLC spots of the crude extracts. All ethyl acetate extracts from Streptomyces strains from Chelem showed strong bioactivity (Table 3). Bioactivity from the ethyl acetate extract of NCA024 (seawater requiring strain from a location other than Chelem) was weak, while NCA049 (non–seawater-requiring strain from Cozumel) showed no bioactivity on the assays. Extracts from NCA005 (non–seawater-requiring strain from Chelem) also showed high bioactivity (see “Materials and Methods”), suggesting that Chelem is a good location to undertake broader sampling efforts. Based on these results, NCA004 and NCA008, both seawater-requiring strains from Chelem, were selected for further chemical analyses.
Successive chromatographic purifications of the ethyl acetate extract of strain NCA004 showed high activity of the semi-purified fractions (Supplementary Figure S3) and resulted in the isolation of metabolite 1, which was identified by liquid chromatography and mass spectroscopy (UPLC-MS) as resistomycin, a quinone-related antibiotic from the family of phenanthrenes and derivatives that possesses bactericidal and vasoconstrictive activity [26]. The resistomycin peak appeared at 12.057 min in the chromatogram (Supplementary Figure S4). The UV profile and MS data were both consistent with resistomycin (Supplementary Figure S5). The parent ion peak at m/z (M-H) found for C22H15O6 was 375.7.
Additionally, a dichloromethane fraction of the acetone extract from strain NCA004 was further fractionated and seven main semi-purified fractions were analyzed by GC-MS (Table 4). Retention time and molecular weight corresponding to the following secondary metabolites were observed: carboxylic acids (13-octadecenoic acid, tetradecanoic acid, pentadecanoic acid, 13-octadecenoic acid, 1,3-benzenedicarboxylic acid mono 2-ethylhexyl ester), hydrocarbons (9-eicosene, 10-eicosene, decane, undecane, dodecane), amides (9-octadecenamide), esters (butoxyethoxyethyl acetate, terephthalic acid ester derivate), and amines (piperazine and pyrrolol derivates).
Discussion
Marine actinomycetes have long been known as a promising source of novel metabolites with biological activity [6, 27,28,29,30]. Nevertheless, successful selection of strains producing novel bioactive compounds is still a challenge. One of the main obstacles for natural product discovery from marine sources lies in the number of isolates that can be recovered. Even though selective isolation methods have allowed us to cultivate a considerable amount of bacterial phyla, the number of bacteria that can be brought to pure culture remains around the 1% range [49]. Marine actinomycete cultivation techniques allow us to obtain up to hundreds of strains in a plate [14]; however, selection of strains for chemical and bioactivity analyses is usually a random process. Cultivation-dependent methods are still considered as a useful method bioprospecting, especially as they allow further strain manipulation for compound isolation [32]. Novel technologies like matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS), mass spectrometry-based molecular networking, and genome mining MS-guided metabolomics [50] have been suggested as a good way to prioritize strain selection in natural product discovery [33]. However, many of these methods are costly. Here, we present a study that focuses on the use of accessible microbial ecology techniques as a tool to increase the selective isolation of bioactive actinomycete strains cultivated from the Yucatan Peninsula.
The coastal marine sediments of the Yucatan Peninsula harbor a notable diversity of cultivable actinomycetes (Table 1), with nine different genera well represented among 85 strains from 11 locations (Fig. 1). Other studies of Mexican coastal environments have reported the isolation of 300 to 1497 strains; however, 17 culture media were used [35]. In this case, the use of one selective medium amended with three antibiotics applied to 101 samples resulted in the isolation of three actinomycete genera not reported yet from Mexican coastal sediments: Jiangella, Pseudonocardia, and Brachybacterium, and four others that have been previously reported from the Gulf of Mexico and Gulf of California: Streptomyces, Salinispora, Verrucosispora, and Nocardiopsis [35,36,37,41]. Among this diversity, Streptomyces was the most dominant genus (Fig. 1). This is not surprising as this genus is often isolated from marine sediment samples using selective media [43, 44]. Furthermore, the use of nitrogen compounds in culture media has been reported to favor the growth of antibiotic-producing strains, like Streptomyces [47]. This over-representation of the Streptomyces can be reduced when samples are recovered further from shore, even when the same culture media are used [48, 51]. The only location where no Streptomyces strains were isolated was Banco Chinchorro, while Cancun, Dzilam, and Rio Lagartos showed a higher number of distinct genera reducing the overall Streptomyces dominance. The presence of four Salinispora OTUs may be misleading regarding the microdiversity in this genus which has been shown to share over 99% of its 16S rRNA variance among species [52]. However, careful examination of the sequences show that most of these differences were mostly due to clear sequencing errors in the Salinispora chromatograms that were detected when the 2X full sequence was carefully re-examined. The presence of S. arenicola and S. tropica strains in these geographic locations could lead to the discovery of different chemical structures from the secondary metabolite families they produce since studies have shown that phylogenetically related strains collected from diverse locations produce distinct secondary metabolites [52]. Although several OTUs (31) were recovered from the 12 locations and 101 samples, the rarefaction curve did not reach saturation, and taken together with the Chao 1 richness estimator, it is clear that further sampling or processing of samples would yield more OTUs (Fig. 2) [51].
All isolates were tested for seawater requirements and Salinispora and Streptomyces were the only two genera with strains that showed a clear dependency on it for growth. Locations on the North and East of the Yucatan Peninsula had from one to seven seawater-requiring Streptomyces strains and Celestún, Dzilam, and Cancun each were the only locations without Streptomyces with some requirement of seawater for growth (Fig. 4). The highest percentage of seawater-requiring Streptomyces strains occurred in Chelem, followed by Mahahual, La Bocana, and Akumal. According to Aranda-Cirerol [53], Chelem and La Bocana are both found in a similar hydrochemical area, but the coastal area in Chelem has a higher wastewater discharge [54] and its salinity values range from 36 to 40 ppt [55]. In spite of being constantly under terrestrial influence, most of the strains in Chelem displayed strict marine attributes, with their closest relatives also isolated from the marine environment (data not shown), while many of the strains isolated from La Bocana show slow growth without seawater, suggesting they may be in a process of adaptation to marine conditions. The high number of strains with some degree of requirement of seawater for growth found in these coastal sediments could be due to the high tolerance of salt reported for this genus (up to 14%) [56] and to the evolutionary adaptations to the marine environment that have been described and which would allow terrestrial strains to reach the ocean (33 ppt of salt) and colonize a new niche and eventually become a different taxon with a dependency on seawater for their growth [57]. Recent genome sequencing of marine-derived Streptomyces has shown a potential for synthesizing novel antimicrobial peptides [58] as well as prenyltransferases involved in the synthesis of isoprenoids [59]. Furthermore, recent statistical studies on the diversity of marine natural products by Pye and collaborators have shown that compounds isolated from novel marine sources have a higher chance of differing from the usual chemical scaffolds [9]. These authors highlight the importance of designing new biological prioritization methods to improve the effective selection of these sources [9]. The production of interesting and novel chemical compounds from strictly marine strains [60, 61] highlights the importance of generating an ecological perspective from the isolates including their requirement of seawater for growth, their similarity, or lack of it, to terrestrial strains, their chemical interactions, and their resulting biological activities to select for novel natural product production.
Phylogenetic analysis shows the presence of nine different genera of cultivable marine actinomycetes within the Yucatan Peninsula’s coastal sediments (Fig. 4). All partially sequenced strains shared > 98% of their partial 16S rRNA gene sequences with reported strains in public databases and were therefore not considered novel; however, the requirement of seawater for growth in some Streptomyces strains could suggest marine adaptations of novel ecotypes. Comparative genomics of marine-derived Streptomyces has evidenced adaptations on solute transport and nutrient acquisition mechanisms revealing their genetic adaptation to the marine environment [57]. Therefore, it is not surprising that many of the strains isolated in this study make phylogenetically distinct clades when compared to their nearest reported neighbors.
Although the environmental characteristics between sediments from the Yucatan and the Caribbean differ greatly, i.e., submerged groundwater discharges vs coral reef ecosystem [54, 62], a UniFrac analysis shows no influence of sample location on the phylogenetic diversity of cultivated actinomycetes. Instead, a clear separation between Streptomycetaceae and Micromonosporaceae is observed (Fig. 5). Preliminary chemical analyses on the sediments show no relationship between C or N content and the UniFrac results (data not shown), but further analyses could determine if other physico-chemical and biological features of those habitats select for specific actinomycete genera [63].
To our knowledge, this is the first study that shows that simple ecological features can be used as signs of marine adaptation from Streptomyces strains and can be taken as criteria for the selection of strains for natural product discovery. This approach has proven useful in our search for bioactive actinomycetes from the Yucatan. These simple and non-expensive methods can be the first step to designing a more robust and innovative discovery strategy involving bioprospecting of overlooked marine environments. Furthermore, together with genomic and metagenomic exploration of the microorganisms potential for natural product biosynthesis and dereplication strategies like the Global Natural Product Social Molecular Networking platform or MALDI-TOF MS, the rate of natural product discovery should be notably increased.
References
Kong DX, Jiang YY, Zhang HY (2010) Marine natural products as sources of novel scaffolds: achievement and concern. Drug Discov Today 15(21–22):884–886. https://doi.org/10.1016/j.drudis.2010.09.002.
Mayer AM, Glaser KB, Cuevas C, Jacobs RS, Kem W, Little RD, McIntosh JM, Newman DJ, Potts BC, D S (2010) The odyssey of marine pharmaceuticals: a current pipeline perspective. - PubMed - NCBI. Trends Pharmacol Sci 31(6):255–265. https://doi.org/10.1016/j.tips.2010.02.005
Martins A, Vieira H, Gaspar H, Santos S (2014) Marketed marine natural products in the pharmaceutical and cosmeceutical industries: tips for success. Mar Drugs 12(2):1066–1101. https://doi.org/10.3390/md12021066
Subramani R, Aalbersberg W (2012) Marine Actinomycetes: an ongoing source of novel bioactive metabolites. Microbiol Res 167(10):571–580. https://doi.org/10.1016/j.micres.2012.06.005
Manivasagan P, Venkatesa J, Kim S.-K. Marine microbiology: bioactive compounds and biotechnological applications. Wiley-VCH Verlag GmbH & Co. KGaA . 2013. pp. 1–19.
Manivasagan P, Venkatesan J, Sivakumar K, Kim S-K (2014) Pharmaceutically active secondary metabolites of marine Actinobacteria. Microbiol Res 169(4):262–278. https://doi.org/10.1016/j.micres.2013.07.014
Sharma R, Ranjan R, Kapardar RK, Grover A (2005) Special section: microbial diversity “unculturable” bacterial diversity: an untapped resource. Curr Sci 89(1)
Kyrpides NC, Hugenholtz P, Eisen JA, Woyke T, Göker M, Parker CT, Amann R, Beck BJ, Chain PSG, Chun J, Colwell RR, Danchin A, Dawyndt P, Dedeurwaerdere T, DeLong EF, Detter JC, De Vos P, Donohue TJ, Dong X-Z, Ehrlich DS, Fraser C, Gibbs R, Gilbert J, Gilna P, Glöckner FO, Jansson JK, Keasling JD, Knight R, Labeda D, Lapidus A, Lee J-S, Li W-J, MA J, Markowitz V, Moore ERB, Morrison M, Meyer F, Nelson KE, Ohkuma M, Ouzounis CA, Pace N, Parkhill J, Qin N, Rossello-Mora R, Sikorski J, Smith D, Sogin M, Stevens R, Stingl U, Suzuki K, Taylor D, Tiedje JM, Tindall B, Wagner M, Weinstock G, Weissenbach J, White O, Wang J, Zhang L, Zhou Y-G, Field D, Whitman WB, Garrity GM, Klenk H-P, Pace N, Achtman M, Wagner M, Pace N, Whitman W, Coleman D, Wiebe W, López-García P, Rodríguez-Valera F, Pedrós-Alió C, Moreira D, Amann R, Ludwig W, Schleifer K, Hugenholtz P, Rappé M, Giovannoni S, Kyrpides N, Hugenholtz P, Kyrpides N, Woese C, Pagani I, Liolios K, Jansson J, Chen I, Smirnova T, Tindall B, Rosselló-Móra R, Busse H, Ludwig W, Kämpfer P, Trüper H, Euzéby J, Tindall B, Kämpfer P, Euzéby J, Oren A, Tindall B, Garrity G, Chain P, Grafham D, Fulton R, Fitzgerald M, Hostetler J, Göker M, Klenk H, Wu D, Hugenholtz P, Mavromatis K, Pukall R, Dalin E, Shih P, Wu D, Latifi A, Axen S, Fewer D, Rinke C, Schwientek P, Sczyrba A, Ivanova N, Anderson I, Nelson K, Weinstock G, Highlander S, Worley K, Creasy H, Field D, Amaral-Zettler L, Cochrane G, Cole J, Dawyndt P, Field D, Garrity G, Gray T, Morrison N, Selengut J, Field D, Sansone S, Collis A, Booth T, Dukes P, Yilmaz P, Kottmann R, Field D, Knight R, Cole J, Walker A, Göker M, Klenk H, Buckley M, Roberts R, Garrity G, Field D, Kyrpides N, Hirschman L, Sansone S, Garrity G, Wilke A, Harrison T, Wilkening J, Field D, Glass E, Yarza P, Richter M, Peplies J, Euzeby J, Amann R, Dedeurwaerdere T, DeVos P, Dijkshoorn L, Connon S, Giovannoni S, Staley J, Konopka A, Faith D, Qin J, Li R, Raes J, Arumugam M, Burgdorf K, Gilbert J, Meyer F, Jansson J, Gordon J, Pace N (2014) Genomic encyclopedia of bacteria and archaea: sequencing a myriad of type strains. PLoS Biol 12(8):e1001920. https://doi.org/10.1371/journal.pbio.1001920
Pye CR, Bertin MJ, Lokey RS, Gerwick WH, Linington RG (2017) Retrospective analysis of natural products provides insights for future discovery trends. Proc Natl Acad Sci U S A 114(22):5601–5606. https://doi.org/10.1073/pnas.1614680114
Gomez-Escribano JP, Alt S, Bibb MJ (2016) Next generation sequencing of Actinobacteria for the discovery of novel natural products. Mar Drugs 14(4):6–8. https://doi.org/10.3390/md14040078.
Jensen PR, Dwight R, Fenical W (1991) Distribution of actinomycetes in near-shore tropical marine sediments, 57 (4).
Mincer TJ, Jensen PR, Kauffman CA, Fenical W (2002) Widespread and persistent populations of a major new marine actinomycete taxon in ocean sediments widespread and persistent populations of a major new marine actinomycete taxon in ocean sediments. Appl Environ Microbiol 68(10):5005–5011. https://doi.org/10.1128/AEM.68.10.5005.
Prieto-Davo A (2013) Phylogenetic and chemical diversity of marine-derived actinomycetes from Southern California sediments. University of California
Prieto-Davo A, Dias T, Gomes SE, Rodrigues S, Parera-Valadez Y, Borralho PM, Pereira F, Rodrigues CMP, Santos-Sanches I, Gaudencio SP (2016) The Madeira Archipelago as a significant source of marine-derived actinomycete diversity with anticancer and antimicrobial potential. Front Microbiol 7(OCT):1–12. https://doi.org/10.3389/fmicb.2016.01594.
Fenical W (1994) Strategies for the discovery of secondary metabolites from marine bacteria: ecological perspectives. Annu Rev Microbiol 48:559–584. https://doi.org/10.1146/annurev.micro.48.1.559
Netzker T, Fischer J, Weber J, Mattern DJ, König CC, Valiante V, Schroeckh V, Brakhage AA (2015) Microbial communication leading to the activation of silent fungal secondary metabolite gene clusters. Front Microbiol 6:299. https://doi.org/10.3389/fmicb.2015.00299
Lam KS (2006) Discovery of novel metabolites from marine actinomycetes. Curr Opin Microbiol 9(3):245–251. https://doi.org/10.1016/j.mib.2006.03.004
Ragini K, Aalbersberg W (2014) Isolation and characterisation of rifamycin W and phenylethylamides from a Fijian marine actinomycete Salinispora arenicola. South Pacific J Nat Appl Sci 32(2):43. https://doi.org/10.1071/SP14007
El Naggar MM, El-Assar SA, Shata AMA (2015) Production of antitumor agents from novel marine actinomycetes isolated from Alexandria, Egypt. Single Cell Biol 4(1). https://doi.org/10.4172/2168-9431.1000e122.
Sanjivkumar M, Ramesh Babu D, Sunganya AM, Silambarasan AM, Balagurunathan R, Immanuel G (2016) Investigation on pharmacological activities of secondary metabolite extracted from a mangrove associated actinobacterium Streptomyces olivaceus (MSU3). Biocatal Agric Biotechnol 6:82–90. https://doi.org/10.1016/J.BCAB.2016.03.001.
Kumar KN, Elavarasi TGA (2013) Studies on antimicrobial activity of marine actinomycetes isolated from Rameswaram , 4 (4), 706–710.
Messaoudi O, Bendahou M, Benamar I, Abdelwouhud D (2015) Identification and preliminary characterization of non-polyene antibiotics secreted by new strain of actinomycete isolated from Sebkha of Kenadsa, Algeria. Asian Pac J Trop Biomed 5(6):438–445. https://doi.org/10.1016/J.APJTB.2015.04.002
Bauer-Gottwein P, Gondwe BRN, Charvet G, Marín LE, Rebolledo-Vieyra M, Merediz-Alonso G (2011) Review: the Yucatán Peninsula karst aquifer, Mexico. Hydrogeol J 19(3):507–524. https://doi.org/10.1007/s10040-010-0699-5
Pérez L, Bugja R, Lorenschat J, Brenner M, Curtis J, Hoelzmann P, Islebe G, Scharf B, Schwalb A (2011) Aquatic ecosystems of the Yucatán Peninsula (Mexico), Belize, and Guatemala. Hydrobiologia 661(1):407–433. https://doi.org/10.1007/s10750-010-0552-9
Lozupone C, Knight R (2005) UniFrac: a new phylogenetic method for comparing microbial communities. Appl Environ Microbiol 71(12):8228–8235. https://doi.org/10.1128/AEM.71.12.8228-8235.2005
Kock I, Maskey RP, Biabani MAF, Helmke E, Laatsch H (2005) 1-Hydroxy-1-norresistomycin and resistoflavin methyl ether: new antibiotics from marine-derived streptomycetes. J Antibiot (Tokyo) 58(8):530–534. https://doi.org/10.1038/ja.2005.73
Mohanraj G, Sekar T (2013) Isolation and screening of actinomycetes from marine sediments for their potential to produce antimicrobials. Int J life Sci Biotechnol pharma Res 2(3):115–126
Fenical W, Jensen PR (2006) Developing a new resource for drug discovery: marine actinomycete bacteria. Nat Chem Biol 2(12):666–673. https://doi.org/10.1038/nchembio841
Tiwari K, Gupta RK (2012) Rare actinomycetes: a potential storehouse for novel antibiotics. Crit Rev Biotechnol 32(2):108–132. https://doi.org/10.3109/07388551.2011.562482
Liu X, Ashforth E, Ren B, Song F, Dai H, Liu M, Wang J, Xie Q, Zhang L (2010) Bioprospecting microbial natural product libraries from the marine environment for drug discovery. J Antibiot (Tokyo) 63(8):415–422. https://doi.org/10.1038/ja.2010.56
Hugenholtz P, Hooper SD, Kyrpides NC (2009) Focus: synergistetes: genomics update. Environ Microbiol 11(6):1327–1329. https://doi.org/10.1111/j.1462-2920.2009.01949.x
Milshteyn A, Schneider JS, Brady SF (2014) Mining the metabiome: identifying novel natural products from microbial communities. Chem Biol 21(9):1211–1223. https://doi.org/10.1016/j.chembiol.2014.08.006
Crüsemann M, O’Neill EC, Larson CB, Melnik AV, Floros DJ, Da Silva RR, Jensen PR, Dorrestein PC, Moore BS (2017) Prioritizing natural product diversity in a collection of 146 bacterial strains based on growth and extraction protocols. J Nat Prod 80(3):588–597. https://doi.org/10.1021/acs.jnatprod.6b00722
Rojas-Herrera R, Zamudio-Maya M, Arena-Ortiz L, Pless R, O’Connor-Sánchez A (2011) Microbial diversity, metagenomics and the Yucatán aquifer. Divers. microbiana, metagenómica y el acuífero Yucatán, 80 (130), 231–240.
Maldonado L, Fragoso-Yáñez D, Pérez-García A, Rosellón-Druker J, Quintana ET (2009) Actinobacterial diversity from marine sediments collected in Mexico. Antonie Van Leeuwenhoek 95(2):111–120. https://doi.org/10.1007/s10482-008-9294-3
Martin GD a, Tan LT, Jensen PR, Dimayuga RE, Fairchild CR, Raventos-Suarez C, Fenical W (2007) Marmycins A and B, cytotoxic pentacyclic C-glycosides from a marine sediment-derived actinomycete related to the genus Streptomyces. J Nat Prod 70(9):1406–1409. https://doi.org/10.1021/np060621r
Maldonado L a, Fenical W, Jensen PR, Kauffman C a, Mincer TJ, Ward AC, Bull AT, Goodfellow M (2005) Salinispora arenicola gen. nov., sp. nov. and Salinispora tropica sp. nov., obligate marine actinomycetes belonging to the family Micromonosporaceae. Int J Syst Evol Microbiol 55:1759–1766. https://doi.org/10.1099/ijs.0.63625-0
Torres-Beltrán M (2012) Evaluation of the Gulf of California as a potential source of bioactive marine Actinobacteria. Ciencias Mar 38(4):609–624. https://doi.org/10.7773/cm.v38i4.2131
Goodfellow M, Kämpfer P, Hans-Jürgen B, Trujillo ME, Suzuki K, Ludwig W, Whitman WB (2012) Bergey’s manual of systematic bacteriology Vol. Five, Second.; London.
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol Biol Evol 30(12):2725–2729. https://doi.org/10.1093/molbev/mst197
Becerril-Espinosa A, Freel KC, Jensen PR, Soria-Mercado IE (2013) Marine Actinobacteria from the Gulf of California: diversity, abundance and secondary metabolite biosynthetic potential. Antonie Van Leeuwenhoek 103(4):809–819. https://doi.org/10.1007/s10482-012-9863-3
Powers EM (1995) Efficacy of the Ryu nonstaining KOH technique for rapidly determining gram reactions of food-borne and waterborne bacteria and yeasts. Appl Environ Microbiol 61(10):3756–3758
Kokare CR, Mahadik KR, Kadam SS, Chopade BA (2004) Isolation of bioactive marine actinomycetes from sediments isolated from Goa and Maharashtra Coastlines (West Coast of India) 33(September):248–256
Duncan K, Haltli B, Gill K a, Kerr RG (2014) Bioprospecting from marine sediments of New Brunswick, Canada: exploring the relationship between total bacterial diversity and Actinobacteria diversity. Mar Drugs 12(2):899–925. https://doi.org/10.3390/md12020899
Schloss PD, Westcott SL, Ryabin T, Hall JR, Hartmann M, Hollister EB, Lesniewski R a, Oakley BB, Parks DH, Robinson CJ, Sahl JW, Stres B, Thallinger GG, Van Horn DJ, Weber CF (2009) Introducing Mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl Environ Microbiol 75(23):7537–7541. https://doi.org/10.1128/AEM.01541-09
Chao A, Chiu C-HSR (2016) Estimation and comparison. Wiley StatsRef: statistics reference online; John Wiley & Sons, Ltd: Chichester, UK, pp 1–26
Özcan K, Aksoy SÇ, Kalkan O, Uzel A, Hames-Kocabas EE, Bedir E (2013) Diversity and antibiotic-producing potential of cultivable marine-derived actinomycetes from coastal sediments of Turkey. J Soils Sediments 13(Wilkins 1996):1493–1501. https://doi.org/10.1007/s11368-013-0734-y
Jensen PR, Dwight R, Fenical W (1991) Distribution of actinomycetes in near-shore tropical marine sediments. Appl Environ Microbiol 57(4):1102–1108
Youssef NH, Couger MB, McCully AL, Criado AEG, Elshahed MS (2015) Assessing the global phylum level diversity within the bacterial domain: a review. J Adv Res 6:269–282
Floros DJ, Jensen PR, Dorrestein PC, Koyama N (2016) A metabolomics guided exploration of marine natural product chemical space. Metabolomics 12
Prieto-Davó, a; Fenical, W.; Jensen, P. Comparative actinomycete diversity in marine sediments. Aquat Microb Ecol 2008, 52 (July), 1–11 DOI: https://doi.org/10.3354/ame01211.
Jensen PR, Williams PG, Oh D-C, Zeigler L, Fenical W (2007) Species-specific secondary metabolite production in marine actinomycetes of the genus Salinispora. Appl Environ Microbiol 73(4):1146–1152. https://doi.org/10.1128/AEM.01891-06
Aranda-Cirerol N (2002) Water quality and sustainable development in Yucatán Peninsula. Environ. Res. J, 5 (6).
Herrera-Silveira JA, Morales-ojeda SM (2009) Evaluation of the health status of a coastal ecosystem in Southeast Mexico: assessment of water quality, phytoplankton and submerged aquatic vegetation. Mar Pollut Bull 59(1–3):72–86. https://doi.org/10.1016/j.marpolbul.2008.11.017
Valdes DS, Real E (1998) Variations and relationships of salinity, nutrients and suspended solids in Chelem coastal lagoon at Yucatan, Mexico. Indian J Mar Sci 27(2):149–156
Rashad FM, Fathy HM, El-zayat AS, Elghonaimy AM (2015) Isolation and characterization of multifunctional Streptomyces species with antimicrobial, nematicidal and phytohormone activities from marine environments in Egypt. 175:34–47.
Tian X, Zhang Z, Yang T, Chen M, Li J, Chen F, Yang J, Li W, Zhang B, Zhang Z, Wu J, Zhang C, Long L, Xiao J (2016) Comparative genomics analysis of Streptomyces species reveals their adaptation to the marine environment and their diversity at the genomic level. Front Microbiol 7:998. https://doi.org/10.3389/fmicb.2016.00998
Fan L, Liu Y, Li Z, Baumann HI, Kleinschmidt K, Ye W, Imhoff JF, Kleine M, Cai D (2011) Draft genome sequence of the marine Streptomyces sp. strain PP-C42, isolated from the Baltic Sea. J Bacteriol 193(14):3691–3692. https://doi.org/10.1128/JB.05097-11
Leipoldt F, Zeyhle P, Kulik A, Kalinowski J, Heide L, Kaysser L (2015) Diversity of ABBA prenyltransferases in marine Streptomyces sp. CNQ-509: promiscuous enzymes for the biosynthesis of mixed terpenoid compounds. PLoS One 10(12):e0143237. https://doi.org/10.1371/journal.pone.0143237
Feling RH, Buchanan GO, Mincer TJ, Kauffman CA, Jensen PR, Fenical W (2003) Salinosporamide A: a highly cytotoxic proteasome inhibitor from a novel microbial source, a marine bacterium of the new genus Salinospora. Angew Chem Int Ed 42(3):355–357. https://doi.org/10.1002/anie.200390115
Tian XP, Long LJ, Wang FZ, Xu Y, Li J, Zhang J, Zhang CS, Zhang S, Li WJ (2012) Streptomyces nanhaiensis sp. Nov., a marine streptomycete isolated from a deep-sea sediment. Int J Syst Evol Microbiol 62(4):864–868. https://doi.org/10.1099/ijs.0.031591-0
Murray G (2007) Constructing paradise: the impacts of big tourism in the Mexican coastal zone. Coast Manag 35(2–3):339–355. https://doi.org/10.1080/08920750601169600
Fenchel T, Finlay BJ (2004) The ubiquity of small species: patterns of local and global diversity. Bioscience 54(8):777. https://doi.org/10.1641/0006-3568(2004)054[0777:TUOSSP]2.0.CO;2
R Core Team. R: a language and environment for statistical computing; 2015; Vol. 1.
Rahalison L, Hamburger M, Hostettmann K, Monod M, Frenk E (1991) A bioautographic agar overlay method for the detection of antifungal compounds from higher plants. Phytochem Anal 2(5):199–203. https://doi.org/10.1002/pca.2800020503
Acknowledgements
Authors would like to acknowledge Dr. Emanuel Hernández-Núñez at the Department for Marine Resourcesin CINVESTAV, Mérida for his help with the GC-MS analysis of the crude extracts.
Funding
We would like to recognize the Mexican Council of Science and Technology (CONACyT) for their support through fellowships for Parera-Valadez (Masters in Science fellowship: 560614) and Yam-Puc (Postdoctoral fellowship). We would like to recognize the National Autonomous University of Mexico (UNAM) for their funding through PAPIIT TA200212 and TA200415 and UNAM School of Chemistry for their funding through PAIP.
Author information
Authors and Affiliations
Contributions
Parera-Valadez contributed by heading the research and writing the paper. Yam-Puc and Figueroa-Saldivar contributed with chemical analyses, bioautography assays, and manuscript writing. López-Aguiar and Márquez-Velázquez contributed with field collections, laboratory procedures, and manuscript revisions. Borges-Argáez and Cáceres-Farfán contributed with bioautography assays.
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Electronic Supplementary Material
Figure S1
Phylogenetic tree with all actinomycete isolates from the coast of the Yucatan peninsula. Alignment and phylogeny were performed following same methods as those reported in Fig. 1 (PDF 37 kb)
Figure S2
Rarefaction curves for the 12 locations sampled. No location was thoroughly sampled and more processing or sampling are needed to reach saturation. (PDF 299 kb)
Figure S3
Detection of the antimicrobial activity of the ethyl acetate extract (5%) from actinomycete NCA004 against Staphylococcus aureus by the bioautography assay. The highly active yellow compound corresponds to resistomycin. 1) Ethyl acetate extract of strain NCA002. 2) Ethyl acetate extract of strain NCA004. 3) Dichloromethane extract of strain NCA004. 4) Acetone extract of strain NCA004. Positive control: amikacin (0.1 mg/mL), negative control: 5 μl of dichloromethane/methanol 1:1. (PDF 728 kb)
Figure S4
Chromatogram displaying resistomycin peak at 12.057 min. Figure S4: UPLC-MS analysis of Ethyl Acetate extract for strain NCA004 showing UV peaks and mass consistent with resistomycin. (PDF 5364 kb)
Figure S5 UV profile and MS data that confirm presence of resitomycin in extract Figure S5 was mentioned in the text but the corresponding data is missing. Please provide.Figure S5 has been uploaded
Rights and permissions
About this article

Cite this article
Parera-Valadez, Y., Yam-Puc, A., López-Aguiar, L.K. et al. Ecological Strategies Behind the Selection of Cultivable Actinomycete Strains from the Yucatan Peninsula for the Discovery of Secondary Metabolites with Antibiotic Activity. Microb Ecol 77, 839–851 (2019). https://doi.org/10.1007/s00248-019-01329-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00248-019-01329-3