
Abstract
Variability in habitat positioning within seascapes (over kilometres) influences fauna assemblage composition, but the characteristics of a habitat patch (10–100s of metres) influence how species use that patch and how variable assemblages are within habitats. Understanding the relative influence of these two contrasting scales is crucial to improving the management of marine habitats. We used baited remote underwater video systems (BRUVS) to quantify seagrass fish assemblages, and took seagrass cores to quantify seagrass metrics, at ten sites across three seasons in Moreton Bay, Australia to determine if fish are influenced more by seascape context or metrics of seagrass habitat complexity. We found that fish species richness and assemblage composition are most influenced by large-scale variability in seascape (e.g. proximity to ocean and mangroves). However, variability in habitat complexity (e.g. seagrass blade length and density) and proximity to mangrove forests had the greatest effect on assemblage beta diversity. Connectivity with other habitats plays a vital role in structuring the fish community, as it is crucial for daily feeding excursions (mangrove forests) and spawning/reproduction (proximity to ocean). Continuous, non-patchy seagrass beds are however vital to how individuals use a seagrass meadow, with beta diversity being higher in seagrass meadows that were less patchy. Identifying how habitat attributes and context influence fish assemblages is vital for optimizing conservation initiatives. Therefore, we suggest that monitoring populations with biodiversity metrics such as beta diversity can be effective in determining areas that are critical for conservation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Animal movement is influenced by dispersal capabilities, connectivity with adjacent habitats, abiotic variables and anthropogenic influences (Cloern 2007; Olds et al. 2016). Variations in land and seascapes (e.g. proximity to other habitats and the size of a habitat) influence assemblage composition and biodiversity, structuring the way animals use a landscape and the processes that underpin ecosystem resilience (Massol et al. 2011; Turner 1989). Many studies have focused on how marine ecosystems are influenced by the spatial context of a seascape, with many of these especially focusing on fish and invertebrates in seagrass meadows (Boström et al. 2011; Irlandi and Crawford 1997; Robbins and Bell 1994); however, many of these are yet to assess the different measures of fish assemblages and how these are influenced by variable seascape factors.
Variability in seascapes has clear effects on assemblage composition; however, variability in the complexity of habitat patches changes the composition of organisms inhabiting the patch (Anderson et al. 2011; Bell et al. 1988; Díaz et al. 2015; Jungerstam et al. 2014). Small-scale changes in habitat complexity within a habitat patch (e.g. habitat density, complexity or area) can interact with the way species use a land or seascape, thereby further altering species composition and ecological functioning (Díaz et al. 2015; Graham and Nash 2012; McElhinny et al. 2005; Munday et al. 2008). Investigating how habitat complexity influences assemblage composition, species diversity and variability within a patch [e.g. beta diversity, (Anderson et al. 2006, 2011)] is therefore important in many habitats, particularly in the context of anthropogenic effects which can cause large and small-scale changes in land or seascape context (Lindenmayer et al. 2000; Ward and Tockner 2001).
Seagrass ecosystems remain one of the most threatened ecosystems in the world, with rates of decline matching that of other threatened ecosystems such as mangroves, coral reefs# and tropical rainforests (Waycott et al. 2009). Seagrasses are highly productive, provide numerous ecosystem services to coastal regions# and act as a nursery for many harvested fish and invertebrate species (Boström et al. 2006; Nagelkerken et al. 2001; Unsworth et al. 2015). Seagrasses are often located within heterogeneous seascapes, and a range of seascape factors are known to influence the composition of the fish assemblage (Bell et al. 1988; Boström et al. 2011; Connolly and Hindell 2006). Similarly, as seagrasses are naturally influenced by light availability, depth# and sedimentation, seagrass beds can have a naturally variable structure within individual seagrass meadows (Maxwell et al. 2014). Despite the ecological and economic importance of seagrass ecosystems, the influence of small- and large-scale variability in the structure of seagrass ecosystems on fish assemblages remains poorly understood.
In this study, we compare the relative influence of seagrass meadow positioning within seascapes to small-scale changes in seagrass complexity on seagrass fish assemblages. We used measures of fish species richness; beta dispersion (a distance-based multivariate measure of beta diversity or variation within a community (Anderson et al. 2006), and henceforth referred to as beta diversity) and composition of the seagrass fish assemblage to determine the main factors influencing fish in seagrass meadows in Moreton Bay, Eastern Australia. These measures provide an understanding of diversity between (e.g. species diversity) and within [e.g. beta diversity, (Anderson et al. 2006)] seagrass meadows, which is crucial for the continued management of fish communities within seagrass meadows. Beta diversity has become a central theme for determining priority areas for conservation as it provides the mechanism for determining how individual habitat patches deal with a range of factors (Anderson et al. 2011; Barboza et al. 2015). Moreton Bay offers an ideal location to test these aims as it presents a heterogeneous seascape of multiple subtropical habitats (especially marginal coral reefs and mangroves) (Gilby et al. 2016) and seagrass meadows that are highly variable in structural complexity (i.e. shoot length, density# and seagrass biomass) due to a gradient from estuarine-dominated water to oceanic (Maxwell et al. 2014). We therefore expect that large-scale variations in seascape context will drive the changes in fish assemblage composition and species richness, as seen elsewhere in Moreton Bay [e.g. coral reefs, see (Olds et al. 2012)], while variability in the structural complexity within seagrass meadows will influence beta diversity more (Gullström et al. 2008). It is becoming more important for managers to understand the factors that influence the distribution of faunal communities within and between habitat patches and here, we provide a simple method to determine these drivers of change in seagrass ecosystems.
Methods
Study site
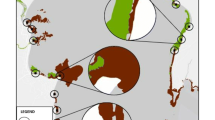
We surveyed fish assemblages and seagrass meadow variables at ten seagrass meadows in Moreton Bay, Queensland, a subtropical embayment in eastern Australia (Fig. 1). Moreton Bay is bordered to the east by three large sand islands, with oceanic water exchanging through three passages, and to the west by mainland Australia, where multiple estuaries discharge into the Bay (Gibbes et al. 2014). Sites within Moreton Bay were located along a gradient of distances from the ocean, with larger seagrass beds located closer to oceanic input and nearby coral reefs and seagrass beds further from the ocean were nearer to mangroves. A water quality gradient was also present between near-ocean sites in clear water, high salinity# and low nutrients, while those further away from oceanic input have higher nutrient levels, reduced salinity and higher turbidity (Maxwell et al. 2014). All meadows studied were comprised of eelgrass Zostera muelleri. The ambient water quality gradient influences seagrass growth, in that those seagrass meadows closer to the ocean have smaller shoots, higher shoot density and a larger proportion of their biomass located underneath the sediment (Maxwell et al. 2014). Each seagrass meadow was sampled three times, in the austral winter (August 2014), spring (November 2014) and summer (February 2015). The intention of this was to include the full range of temporal variability that is experienced in this subtropical system, but as season was not a replicated factor, no conclusions will be made regarding seasonality.

Seagrass habitats in Moreton Bay, Australia (c. 27°18’S; 153°17’E), where BRUVS were used for sampling fish community composition and seagrass metrics were surveyed
Fish assemblage surveys
Fish assemblages were surveyed using eight baited remote underwater video stations (BRUVS) at each of the ten sites during each of the three sampling periods cross one year (Harvey et al. 2007; Malcolm et al. 2007). All BRUVS were deployed in ~1.5 m of water at high tide and were all spaced at least 200 m apart to avoid sampling the same individuals on multiple cameras. A spacing of at least 200 m between each BRUVS deployment was used so that no individual fish was counted on multiple BRUVS there making BRUVS deployments independent of one another, meaning there is no need to have each individual BRUVS nested inside each seagrass meadow (Malcolm et al. 2007). BRUVS consisted of a GoPro video camera recording in high definition fixed to a 5 kg weight with a bait bag (500 g of chopped pilchards) positioned 0.5 m from the camera (Bernard and Götz 2012). Each BRUVS deployment lasted for one hour, giving a total of 240 h of sampling time from the entire study (Harvey et al. 2007; Santana-Garcon et al. 2014). Fish abundance, species richness and fish community composition were recorded from video footage using the MaxN statistic (Willis and Babcock 2000).
Seascape and seagrass variables
Seascape variables included in the study were seagrass meadow area (km2) and proximity to the nearest coral reef (km), mangrove forest (km) and the open ocean (km). These variables were all calculated in ArcGIS (ESRI, Redlands, CA, USA). Distance to the open ocean was used as many species make spawning migrations that include movements outside of Moreton Bay, but it also incorporates a salinity, nutrient and turbidity gradient (Henderson et al. 2017). Seagrass metrics were calculated from ten replicate cores (15 cm diameter, 10 cm deep) taken at each seagrass meadow location during each season, taken directly after fish assemblage surveys. Shoot density, length and above/below ground biomass were calculated for each site.
Statistical analysis
A range of diversity metrics was calculated from the fish assemblage data, including species richness, beta diversity and community composition. Species richness was represented as the total number of species recorded at each site using BRUVS. Beta diversity was calculated for each site during each season using a distance-based test for homogeneity of multivariate dispersions (Anderson et al. 2006, 2011; Anderson and Santana-Garcon 2015). Here, beta diversity is the average distance from a group centroid (calculated using a Modified Gower Log2 resemblance matrix) of the composition of the assemblages recorded from each BRUVS replicate at each site (Anderson et al. 2011). Therefore, a greater average distance from the centroid corresponds to greater variability in species composition, and vice versa. Consequently, we obtained only one value for beta diversity for each seagrass meadow during each season, resulting in thirty observations for analysis. To keep this consistent between analyses, we calculated only one value per site, per season, for species richness also; again from each of the eight BRUVS replicates at each site.
Generalized additive models (GAMs) were used to determine which seascape and seagrass metrics most influenced the species richness and beta diversity measured. All GAMs were fitted in the R statistical environment using the packages mgcv and MuMIn (Bartoń 2013). Season was tested as part of the GAM analysis for both species richness and beta diversity to ensure it had no impact on fish assemblages. However, for the GAM on beta diversity, the standard errors for seagrass blade length and shoot density samples taken in each meadow were used instead of the average measurements as beta diversity is a measure of variability within a site. GAMs were preferred, as we could not assume linearity for all the factors used in the analysis (Gilby et al. 2015). Thirty observations were used in each model and due to this, models were restricted to four or less factors and the models were compared using the Akaike information criterion corrected (AICc), with best-fit models having the lowest AICc. Model selection for GAMs was done using the dredge function in the MuMIn package in the R statistical environment, which selects the best model from all possible combinations of factors. GAMs were fitted using a Gaussian distribution with all temporal factors being represented by the average seasonal value for each site (i.e. the average values for shoot density, length and biomass during each of the three seasons). All models were checked for homogeneity and normality to ensure model performance. GAMs return a best-fit model for each of the measures and also a relative importance value. Relative factor importance was calculated by the summed weighted AICc value for each model containing that factor, with the highest values being those that contribute the most to the models (Burnham and Anderson 2002).
In order to determine which of seascape context and seagrass metrics most influenced the entire fish community composition between sites, a BIOENV was used (Clarke and Ainsworth 1993). Matrices of fish community composition (Bray Curtis similarity) and the environmental factors (Euclidean distance) were compared to determine which factors correlated the most with the fish community, and a principle coordinates ordination (PCO) was used to visualize significant factors (Anderson and Willis 2003). The same PCO was then used to visualize which species correlated most with changes in fish communities. The BIOENV analysis was restricted to include no more than four factors in the best model and returns a correlation value. All factors were compared prior to analysis to ensure no autocorrelation, with no factors being removed from the analysis.
Results
We recorded 61 fish species within seagrass meadows in Moreton Bay. The lowest number of species was recorded at the site furthest from the ocean during summer, with only two species being present across all BRUV replicates (See Table S1). The highest number of species recorded on BRUVS within a single seagrass meadow was 25, which was recorded during summer at site 3. Fish species richness was found to be highest at sites with large seagrass beds (Fig. 2a; Table S1) and was generally higher at sites further away from mangroves (Fig. 2c, e). The two best-fit models for species richness were both found to contain total seagrass area and distance to mangroves, while one also contained shoot density (Table 1). GAM plots for species richness show that total seagrass area and distance to mangroves have a positive relationship with species richness (Fig. 3).

Species richness in seagrass meadows of Moreton Bay, Australia, is highest in seagrass meadows that are larger (a) and further away from mangrove forests (c). Beta diversity was found to be higher in individual seagrass meadows that are more uniform throughout, with lower variation in shoot length (b) and shoot density (d) being the most influential factors along with distance from mangrove forests. In plots e (Species richness) and f (Beta diversity), values nearer to one indicate a higher correlation between that factor and the dependent variable of interest. ± indicates the direction of the relationship for each factor. These numbers represent the relative importance values that are calculated from summed weighted AICc values from each model that each individual variable is present in

Correlation plots from GAM outputs for species richness and beta diversity. ‘s’ on the y-axis indicate the application of a smoother in the GAM function, and numbers indicate the estimated degrees of freedom of the smooth function. Shaded grey regions indicate 95% confidence intervals
Beta diversity was influenced most by seagrass metrics as opposed to the seascape context that the seagrass meadow was present in. Distance to mangroves and variation in shoot length (Fig. 2b) and to a lesser extent shoot density (Fig. 2d) had the largest influence on the measures of beta diversity (Fig. 2f). Similar to the best-fit models for species richness, the best-fit model for beta diversity included distance to mangroves and shoot length standard error (Table 1). All remaining best-fit models for beta diversity included seagrass metrics; with shoot density, length and seagrass biomass all being included (Table 1). GAM plots for beta diversity show that distance to mangroves and shoot length standard error have a negative relationship with beta diversity (Fig. 3). Season was not present in any of the top models for either species richness or beta diversity.
Fish community composition was most influenced by seascape factors; distance to coral reefs (r = 0.515) and distance to mangroves (r = 0.555) correlating most (Fig. 4; Table 2). Fish communities across the bay were dominated numerically by omnivorous fish species Pelates sexlineatus and Acanthopagrus australis, herbivorous species Siganus fuscescens and multiple carnivorous Lethrinus species. Changes in the abundances of these species between seagrass meadow locations are the main driver in differences in seagrass fish community composition. A. australis abundances correlated highest with changes in total seagrass area (Fig. 4). S. fuscescens and Lethrinus species correlated the highest with changes in distance to mangroves (Fig. 4). Similarly, P. sexlineatus abundance changes correlated most with changes in mangrove distance, reef distance and shoot length (Fig. 4).

Principle coordinates ordination (PCO) displaying fish community composition recorded at each seagrass meadow, the effects of seascape context and seagrass metrics and which species were most impacted by changes in community composition. The length of a vector overlay indicates the strength of the correlation. Replicate points represent the three different seasons each site was sampled during the study
Discussion
Large-scale variability in habitat context drives species assemblages in many different terrestrial and marine systems around the world (Olds et al. 2012; Rudnick et al. 2012). We show that variability in seascape context drives differences in fish species richness and assemblage composition between seagrass meadows, but variations in beta diversity are driven more by small-scale changes in the structural complexity (e.g. seagrass blade length and density) of seagrass meadows and proximity to mangroves. Proximity to a mangrove forest was found to decrease species diversity and alter community composition, namely the abundances of species such as S. fuscescens and multiple Lethrinus species (Olds et al. 2012), while the size of seagrass meadows was similarly important for species diversity and the abundance of A. australis, but not overall community composition (Boström et al. 2006). This is important for the management of seagrass ecosystems as this allows managers to determine what factors influence species turnover and variability (beta diversity) and species diversity (species richness), therefore furthering the understanding of the ideal management for this ecosystem.
The location of a seagrass meadow within the broader seascape was a major factor in determining the composition of fish assemblages and diversity. Seagrass meadows that were large and further away from mangroves supported a more diverse fish assemblage, with those seagrass meadows also being within close proximity to coral reefs and the open ocean (Connolly and Hindell 2006; Nagelkerken et al. 2000). Numerous fish species associated to seagrass or coral reefs make daily excursions to other habitats, resulting in their abundance and species richness being higher in seagrass meadows with close proximity to coral reefs (Olds et al. 2012). This likely peaks at high tide when all habitats, coral reefs, mangrove forests and seagrass meadows, are fully inundated, possibly resulting in some individuals being missed by the BRUVS sampling as they were in other habitats (Olds et al. 2012). Similarly, proximity to the open ocean can have a significant effect on the composition of fish species, with the seagrass beds receiving a greater supply of larvae (Ford et al. 2010), but also support species that make ontogenetic movements from inshore areas to deeper offshore reefs (Henderson et al. 2017; Kimirei et al. 2011; Sumpton et al. 2008). Here, our results show that changes in seagrass fish assemblage composition are likely a result of seascape context and connectivity with other habitats.
We used beta diversity to determine how similar assemblage composition is within different seagrass patches and how they are compared to others (Anderson et al. 2006, 2011; Anderson and Santana-Garcon 2015), looking further into the factors between these patches that cause different seagrass beds to be heterogeneous or homogeneous. Variability in seagrass complexity (e.g. shoot density, length and seagrass biomass) was the most important factor in determining how beta diversity varied between seagrass beds in this study (Bennett and Gilbert 2016). The patchiness of seagrass meadows across Moreton Bay had a clear influence on the heterogeneity of fish species within a meadow (Barboza et al. 2015). Seagrass meadows with more variable blade height measurements contained lower levels of beta diversity among fish assemblages throughout the entire meadow. Due to the poor water quality that is experienced in Moreton Bay, particularly within the southern portions of the bay, seagrass beds are often fragmented and heterogeneous in their structure (Maxwell et al. 2014). These heterogeneous, patchy seagrass beds appear to be decreasing the variability within fish communities at the seagrass meadow level. While this result seems counterintuitive to how complexity drives species abundances in many habitats, here we expect that heterogeneity in seagrass meadows is linked to the patchiness experienced in these meadows (Gullström et al. 2008). However, these effects of habitat heterogeneity are unable to be separated from that of the poor environmental conditions experienced in these seagrass meadows. Changes in habitat complexity can be highly important for heterogeneous seagrass fish communities. With beta diversity increasingly being used as a monitoring tool for assessing variability in faunal communities (Anderson et al. 2011; Barboza et al. 2015), it is important to monitor both fish communities and habitat structure so that we can improve the understanding of the factors that drive variability within marine systems (Barboza et al. 2015; Jiang et al. 2015).
Large, well-connected seagrass beds provide habitat for a more diverse seagrass fish community. However, within this context, seagrass meadows with smaller and consistently dense seagrass blades are responsible for increased variability in fish community composition of individual seagrass meadows. These large seagrass meadows are likely to be less patchy and therefore provide more consistent habitat for seagrass fish assemblages. Our results show a strong effect of connectivity, with richness being higher closest to the ocean, where seagrass meadows have a high proximity to inshore coral reefs and are generally larger. We highlight the influences that drive variability within seagrass meadows and also the factors that influence changes at a larger scale. We suggest that empirical data on what factors influence these different scales of variation are critical to improved conservation initiatives in seagrass ecosystems and elsewhere.
References
Anderson MJ, Santana-Garcon J (2015) Measures of precision for dissimilarity-based multivariate analysis of ecological communities. Ecol Lett 18:66–73
Anderson MJ, Willis TJ (2003) Canonical analysis of principal coordinates: a useful method of constrained ordination for ecology. Ecology 84:511–525
Anderson MJ, Ellingsen KE, McArdle BH (2006) Multivariate dispersion as a measure of beta diversity. Ecol Lett 9:683–693
Anderson MJ, Crist TO, Chase JM, Vellend M, Inouye BD, Freestone AL, Sanders NJ, Cornell HV, Comita LS, Davies KF (2011) Navigating the multiple meanings of β diversity: a roadmap for the practicing ecologist. Ecol Lett 14:19–28
Bartoń K (2013) MuMIn: Multi-model inference. In: R package version 1.9. 13. The comprehensive R archive network (CRAN), Vienna, Austria
Barboza LGA, Mormul RP, Higuti J (2015) Beta diversity as a tool for determining priority streams for management actions. Water Sci Technol 71:1429–1435
Bell JD, Steffe AS, Westoby M (1988) Location of seagrass beds in estuaries: effects on associated fish and decapods. J Exp Mar Bio Ecol 122:127–146
Bennett JR, Gilbert B (2016) Contrasting beta diversity among regions: how do classical and multivariate approaches compare? Global Ecol Biogeogr 25:368–377
Bernard A, Götz A (2012) Bait increases the precision in count data from remote underwater video for most subtidal reef fish in the warm-temperate Agulhas bioregion. Mar Ecol Prog Ser 471:235–252
Boström C, Jackson EL, Simenstad CA (2006) Seagrass landscapes and their effects on associated fauna: a review. Estuar Coast Shelf S 68:383–403
Boström C, Pittman SJ, Simenstad C, Kneib RT (2011) Seascape ecology of coastal biogenic habitats: advances, gaps, and challenges. Mar Ecol Prog Ser 427:191–217
Burnham K, Anderson D (2002) Information and likelihood theory: a basis for model selection and inference. Model Sel Multimodel Inference Pract Inf Theor Approach 2:49–97
Clarke K, Ainsworth M (1993) A method of linking multivariate community. Mar Ecol Prog Ser 92:205–219
Cloern JE (2007) Habitat connectivity and ecosystem productivity: implications from a simple model. Am Nat 169:21–33
Connolly RM, Hindell JS (2006) Review of nekton patterns and ecological processes in seagrass landscapes. Estuar Coast Shelf S 68:433–444
Díaz ER, Erlandsson J, Westerbom M, Kraufvelin P (2015) Depth-related spatial patterns of sublittoral blue mussel beds and their associated macrofaunal diversity revealed by geostatistical analyses. Mar Ecol Prog Ser 540:121–134
Ford JR, Williams RJ, Fowler AM, Cox DR, Suthers IM (2010) Identifying critical estuarine seagrass habitat for settlement of coastally spawned fish. Mar Ecol Prog Ser 408:181–193
Gibbes B, Grinham A, Neil D, Olds A, Maxwell P, Connolly R, Weber T, Udy N, Udy J (2014) Moreton Bay and its estuaries: a sub-tropical system under pressure from rapid population growth. Estuaries of Australia in 2050 and beyond. Springer, Netherlands, pp 203–222
Gilby BL, Maxwell PS, Tibbetts IR, Stevens T (2015) Bottom-up factors for algal productivity outweigh no-fishing marine protected area effects in a marginal coral reef system. Ecosystems 18:1–14
Gilby BL, Tibbetts IR, Olds AD, Maxwell PS, Stevens T (2016) Seascape context and predators override water quality effects on inshore coral reef fish communities. Coral Reefs 35:1–12
Graham NAJ, Nash KL (2012) The importance of structural complexity in coral reef ecosystems. Coral Reefs 32:1–12
Gullström M, Bodin M, Nilsson PG, Ohman MC (2008) Seagrass structural complexity and landscape configuration as determinants of tropical fish assemblage composition. Mar Ecol Prog Ser 363:241–255
Harvey ES, Cappo M, Butler JJ, Hall N, Kendrick GA (2007) Bait attraction affects the performance of remote underwater video stations in assessment of demersal fish community structure. Mar Ecol Prog Ser 350:245–254
Henderson CJ, Olds AD, Lee SY, Gilby BL, Maxwell PS, Connolly RM, Stevens T (2017) Marine reserves and seascape context shape fish assemblages in seagrass ecosystems. Mar Ecol Prog Ser 566:135–144
Irlandi EA, Crawford MK (1997) Habitat linkages: the effect of intertidal saltmarshes and adjacent subtidal habitats on abundance, movement, and growth of an estuarine fish. Oecologia 110:222–230
Jiang Y, Xu G, Xu H (2015) Use of multivariate dispersion to assess water quality based on species composition data. Environ Sci Pollut R 23:3267–3272
Jungerstam J, Erlandsson J, McQuaid CD, Porri F, Westerbom M, Kraufvelin P (2014) Is habitat amount important for biodiversity in rocky shore systems? A study of South African mussel assemblages. Mar Biol 161:1507–1519
Kimirei IA, Nagelkerken I, Griffioen B, Wagner C, Mgaya YD (2011) Ontogenetic habitat use by mangrove/seagrass-associated coral reef fishes shows flexibility in time and space. Estuar Coast Shelf S 92:47–58
Lindenmayer DB, Margules CR, Botkin DB (2000) Indicators of biodiversity for ecologically sustainable forest management. Conserv Biol 14:941–950
Malcolm H, Gladstone W, Lindfield S, Wraith J, Lynch T (2007) Spatial and temporal variation in reef fish assemblages of marine parks in New South Wales, Australia - baited video observations. Mar Ecol Prog Ser 350:277–290
Massol F, Gravel D, Mouquet N, Cadotte MW, Fukami T, Leibold MA (2011) Linking community and ecosystem dynamics through spatial ecology. Ecol Lett 14:313–323
Maxwell PS, Pitt KA, Burfeind DD, Olds AD, Babcock RC, Connolly RM (2014) Phenotypic plasticity promotes persistence following severe events: physiological and morphological responses of seagrass to flooding. J Ecol 102:54–64
McElhinny C, Gibbons P, Brack C, Bauhus J (2005) Forest and woodland stand structural complexity: its definition and measurement. Forest Ecol Manag 218:1–24
Munday PL, Jones GP, Pratchett MS, Williams AJ (2008) Climate change and the future for coral reef fishes. Fish Fish 9:261–285
Nagelkerken I, van der Velde G, Gorissen MW, Meijer GJ, Van’t Hof T, den Hartog C (2000) Importance of mangroves, seagrass beds and the shallow coral reef as a nursery for important coral reef fishes, using a visual census technique. Estuar Coast Shelf S 51:31–44
Nagelkerken I, Kleijnen S, Klop T, van den Brand RACJ, Cocheret de la Morinière E, van der Velde G (2001) Dependence of Caribbean reef fishes on mangroves and seagrass beds as nursery habitats: a comparison of fish faunas between bays with and without mangroves/seagrass beds. Mar Ecol Prog Ser 214:225–235
Olds AD, Connolly RM, Pitt KA, Maxwell PS (2012) Primacy of seascape connectivity effects in structuring coral reef fish assemblages. Mar Ecol Prog Ser 462:191–203
Olds AD, Connolly RM, Pitt KA, Pittman SJ, Maxwell PS, Huijbers CM, Moore BR, Albert S, Rissik D, Babcock RC, Schlacher TA (2016) Quantifying the conservation value of seascape connectivity: a global synthesis. Global Ecol Biogeogr 25:3–15
Robbins BD, Bell SS (1994) Seagrass landscapes: a terrestrial approach to the marine subtidal environment. Trends Ecol Evol 9:301–304
Rudnick D, Ryan S, Beier P, Cushman S, Dieffenbach F, Epps C, Gerber L, Hartter J, Jenness J, Kintsch J (2012) The role of landscape connectivity in planning and implementing conservation and restoration priorities. In: Issues in ecology. Ecological Society of America
Santana-Garcon J, Newman SJ, Harvey ES (2014) Development and validation of a mid-water baited stereo-video technique for investigating pelagic fish assemblages. J Exp Mar Bio Ecol 452:82–90
Sumpton W, Mayer D, Brown I, Sawynok B, McLennan M, Butcher A, Kirkwood J (2008) Investigation of movement and factors influencing post-release survival of line-caught coral reef fish using recreational tag-recapture data. Fish Res 92:189–195
Turner MG (1989) Landscape ecology: the effect of pattern on process. Annu Rev Ecol Syst 20:171–197
Unsworth RKF, Collier CJ, Waycott M, McKenzie LJ, Cullen-Unsworth LC (2015) A framework for the resilience of seagrass ecosystems. Mar Pollut Bull 100:34–46
Ward Tockner (2001) Biodiversity: towards a unifying theme for river ecology. Freshw Biol 46:807–819
Waycott M, Duarte CM, Carruthers TJ, Orth RJ, Dennison WC, Olyarnik S, Calladine A, Fourqurean JW, Heck KL, Hughes AR (2009) Accelerating loss of seagrasses across the globe threatens coastal ecosystems. P Natl Acad Sci 106:12377–12381
Willis TJ, Babcock RC (2000) A baited underwater video system for the determination of relative density of carnivorous reef fish. Mar Freshwater Res 51:755–763
Acknowledgements
We would like to thank the staff of the Moreton Bay Research Station for their support with fieldwork and the Australian Rivers Institute and School of Environment at Griffith University for funding this project. We would like to acknowledge the comments from the anonymous reviewers and editor, which have thoroughly improved this manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that there is no conflict of interest. All funding for the project were supplied from the Griffith University School of Environment as part of a postgraduate scholarship.
Ethical approval
All applicable institutional guidelines for the care and use of animals were followed. Ethical approval was under the Griffith University Animal Ethics Committee ENV/07/13 AEC.
Additional information
Responsible Editor: P. Kraufvelin.
Reviewed by undisclosed experts.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Henderson, C.J., Gilby, B.L., Lee, S.Y. et al. Contrasting effects of habitat complexity and connectivity on biodiversity in seagrass meadows. Mar Biol 164, 117 (2017). https://doi.org/10.1007/s00227-017-3149-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-017-3149-2