
Abstract
In the Caribbean, gorgonian corals dominate many coral reefs, while scleractinian coral cover has declined. Gorgonian corals deal with stressors such as lesions caused by wave action, predation, or human activities. In June 2012, artificial wounds were inflicted on branches of the gorgonians Eunicea flexuosa and Pseudoplexaura porosa found at 3–5 m depth on a patch reef (20°52′5.23″N, 86°51′58.92″W) near Puerto Morelos, Mexico. Following healing, injured and uninjured branches were collected to determine the effect of lesions on gorgonian biochemistry, symbiosis, microbiome, and immune response. Lesion recovery in E. flexuosa took twice as long as in P. porosa. In both species, tissues at and surrounding the lesions contained significantly higher sclerite content per dry weight but lower protein per surface area. In and around the lesion area, the density of symbiotic dinoflagellates, Symbiodinium spp., was lower than in uninjured branches, although Symbiodinium photochemical efficiency in tissues surrounding the lesion was not affected. The gorgonian species differed in their bacterial microbiome, but the overall bacterial community and dominant bacterial taxa did not differ between injured and uninjured branches, although the prevalence of some less common bacterial groups did vary. The two species exhibited distinct immune responses, whereby different components of the melanization cascade were activated, and exochitinase was mobilized only in E. flexuosa. While the gorgonian species differed in their lesion recovery response, both healed without signs of disease or colonization by fouling organisms. The capacity to recover successfully from injuries may partly explain why gorgonian corals dominate Caribbean coral reefs.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In contrast to scleractinian corals whose abundance on Caribbean reefs has dramatically declined over the past few decades (Gardner et al. 2003; Alvarez-Filip et al. 2011; Jackson et al. 2014), gorgonian corals dominate many Caribbean reefs (Goldberg 1973; Kinzie III 1973; Jordán-Dahlgren 1989; Ruzicka et al. 2013; Villamizar et al. 2014; Lenz et al. 2015). They often serve as shelter and a food source for coral reef organisms (Voss 1956; Birkeland and Neudecker 1981; Lasker et al. 1988; Vreeland and Lasker 1989). Despite their dominance, knowledge about gorgonian physiology (e.g., Cary 1918; Kanwisher and Wainwright 1967; Lewis and Post 1982; Ramsby et al. 2014; Shirur et al. 2014) and their microbial consortia (Toledo-Hernández et al. 2008; Sunagawa et al. 2010; Hewson et al. 2011; Duque-Alarcón et al. 2012; Correa et al. 2013; Tracy et al. 2015; McCauley et al. 2016; Robertson et al. 2016) is scant. This paucity of data applies both to ambient and adverse conditions, such as injury, that may compromise gorgonian colonies. Investigating lesion healing in gorgonian corals is one-step toward understanding gorgonian abundance on Caribbean reefs.
Due to the upright arborescent morphology of many gorgonian species and their soft tissue, they can be damaged by wave action such as during storm surges (Cary 1914; Wahle 1985) or entanglement with fishing gear (Chiappone et al. 2005). Injuries can range from mild abrasion to uprooting of a colony and result in partial or whole colony mortality. Predation by butterflyfish, Chaetodon spp. (Birkeland and Neudecker 1981; Lasker 1985), the polychaete, Hermodice sp. (Vreeland and Lasker 1989), and snails, Cyphoma spp. (Lasker et al. 1988; Ruesink and Harvell 1990) also leads to lesions in gorgonian corals. While the butterflyfish feed exclusively on polyps, the worms and snails feed on multiple gorgonian tissues. In turn, pathogens may colonize the wound. The injury site may also serve as a substrate for other species, as in the case of the fire coral Millepora sp., which can overgrow parts or entire colonies of the gorgonian Plexaura homomalla (Wahle 1980; Gerhart 1990).
Growth of new tissue at a lesion site requires organic molecules. In scleractinian corals, these molecules may migrate from adjacent uninjured tissues or other parts of the colony (Oren et al. 2001; Henry and Hart 2005). In gorgonian corals, food particles and organic molecules can be transported within branches (Murdock 1978a, b), but the effects of lesions on the biochemical composition of injured and uninjured branches have not been assessed. Since in gorgonian species, the amount of non-skeletal material (soluble protein, lipids and carbohydrates) per dry weight varies from 4 to 25 % (Shirur et al. 2014), lesion recovery may differ between species if recovery is correlated to tissue content. Furthermore, similar to scleractinian corals, many gorgonian species host endosymbiotic dinoflagellates, Symbiodinium spp. Lesions may affect Symbiodinium photosynthesis or density in gorgonian cells around the lesion site.
Lesions on gorgonian branches may enable pathogens to colonize the injury site. Under ambient, non-stressful conditions, various bacteria, viruses, fungi, and protists inhabit tissues of scleractinian and gorgonian corals (Knowlton and Rohwer 2003; Toledo-Hernández et al. 2008; Sunagawa et al. 2010; Hewson et al. 2011; Burge et al. 2012; Duque-Alarcón et al. 2012; Correa et al. 2013; McCauley et al. 2016; Robertson et al. 2016). In scleractinian corals, bacteria are important in cycling sulfur and nitrogen, and some bacteria may limit the growth of opportunistic taxa (reviewed in Thompson et al. 2015). Diseased tissues of scleractinian and octocorals exhibit a different bacterial community than healthy tissues (Sunagawa et al. 2009; Cárdenas et al. 2012; Vezzulli et al. 2013; Meyer et al. 2014; Roder et al. 2014).
Any biological changes at the injury site may trigger an immune response. Gorgonian corals produce a suite of compounds with antimicrobial and antifungal activity (Perkins and Ciereszko 1973; Rodríguez 1995; Jensen et al. 1996; Kim et al. 2000; Hunt et al. 2012). In addition, amoebocytes in gorgonian corals can phagocytize foreign particles (Olano and Bigger 2000; Ruiz-Diaz et al. 2013) and prevent the entry, or limit proliferation of pathogens by depositing melanin (Petes et al. 2003; Mydlarz et al. 2008). During lesion recovery in several scleractinian corals (D’Angelo et al. 2012; van de Water et al. 2015), and when the gorgonian coral G. ventalina was exposed to fungal pathogens (Mydlarz et al. 2008), the activity of phenoloxidase (PO), the key enzyme of the melanization cascade (Cerenius and Söderhäll 2004), increased. Invading organisms may also be killed by cytotoxic intermediates of the pathway like quinones and reactive oxygen species (Cerenius and Söderhäll 2004). In addition to these general immune responses, fungal pathogens are inhibited by the antioxidant peroxidase (POX) and protease inhibitors (Mydlarz and Harvell 2007; Mann et al. 2014), and the release of exochitinase (EXOC) into the surrounding water (Douglas et al. 2007). In gorgonian corals, it is unknown how the immune defenses react to the occurrence of a lesion. The current study tested the hypothesis whether Caribbean gorgonian branches inflicted with lesions differed from uninjured branches in their biochemical composition, Symbiodinium parameters, microbiome, and immune response.
Methods
Study site and experimental design
This study occurred in June 2012 on a patch reef (20°52′5.23″N, 86°51′58.92″W) near the Instituto Ciencas del Mar y Limnologia in Puerto Morelos, Mexico. At 3–5 m depth, eight colonies each of the gorgonian corals Eunicea flexuosa and Pseudoplexaura porosa were tagged. A branch in each colony was chosen, and to simulate injury, 2 cm below the apical tip, the cortex (the outer tissue region housing the polyps and embedded with sclerites) was removed from one side, exposing the underlying proteinaceous axial rod (Fig. 1a, e). The resulting lesion was 10 cm long and approximately half the width of the branch. A branch in a different area of the same colony was chosen as an uninjured control.

Lesion recovery in photographs of representative branches of the gorgonian corals Eunicea flexuosa (a–d) and Pseudoplexaura porosa (e, f) following artificial injury. The lesion on day 1 (a, e), 4 (b), 7 (c, f) and 14 (d) days post injury. Black arrows in (b) and (c) point to sclerites visible along the lesion perimeter of E. flexuosa
Photochemistry of photosystem II (PSII)
Using a fluorometer (Diving-PAM, Walz, Germany), the maximum (at local dusk: F v/F m) and effective (at local noon: ΔF/F m′) photochemical efficiency of PSII was recorded every 2 days. Photochemical efficiency was measured on the injured branch in tissue adjacent to the top, middle, and lower area of the lesion, and in corresponding regions of the uninjured control branch. From the effective and maximum photochemical efficiencies, the maximum excitation pressure over PSII (Q m) was calculated (Iglesias-Prieto et al. 2004).
Sample collection and processing
Gorgonian branches were collected when, at the lesion site, tissues were flush with surrounding uninjured tissues, and newly regenerated polyps emerged along the perimeter (Fig. 1d, f). This occurred after 7 days in P. porosa and 14 days in E. flexuosa. Measuring from the branch tip, 20-cm-long fragments were cut from both the injured and uninjured branches. These fragments were sub-partitioned, and 2-cm pieces, located 6–8 cm from the branch tips, were removed. The length, diameter, and wet weight of the 2-cm pieces were recorded, and both the 2-cm pieces and the remaining branch segments were frozen in liquid nitrogen and stored at −80 °C until further processing.
Symbiodinium parameters and host protein were quantified from the 2-cm-long pieces. The remaining branch segments were processed for the other assays by separating the cortex from the inner axial rod and discarding the rod. For the injured branch, subsamples were taken from regenerated tissue at the lesion and uninjured tissue on the other side of the branch (8–11 cm from the branch tip), and from tissue at least 2 cm below the lesion (14–15.5 cm from the branch tip). For the uninjured branch, subsamples were taken from 8 to 9.5 cm from the branch tip. Subsamples were freeze-dried prior to analysis of sclerite, protein, carbohydrate, and melanin content.
Determination of Symbiodinium cell density and chlorophyll a (Chl a) and c 2 (Chl c 2) content followed described protocols (Shirur et al. 2014), standardizing the values per surface area, per mg host protein or per symbiont cell. Symbiodinium cells were enumerated using a FlowCAM particle analyzer (Fluid Imaging Technologies Inc., USA). Host protein was quantified from the supernatant in the Symbiodinium isolation protocol (Dove et al. 2006). Sclerite, protein, and carbohydrate content determination followed the protocols described in Shirur et al. (2014), with the contents standardized to dry weight of the cortex (%g DW) (Shirur et al. 2014).
Genetic identification of Symbiodinium and bacteria
DNA was extracted from regenerated tissue at the lesion and the uninjured tissue adjacent to it (on the other side of the branch), and in the uninjured branch from tissues located roughly 4 cm away from the branch tip, following standard protocols (Shirur et al. 2014). The internal transcribed spacer 2 region (ITS2) of the ribosomal DNA was used for Symbiodinium identification (Shirur et al. 2014).
The bacterial microbiome of gorgonian tissues was characterized based on a 250-bp portion of the bacterial 16S rRNA gene using Illumina MiSeq sequencing (Kozich et al. 2013). Samples were pooled, spiked with 5 % PhiX (Jackson et al. 2015), and sequenced at the University of Mississippi Medical Center Molecular and Genomics Core Facility. Raw sequences were processed using the bioinformatics software mothur 1.35.1 (Schloss et al. 2009, 2011) according to the protocol of Kozich et al. (2013). Sequences were aligned against the SILVA 16S rRNA database (Quast et al. 2013), and chimeras detected using UCHIME (Edgar et al. 2011). Valid sequences were classified using the Greengenes 16S rRNA classification scheme (DeSantis et al. 2006). Archaeal and eukaryotic sequences were removed, and the bacterial sequences were grouped into operational taxonomic units (OTUs) based on 97 % sequence similarity.
Melanin content and enzyme activity
Melanin was extracted from the freeze-dried tissues following the Palmer et al. (2011a) protocol and standardized to the amount of organic matter within the sample (Shirur et al. 2014). For the injured branch, enzyme activities were quantified from regenerated tissue at the lesion and uninjured tissue adjacent to the lesion (on the other side of the branch, both 3–6 cm from the branch tip) and from tissues at least 2 cm below the lesion (17–18.5 cm from the branch tip). To quantify enzyme activity, proteins were extracted in 100 mM phosphate buffer pH 7.8 using the Mydlarz and Harvell (2007) protocol.
Total potential phenoloxidase (PO) activity was quantified using the Palmer et al. (2011a) protocol. Peroxidase (POX) activity was obtained by modifying the Mydlarz and Harvell (2007) protocol whereby 20 µl of the extract was diluted with 20 µl of 0.01 mM PBS pH 6.0, and 25 µl of 25 mM guaiacol was added to the dilution. The reaction was initiated with 60 μl of 25 mM H2O2. For PO and POX, the change in absorbance per minute was calculated from the linear portion of the curve. Exochitinase (EXOC) activity was estimated by modifying the Couch et al. (2008) protocol using 60 μl of the diluted extract in the assay. Background fluorescence was quantified in wells containing either the sample or substrate and aliquots of sodium acetate buffer and 0.5 M Na2CO3, and it was subtracted from fluorescence detected in the samples. Enzyme activity assays were run in duplicate and normalized to mg protein present in the aliquot of extract used. Protein content (mg ml−1) of the enzyme extract was determined using the RED660™ Protein Assay Kit (G-Biosciences, USA). PO and EXOC activity were quantified in both species, but POX activity was only measured in E. flexuosa because the mucus in P. porosa interfered with the assay.
Statistical analyses
With the exception of the bacterial community analyses, the two gorgonian species were analyzed individually since the time to lesion closure differed between them. For most comparisons, linear mixed-effects models using the restricted maximum likelihood method were run with the lme4 (Bates et al. 2013) and lmerTest (Kuznetsova et al. 2014) packages within R 3.2.2 (R Core Team 2015). Photochemistry was analyzed with a linear mixed-effects model where treatment (tissue adjacent to the lesion and uninjured branches) and time (days) were the fixed effects and parent colonies were the random effect. A paired Student’s t test was used to test differences in host protein and the Symbiodinium parameters between tissues from the wounded region of the injured branch (which included regenerated tissues at the lesion and uninjured tissue on the other side of the branch) and those from the uninjured branch.
In the linear mixed model used to assess the lesion effect on tissue biochemical composition, melanin content, and enzyme activities, the fixed effect was the experimental treatment (four levels: regenerated tissue at the lesion, uninjured tissue adjacent to it on the other side of the branch, and at least 2 cm below the lesion, and tissue from the uninjured branch), and the parent colony was the random effect. Results of the mixed model analyses were further explored using post hoc planned contrasts with the multcomp package (Hothorn et al. 2008). For each parameter, values at the lesion were compared to those adjacent to it. When significant differences occurred, the tissues adjacent to the lesion were compared to those below it (i.e., uninjured at the lesion area vs. uninjured below lesion area). If the injured and adjacent tissues were not statistically different, they were pooled and compared to tissues sampled from below the lesion. If the three regions in the injured branches did not significantly differ from each other, they were pooled and compared to tissues of the uninjured branch. If tissues at and adjacent to the lesion significantly differed from tissues below it, then the latter was separately compared to tissues from the uninjured branch. The P values for all comparisons were corrected using the Bonferroni method (Westfall 1997).
Bacterial community structure was explored with the theta index of dissimilarity (Yue and Clayton 2005), using mothur 1.35.1. To standardize for different sequence depth between samples, data were randomly subsampled to 425 sequences per sample (the lowest number of valid reads from any sample), and the mean of 1000 subsampling iterations was used for all analyses (Schloss et al. 2011). Analysis of molecular variance (AMOVA) was run on the whole dataset to test whether the bacterial community differed between E. flexuosa and P. porosa, and again for each gorgonian species separately to test whether the community differed between injured and uninjured tissues. Indicator analysis identified bacterial OTUs responsible for the detected differences. Linear mixed-effects models tested the effect of injury on the relative abundance of bacterial taxa (phyla, families, and individual OTUs) in tissues of each gorgonian species, where treatment (three levels: regenerated tissue at the lesion, uninjured tissue adjacent to it on the other side of the branch, and tissue from the uninjured branch) was the fixed effect, and parent colonies were the random effect. To test the effect of injury on the number of OTUs found in tissues, a generalized linear model (using a Poisson distribution) was used.
Results
Lesion recovery
The artificial injury exposed the axial rod to the environment (Fig. 1a, e). In both gorgonian species, 4 days post injury, the axial rod was no longer visible, and tissues at the lesion were violet in contrast to the beige coloration of adjacent uninjured tissues and the rest of the colony. The healing process, however, differed between the two gorgonian species. In E. flexuosa, large violet and white spindle-shaped sclerites were visible in the wound (Fig. 1b, c). These sclerites were subsequently overgrown by tissues (Fig. 1c, d). A week post injury in E. flexuosa, the deep gash in the center of the lesion still remained, and tissue along its perimeter was devoid of polyps (Fig. 1c). The gradual advance of opposing fronts until they fused with each other, and the formation of polyps along the lesion perimeter, took an additional 7 days (Fig. 1d). Conversely, in P. porosa, surface tissues and polyps occurred at the lesion site a week post injury (Fig. 1f), and small dark pigmented sclerites were visible underneath the tissues.
Biochemical composition of tissues
In E. flexuosa, the sclerite content of the cortex in tissues at and surrounding the lesions was significantly greater than in tissues below the lesions and in the uninjured branches [Fig. 2a, ANOVA, F(3, 15) = 4.0, P = 0.029]. In contrast, sclerite content in P. porosa did not differ between the three regions of the injured branches, but was significantly higher in the injured than in the uninjured branches [Fig. 2a, ANOVA, F(3, 14) = 24.9, P < 0.001]. The mean protein content of the cortex did not differ between the four sampled regions (injured branch: injured, adjacent uninjured and below uninjured, and the uninjured branch) in E. flexuosa [ANOVA, F(3, 15) = 3.1, P = 0.060] and P. porosa [ANOVA, F(3, 14.1) = 2.7, P = 0.089], although a trend existed whereby the protein content was lower in the injured than the uninjured branches (Fig. 2b). For both species, the areal content of host protein was significantly lower in tissues at and adjacent to the lesion than in the uninjured branches (Table 1). Conversely, carbohydrate content of the cortex in both species did not differ between the injured and uninjured branch regions (Table 1).

Mean a sclerite and b protein contents per dry weight (%g DW), and activities of the enzymes, c total potential prophenoloxidase (PO) and d exochitinase (EXOC) in tissues of the gorgonian corals Eunicea flexuosa and Pseudoplexaura porosa. Samples (see inset) were taken from uninjured branches (Uninjured) and three regions of the injured branches (Injured): branch tissue away from the lesion (Away), uninjured tissue adjacent to the lesion (Near), and regenerated tissue at the lesion (Lesion). Data are mean ± SE with n = 6 for most data and n = 5 for P. porosa-Uninjured. Letters (e, f for E. flexuosa and r, s for P. porosa) above the bars denote significant differences (P < 0.05) in the planned contrasts
Melanin content and enzyme activity
Since sclerite, and thus organic content, significantly differed between the injured and uninjured branches, the effect of tissue injury on melanin content was only tested within sampled regions of the injured branches. In E. flexuosa, tissues at the lesion contained significantly less melanin in organic matter (%g OM) than the tissues surrounding and those located below the lesion (Table 1). Conversely, in P. porosa, melanin content did not significantly differ between the three regions of the injured branches (Table 1). The two species also exhibited contrasting trends in enzyme activity. PO and POX activities in E. flexuosa were significantly higher [PO: Fig. 2c, ANOVA, F(3, 15) = 16.3, P < 0.001; POX: Table 1], while in P. porosa, PO activity was significantly lower in tissues at and surrounding the lesion compared to tissues from below the lesion or from uninjured branches [Fig. 2c, ANOVA, F(3, 14) = 12, P < 0.001]. In E. flexuosa, EXOC activity was significantly lower in tissues at and surrounding the lesions compared to in tissues from below the lesions and from the uninjured branches [Fig. 2d, ANOVA, F(3, 15) = 5.4, P = 0.010]. In contrast, in P. porosa, EXOC activity did not significantly differ between injured and uninjured branches [Fig. 2d, ANOVA, F(3, 14.1) = 1.4, P = 0.291].
Symbiodinium parameters
Several of the Symbiodinium in this study were novel and ascribed to the new types B41a (GenBank Accession No. KX344964), B41b (GenBank accession no. KX344965), and B42 (GenBank Accession No. KX344981). Of the eight sampled E. flexuosa colonies, one hosted Symbiodinium type B41a, while seven contained B41b. In P. porosa, six colonies contained Symbiodinium type B1i (Finney et al. 2010; GenBank Accession No. GU907636), and two colonies contained type B42. Since different Symbiodinium exhibit different physiologies (Goulet et al. 2005; Ramsby et al. 2014), we excluded from the statistical analyses, the one and two colonies containing other Symbiodinium types. In both gorgonian species, the Symbiodinium type found in the healed tissues, the tissues surrounding the lesion site, and in the uninjured branches was the same.
In both E. flexuosa and P. porosa, Symbiodinium density (Fig. 3a, Paired t test, E. flexuosa: t6 = 10.7, P < 0.001; P. porosa: t5 = 3.4, P = 0.020) and Chl a (Fig. 3c, Paired t test, E. flexuosa: t6 = 7.0, P < 0.001; P. porosa: t5 = 3.9, P = 0.012) and c 2 (Table 2) per surface area were significantly less in tissues at and those surrounding the lesion than in the uninjured branches. Similar patterns occurred when symbiont density (Fig. 3b, Paired t test, E. flexuosa: t6 = 4.0, P = 0.007; P. porosa: t5 = 2.0, P = 0.105) and Chl a and c 2 contents (Table 2) were expressed per host protein, although in P. porosa, the difference was not statistically significant. In both species, the Chl a (Fig. 3d, Paired t test, E. flexuosa: t6 = −0.4, P = 0.740; P. porosa: t5 = −0.9, P = 0.423) and c 2 content per Symbiodinium cell and the ratio of Chl a: c 2 did not significantly differ between the lesion site, surrounding tissues, and the uninjured branches (Table 2).

Mean Symbiodinium density a per surface area and b per mg host protein, and chlorophyll a content, c per surface area and d per Symbiodinium cell, in the gorgonian corals Eunicea flexuosa and Pseudoplexaura porosa. Samples were taken from uninjured branches (Uninjured) and from areas in the injured branch containing the lesion on one side and uninjured tissue on the other (Injured). Data are mean ± SE, with n = 7 for E. flexuosa and n = 6 for P. porosa. Statistically significant results (P < 0.05) detected by paired t tests are denoted with (asterisk)
F v/F m, ΔF/F m′, and Q m did not significantly differ between the lower, middle, and upper regions of branches for either species. Therefore, the values from the three regions were pooled. During lesion recovery, F v/F m, ΔF/F m′, and Q m in both gorgonian species did not significantly differ between tissues surrounding the lesion and those from the uninjured branches (Online Resources 1 and 2).
Bacterial community
The two gorgonian species harbored distinct bacterial communities, with 160,736 (mean: 8113, median: 7789) and 137,924 (mean: 9455, median: 5063) bacterial sequences isolated from E. flexuosa and P. porosa, respectively. Bacterial sequence data are available in the NCBI Sequence Reads Archive under accession SRP076113. In uninjured branches of E. flexuosa, the majority (73 %) of bacterial sequences could not be classified, while Proteobacteria comprised 21 % of the sequences (Fig. 4a; Table 3). In P. porosa, the phyla Tenericutes (77 %) and Proteobacteria (15 %) accounted for most of the bacterial community in uninjured branches (Fig. 4a; Table 3). In both gorgonian species, other phyla individually comprised <2 % of the bacterial community (Online Resource 3).

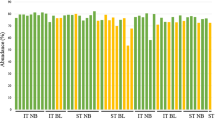
Prevalence (%) of major bacterial phyla within the microbiome (a), bacterial families Hahellaceae (b) and Enterobacteriaceae (c) within Gammaproteobacteria, and the family Bradyrhizobiaceae (d) within Alphaproteobacteria, in tissues of the gorgonian corals Eunicea flexuosa and Pseudoplexaura porosa. Samples were taken from uninjured branches (Uninjured) and two regions of injured branches (Injured): uninjured tissue adjoining the lesion (Near), and regenerated tissue at the lesion (Lesion). Data are presented as mean ± SE, with n = 5 for most parameters and n = 6 for E. flexuosa-Lesion and P. porosa-Near and Lesion in (c), and E. flexuosa-Near and P. porosa-Lesion in (d). Letters (e–g for E. flexuosa and r, s for P. porosa) above the bars denote significant differences (P < 0.05) in the planned contrasts
The prevalence of Tenericutes in P. porosa, and Proteobacteria in E. flexuosa, did not differ between injured and uninjured branches (Fig. 4a; Table 3). In contrast, Proteobacteria in P. porosa were significantly less prevalent at the lesion and adjacent tissues than in the uninjured branch (Fig. 4a; Table 3). Within Proteobacteria, in both gorgonian species, the class Gammaproteobacteria (Table 3), specifically the family Hahellaceae (which in the literature sometimes appears as Endozoicimonaceae, Neave et al. 2016), were significantly less prevalent in tissues at and surrounding the lesion, than in uninjured branches [Fig. 4b, ANOVA, E. flexuosa: F(2, 8) = 20, P < 0.001; P. porosa: F(2, 9) = 18.2, P < 0.001]. Furthermore, Hahellaceae in E. flexuosa were significantly less prevalent in tissues at than in those surrounding the lesion (Fig. 4b). In contrast, the family Enterobacteriaceae in both gorgonian species [Fig. 4c, ANOVA, E. flexuosa: F(2, 8.3) = 11.1, P = 0.005; P. porosa: F(2, 9.1) = 8.5, P = 0.008] and Pseudomonadaceae in P. porosa (Table 3) were significantly more prevalent in injured than in uninjured branches. Pseudomonadaceae and the class Alphaproteobacteria in E. flexuosa were significantly more prevalent in tissues at than in tissues surrounding the lesion and from uninjured branches (Table 3). Within the Alphaproteobacteria, in both gorgonian corals, the prevalence of families Rhodobacteraceae, Methylobacteriaceae, and Hyphomicrobiaceae did not significantly differ between injured and uninjured branches (Table 3). In P. porosa, the family Bradyrhizobiaceae was significantly more prevalent in injured than in uninjured branches [Fig. 4d, ANOVA, E. flexuosa: F(2, 9.2) = 0.1, P = 0.895; P. porosa: F(2, 8.9) = 4.4, P = 0.047].
At a finer taxonomic resolution 1818, OTUs were identified and a significantly different bacterial consortium was found associated with each species [Fig. 5, AMOVA, F(1, 33) = 82, P < 0.001]. Certain OTUs occurred in one gorgonian species and not the other (Fig. 5), while others were found in both species (Table 4). In both gorgonian species, the bacterial community structure [AMOVA, E. flexuosa: F(2, 16) = 0.4, P = 0.813; P. porosa: F(2, 16) = 0.4, P = 0.760] and the prevalence of dominant OTUs 2 and 3 (Unclassified bacteria) in E. flexuosa, and OTU 1 (Mycoplasma sp.) and OTU 3 in P. porosa, did not significantly differ between injured and uninjured branches (Table 4). In E. flexuosa, tissues at the lesion site contained significantly more OTUs than tissues surrounding the lesion and in the uninjured branches, while in P. porosa, the number of OTUs did not significantly vary between sampled tissues (Table 4).

Non-metric multidimensional scaling ordination plot generated using the OTUs identified in the gorgonian corals Eunicea flexuosa (filled symbols) and Pseudoplexaura porosa (open symbols). Samples were taken from uninjured branches (squares) and two regions of injured branches: uninjured tissue adjoining the lesion (circles), and regenerated tissue at the lesion (triangles). Dotted arrows indicate OTUs that primarily associated with one or the other gorgonian species
Discussion
Gorgonian corals dominate many Caribbean coral reefs (Goldberg 1973; Kinzie III 1973; Jordán-Dahlgren 1989; Ruzicka et al. 2013), and how these corals respond to injury may help explain their abundance. Wound healing can vary based on lesion characteristics (Cary 1914; Lang da Silveira and Van’t Hof 1977; Meesters et al. 1997; Oren et al. 1997; van Woesik 1998) and environmental conditions (Wahle 1983; Henry and Hart 2005; Denis et al. 2011; Sabine et al. 2015). Since in our study, these variables were similar for both E. flexuosa and P. porosa, the different recovery rates likely reflect interspecies variability in recovery similar to the observed ranges of 6–8 days in E. mammosa, 7–9 days in Plexaura homomalla, and 7–11 days in Plexaurella dichotoma (Wahle 1983). Furthermore, although E. flexuosa and P. porosa both belong to the family Plexauridae (Wirshing et al. 2005; Daly et al. 2007), they may be quite different from one another. In chemotaxonomic and phylogenetic studies, Eunicea spp. clustered separately from Pseudoplexaura spp., but since the techniques used could only resolve clade-level differences, the clusters were not significantly different (Gerhart 1983; Sánchez et al. 2003; Wirshing et al. 2005). In another study, on the phylogenetic relationships between Eunicea species, where finer resolution was possible with the ITS2 marker, Pseudoplexaura crucis, which is in the same genus as P. porosa (Wirshing et al. 2005), was used as an outgroup, demonstrating the differences between Eunicea spp. and Pseudoplexaura spp. (Grajales et al. 2007). Therefore, taxonomic separation may contribute to the differences between E. flexuosa and P. porosa including why lesion recovery in E. flexuosa took twice as long as in P. porosa, although recovery in E. flexuosa was comparable to an earlier study on that species (Lang da Silveira and Van’t Hof 1977).
Energetic sources for lesion recovery
E. flexuosa and P. porosa also differ in their biochemical composition. The combined sclerite and refractory content in E. flexuosa (81 and 13 % of dry weight) is higher than that in P. porosa (46 % sclerite and 29 % refractory content per dry weight) (Shirur et al. 2014). Consequently, protein, lipid, and carbohydrate content together average just 6 % of the dry weight of E. flexuosa compared to over four times as much (25 %) in P. porosa (Shirur et al. 2014). Using the mean protein, lipid and carbohydrate content per organic matter (Shirur et al. 2014), and their enthalpies of combustion (Gnaiger and Bitterlich 1984), we estimate that tissues in E. flexuosa contain almost 40 % less energy than those in P. porosa (9650 vs. 15,730 J g−1 OM). Furthermore, E. flexuosa polyps capture fewer and smaller prey than P. porosa polyps (Ribes et al. 1998).
In addition, Symbiodinium photosynthesis in E. flexuosa may be lower than that in P. porosa. The net photosynthetic rate of Symbiodinium in E. tourneforti, a species with a biochemical composition similar to E. flexuosa (Shirur et al. 2014), is almost three times lower than in P. porosa (Ramsby et al. 2014). This could be due to genetic differences between the gorgonians (Gerhart 1983; Wirshing et al. 2005; Grajales et al. 2007) as well as different Symbiodinium types (Ramsby et al. 2014). Besides photosynthetic differences at ambient conditions, biochemical changes in branches during lesion recovery could affect Symbiodinium in surrounding tissues. Diversion of organic molecules by the host toward lesion recovery could reduce the host’s regular translocation of inorganic carbon, fatty acids, and nitrogenous compounds to the Symbiodinium (Hughes et al. 2010; Imbs et al. 2014; Tanaka et al. 2015), subsequently affecting photosynthesis and the routine transfer of photosynthetic products to the host (Kopp et al. 2015). Symbiodinium in tissues near the lesion, however, did not seem to be affected since their photochemical parameters, chlorophyll content of their cells, and the Chl a:c 2 ratio did not differ from those in the uninjured branches. Similarly, in the scleractinian corals Pocillipora verrucosa and Acropora muricata, the maximum quantum yield of photosystem II did not differ between tissues surrounding a lesion and those in uninjured branches (Lenihan and Edmunds 2010; Denis et al. 2013).
Conversely, Symbiodinium density was lower in tissues near the lesion than in uninjured branches. In scleractinian corals, Symbiodinium density depends on the host biomass available to harbor the algae (Drew 1972; Jones and Yellowlees 1997; Thornhill et al. 2012). In our study, in both gorgonian corals, sclerite content was significantly higher and host protein content per surface area was significantly lower at the lesion and in surrounding tissues than in uninjured branches, indicating that the regions near the lesion contained less host biomass. Therefore, fewer Symbiodinium-containing host cells in the injured gorgonian branches may have led to the lower Symbiodinium density. Alternatively, lower Symbiodinium density could occur because the remaining host cells each contained fewer Symbiodinium. Symbiodinium density per host protein was lower in injured branches of E. flexuosa and P. porosa, although this difference was significant only in E. flexuosa. Thus, the longer recovery time in E. flexuosa compared to P. porosa may reflect a combination of less protein, lipids, and carbohydrates and hence existing energy reserves, and lower amounts of heterotrophically and photosynthetically acquired nutrients.
The process of lesion recovery and its implications
Although the rate of recovery differed between the two species, sclerites were visible at the lesion sites in E. flexuosa and P. porosa, and sclerite content in both species was significantly higher at the lesion and in surrounding tissues than in uninjured branches. This suggests that sclerites are integral to lesion recovery. Consequently, conditions that detrimentally affect sclerites may adversely affect wound healing. For example, in scleractinian corals, low pH hinders calcification (Hoegh-Guldberg et al. 2007; Anthony et al. 2011; Dove et al. 2013). Potentially, because sclerites are embedded within tissues, low pH conditions did not affect sclerites in the gorgonian corals E. flexuosa (Enochs et al. 2016), E. fusca (Gómez et al. 2015) and in the octocoral Ovabunda macrospiculata (Gabay et al. 2014). The lack of effect on sclerites could be one of the reasons for a transition from scleractinian coral to octocoral-dominated benthos closer to naturally occurring volcanically acidified water (Inoue et al. 2013). On the other hand, when sclerites were removed from tissues of the octocoral O. macrospiculata, thereby directly exposing them to lower pH, partial sclerite dissolution did occur (Gabay et al. 2014). Ocean acidification could therefore detrimentally affect exposed sclerites at lesion sites and hinder wound healing in gorgonian corals, potentially, similar to the effect of lower pH on wound healing in scleractinian corals (Horwitz and Fine 2014; Hall et al. 2015).
The bacterial community in injured and uninjured tissues
In addition to hosting Symbiodinium spp. (zooxanthellae), scleractinian corals and octocorals contain a consortium of organisms and their microbiome (Knowlton and Rohwer 2003; Toledo-Hernández et al. 2008; Sunagawa et al. 2010; Hewson et al. 2011; Thompson et al. 2015). Since the bacterial portion of this microbiome was unknown for E. flexuosa and P. porosa, we characterized these communities. In E. flexuosa, the majority of bacteria were identified as two OTUs (OTU 2, 3) that could be classified only to Bacteria, while in P. porosa, the majority of bacteria belonged to Tenericutes (OTU 1: Mycoplasma spp.). Tenericutes are host-specific commensals or parasites (Razin et al. 1998) which can be major components of the bacterial microbome in azooxanthellate octocorals (Gray et al. 2011; Porporato et al. 2013; Holm and Heidelberg 2016) and the azooxanthellate scleractinian coral Lophelia pertusa (Kellogg et al. 2009). They are rare or absent in zooxanthellate gorgonian and scleractinian corals in which Proteobacteria dominate the bacterial community (Sunagawa et al. 2010; Cárdenas et al. 2012; Duque-Alarcón et al. 2012; Lee et al. 2012; Correa et al. 2013; Tracy et al. 2015; McCauley et al. 2016; Robertson et al. 2016).
Proteobacteria was the second most abundant phylum in both E. flexuosa and P. porosa, represented predominantly by the Gammaproteobacteria and Alphaproteobacteria classes. Among Gammaproteobacteria, most were Hahellaceae, a group that may be part of the core microbiome of scleractinian corals and octocorals (Bayer et al. 2013; Correa et al. 2013; La Rivière et al. 2013, 2015; McCauley et al. 2016; Neave et al. 2016; Robertson et al. 2016). The Hahellaceae in our samples were most closely related to Endozoicomonas euniceicola and E. gorgoniicola identified from the Caribbean gorgonian corals Eunicea fusca and Plexaura sp., respectively (Pike et al. 2013), E. montiporae and E. elysicola isolated from the scleractinian coral Montipora aequituberculata (Yang et al. 2010) and the slug Elysia ornata (Kurahashi and Yokota 2007), respectively, from the Indo-Pacific.
Other Gammaproteobacteria were identified as Pseudomonadaceae, which have been previously found in octocorals and scleractinian corals (Penn et al. 2006; Duque-Alarcón et al. 2012; Lee et al. 2012; Morrow et al. 2012; Bourne et al. 2013; Correa et al. 2013). Lastly, 1.5 % of Gammaproteobacteria in E. flexuosa and 4.2 % in P. porosa were Enterobacteriaceae, including Escherichia coli. The E. coli source could be runoff from housing along the coastline, or from contaminated groundwater (Baker et al. 2010; Leal-Bautista et al. 2011), which around the town Puerto Morelos may reach the ocean through underwater springs (Carruthers et al. 2005). Among the Alphaproteobacteria, two orders, Rhodobacterales (primarily Rhodobacteraceae) and Rhizobiales (Methylobacteriaceae, Hyphomicrobiaceae and Bradyrhizobiaceae), were the most prevalent, both of which have been reported in octocorals and scleractinian corals (Penn et al. 2006; Kellogg et al. 2009; Mouchka et al. 2010; Gray et al. 2011; Duque-Alarcón et al. 2012; Morrow et al. 2012; Bourne et al. 2013; Lema et al. 2014; Ainsworth et al. 2015; Robertson et al. 2016). In E. flexuosa and P. porosa, 9 and 73 % of the Alphaproteobacteria could not be classified further (Online Resource 4).
Injury did not cause a change in the overall structure of the bacterial microbiome of E. flexuosa and P. porosa, a finding similar to that in the scleractinian coral Acropora aspera (van de Water et al. 2015). The prevalence of some individual bacterial taxa did differ between injured and uninjured branches. Hahellaceae were 2.4× and 1.6× less prevalent in tissues at and adjacent to the lesion in E. flexuosa than in uninjured branches, and in injured branches of P. porosa, they were 3× less prevalent. A reduction in the prevalence of Hahellaceae also occurred in corals sampled from sites exposed to human impacts (Vezzulli et al. 2013; Roder et al. 2015; Ziegler et al. 2016), acidified conditions (Morrow et al. 2015), and in corals exhibiting bleaching (Bourne et al. 2008) or tissue lesions (Meyer et al. 2014).
In contrast, the proportion of E. coli was significantly higher (9.8× for E. flexuosa, 4.8× for P. porosa) at sites at and surrounding the lesion. A similar pattern presented itself for Pseudomonadaceae in E. flexuosa at the lesion site (2.8×) and in P. porosa (4.4×) in tissues at and surrounding the lesion. Increases in the prevalence of Pseudomonadaceae and Rhizobiales also occurred in white plague diseased tissues of scleractinian corals, but these bacteria may be opportunistic rather than the disease-causing pathogens (Sunagawa et al. 2009; Mouchka et al. 2010; Cárdenas et al. 2012; Roder et al. 2014). In addition, in E. flexuosa, the injured sites had a more diverse range of bacterial OTUs (1.6×). In P. porosa, Bradyrhizobiaceae were more prevalent (5.4×) near and at the lesion site, and injury-induced shifts also occurred in some less prevalent (≤1 % of the bacterial community) Gammaproteobacteria and Alphaproteobacteria taxa (Online Resources 4, 5). Never the less, given that tissue recovery proceeded in both E. flexuosa and P. porosa without symptoms of disease, the two species coped with the lesions and the bacteria present in the injured area, although their coping mechanisms may differ.
Defense mechanisms against pathogens
In E. flexuosa, even though some bacterial groups were more abundant at the lesion than in surrounding tissues, the lack of a major bacterial community shift could have been due to immune activity. Defensive mechanisms such as the nonspecific immune responses melanin and PO may protect corals against pathogens (Toledo-Hernández and Ruiz-Diaz 2014). In E. flexuosa, the lesion site and the surrounding tissues had lower melanin content and higher PO and POX activity compared to levels and activity away from the lesion, indicating that the melanization cascade was activated. Melanin may have been released from cells to aid in clot formation as seen in the scleractinian coral Porites cylindrica (Palmer et al. 2011b), or to coat pathogens to limit their proliferation at the lesion as in the gorgonian G. ventalina (Petes et al. 2003; Mydlarz et al. 2008). In addition, high PO activity may produce additional melanin, and intermediate reactive oxygen species (ROS) and quinones that are cytotoxic to pathogens (Cerenius and Söderhäll 2004; Cerenius et al. 2008). High POX activity may protect tissues from ROS produced by the action of PO (Mydlarz and Palmer 2011), and aid in preventing fungal infection at the lesion site (Mydlarz and Harvell 2007). While melanin and PO are nonspecific immune responses against pathogens in general, EXOC specifically breaks down fungal cell walls. In E. flexuosa, EXOC activity was significantly lower at the lesion and in surrounding tissues than in tissues away from the lesion, and therefore, as in G. ventalina, EXOC may have been released from tissues to attack fungi in the water surrounding the lesion (Douglas et al. 2007).
Similar to E. flexuosa, even though in P. porosa the proportion of some minor components of the bacterial microbiome were different in injured versus uninjured branches, the overall bacterial community did not change. In contrast to E. flexuosa, P. porosa did not exhibit activation of many parameters associated with an immune response. Melanin content did not vary, and PO activity was significantly lower in tissues at and surrounding the lesion than further away, suggesting that fewer components of the melanization cascade were activated. Furthermore, EXOC activity did not differ between sampled tissues. Therefore, P. porosa may employ alternate defense mechanisms (Perkins and Ciereszko 1973; Rodríguez 1995; Jensen et al. 1996; Hunt et al. 2012). Tissue extracts of gorgonian species, including E. flexuosa and P. porosa, contain substances with antimicrobial activity (Jensen et al. 1996). In addition, gorgonian compounds may alter quorum sensing between bacteria (Hunt et al. 2012) which could affect bacterial virulence (Waters and Bassler 2005; Rutherford and Bassler 2012), although in gorgonian corals, the mechanism of these defenses is unknown. The composition of such substances may be different and/or the amount may be higher in P. porosa than in E. flexuosa. For example, P. porosa tissue extracts are significantly more potent against the fungal pathogen Aspergillus sydowii than those from E. flexuosa (Kim et al. 2000), and P. porosa tissues contain crassin acetate, which is toxic to ciliates (Perkins and Ciereszko 1973).
Conclusion
The Caribbean gorgonian corals E. flexuosa and P. porosa dealt with artificially induced lesions, and the wounds healed with no visible signs of infection or necrosis. The two gorgonian species, however, differed in how they coped with the lesions, probably due to the structural, biochemical, and symbiotic differences between the two. E. flexuosa and P. porosa also differed in their ambient bacterial consortia and in their immune response. These multiple distinctions illustrate the importance of investigating numerous gorgonian species. Despite their differences, both P. porosa and E. flexuosa handled the potential stress brought about by injury. The capacity to recover from this stressor may in part explain why gorgonian corals are maintaining and/or increasing in abundance on Caribbean reefs.
References
Ainsworth TD, Krause L, Bridge T, Torda G, Raina J-B, Zakrzewski M, Gates RD, Padilla-Gamiño JL, Spalding HL, Smith C, Woolsey ES, Bourne DG, Bongaerts P, Hoegh-Guldberg O, Leggat W (2015) The coral core microbiome identifies rare bacterial taxa as ubiquitous endosymbionts. ISME J 9:2261–2274. doi:10.1038/ismej.2015.39
Alvarez-Filip L, Côté IM, Gill JA, Watkinson AR, Dulvy NK (2011) Region-wide temporal and spatial variation in Caribbean reef architecture: is coral cover the whole story? Glob Change Biol 17:2470–2477. doi:10.1111/j.1365-2486.2010.02385.x
Anthony KRN, Maynard JA, Diaz-Pulido G, Mumby PJ, Marshall PA, Cao L, Hoegh-Guldberg O (2011) Ocean acidification and warming will lower coral reef resilience. Glob Change Biol 17:1798–1808. doi:10.1111/j.1365-2486.2010.02364.x
Baker DM, Jordán-Dahlgren E, Angel Maldonado M, Harvell CD (2010) Sea fan corals provide a stable isotope baseline for assessing sewage pollution in the Mexican Caribbean. Limnol Oceanogr 55:2139–2149. doi:10.4319/lo.2010.55.5.2139
Bates D, Maechler M, Bolker B, Walker S (2013) lme4: Linear mixed-effects models using Eigen and S4. R package version 1.1
Bayer T, Neave MJ, Alsheikh-Hussain A, Aranda M, Yum LK, Mincer T, Hughen K, Apprill A, Voolstra CR (2013) The microbiome of the Red Sea coral Stylophora pistillata is dominated by tissue-associated Endozoicomonas bacteria. Appl Environ Microbiol 79:4759–4762. doi:10.1128/AEM.00695-13
Birkeland C, Neudecker S (1981) Foraging behavior of two Caribbean chaetodontids: Chaetodon capistratus and C. aculeatus. Copeia 1:169–178. doi:10.2307/1444051
Bourne DG, Iida Y, Utchike S, Smith-Keune C (2008) Changes in coral-associated microbial communities during a bleaching event. ISME J 2:350–363. doi:10.1038/ismej.2007.112
Bourne DG, Dennis PG, Uthicke S, Soo RM, Tyson GW, Webster N (2013) Coral reef invertebrate microbiomes correlate with the presence of photosymbionts. ISME J 7:1452–1458. doi:10.1038/ismej.2012.172
Burge CA, Douglas NL, Conti-Jerpe I, Weil E, Roberts S, Friedman CS, Harvell CD (2012) Friend or foe: the association of Labyrinthulomycetes with the Caribbean sea fan Gorgonia ventalina. Dis Aquat Org 101:1–12. doi:10.3354/dao02487
Cárdenas A, Rodriguez-R LM, Pizarro V, Cadavid LF, Arévalo-Ferro C (2012) Shifts in bacterial communities of two Caribbean reef-building coral species affected by white plague disease. ISME J 6:502–512. doi:10.1038/ismej.2011.123
Carruthers TJB, van Tussenbroek BI, Dennison WC (2005) Influence of submarine springs and wastewater on nutrient dynamics of Caribbean seagrass meadows. Estuar Coast Shelf Sci 64:191–199. doi:10.1016/j.ecss.2005.01.015
Cary LR (1914) Observations upon the growth-rate and oecology of gorgonians. Publ Carnegie Inst Wash 5:81–92
Cary LR (1918) A study of respiration in Alcyonaria. Publ Carnegie Inst Wash 12:185–191
Cerenius L, Söderhäll K (2004) The prophenoloxidase-activating system in invertebrates. Immunol Rev 198:116–126. doi:10.1111/j.0105-2896.2004.00116.x
Cerenius L, Lee BL, Söderhäll K (2008) The proPO-system: pros and cons for its role in invertebrate immunity. Trends Immunol 29:263–271. doi:10.1016/j.it.2008.02.009
Chiappone M, Dienes H, Swanson DW, Miller SL (2005) Impacts of lost fishing gear on coral reef sessile invertebrates in the Florida Keys National Marine Sanctuary. Biol Conserv 121:221–230. doi:10.1016/j.biocon.2004.04.023
Correa H, Haltli B, Duque C, Kerr R (2013) Bacterial communities of the gorgonian octocoral Pseudopterogorgia elisabethae. Microb Ecol 66:972–985. doi:10.1007/s00248-013-0267-3
Couch CS, Mydlarz LD, Harvell CD, Douglas NL (2008) Variation in measures of immunocompetence of sea fan coral, Gorgonia ventalina, in the Florida Keys. Mar Biol 155:281–292. doi:10.1007/s00227-008-1024-x
Daly M, Brugler M, Cartwright P, Collins AG, Dawson M, Fautin DG, France SC, McFadden CS, Opresko DM, Rodriguez E, Romano SL, Stake JL (2007) The phylum Cnidaria: a review of phylogenetic patterns and diversity 300 years after Linnaeus. In: Zhang Z-Q, Shear WA (eds) Linnaeus tercentenary: progress in invertebrate taxonomy. Magnolia Press, Auckland, pp 127–182
D’Angelo C, Smith E, Oswald F, Burt J, Tchernov D, Wiedenmann J (2012) Locally accelerated growth is part of the innate immune response and repair mechanisms in reef-building corals as detected by green fluorescent protein (GFP)-like pigments. Coral Reefs 31:1045–1056. doi:10.1007/s00338-012-0926-8
Denis V, Debreuil J, De Palmas S, Richard J, Guillaume M, Bruggemann JH (2011) Lesion regeneration capacities in populations of the massive coral Porites lutea at Réunion Island: environmental correlates. Mar Ecol Prog Ser 428:105–117. doi:10.3354/meps09060
Denis V, Guillaume MMM, Goutx M, de Palmas S, Debreuil J, Baker AC, Boonstra RK, Bruggemann JH (2013) Fast growth may impair regeneration capacity in the branching coral Acropora muricata. PLoS One 8:e72618. doi:10.1371/journal.pone.0072618.t001
DeSantis TZ, Hugenholtz P, Larsen N, Rojas M, Brodie EL, Keller K, Huber T, Dalevi D, Hu P, Andersen GL (2006) Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl Environ Microbiol 72:5069–5072. doi:10.1128/AEM.03006-05
Douglas N, Mullen K, Talmage S, Harvell CD (2007) Exploring the role of chitinolytic enzymes in the sea fan coral, Gorgonia ventalina. Mar Biol 150:1137–1144. doi:10.1007/s00227-006-0444-8
Dove SG, Ortiz JC, Enríquez S, Fine M, Fisher PL, Iglesias-Prieto R, Thornhill DJ, Hoegh-Guldberg O (2006) Response of holosymbiont pigments from the scleractinian coral Montipora monasteriata to short-term heat stress. Limnol Oceanogr 51:1149–1158
Dove SG, Kline DI, Pantos O, Angly FE, Tyson GW, Hoegh-Guldberg O (2013) Future reef decalcification under a business-as-usual CO2 emission scenario. Proc Natl Acad Sci USA 110:15342–15347. doi:10.1073/pnas.1302701110
Drew EA (1972) The biology and physiology of alga-invertebrates symbioses. II. The density of symbiotic algal cells in a number of hermatypic hard corals and alcyonarians from various depths. J Exp Mar Biol Ecol 9:71–75. doi:10.1016/0022-0981(72)90008-1
Duque-Alarcón A, Santiago-Vásquez LZ, Kerr RG (2012) A microbial community analysis of the octocoral Eunicea fusca. Electron J Biotechnol 15:1–9. doi:10.2225/vol15-issue5-fulltext-11
Edgar RC, Haas BJ, Clemente JC, Quince C, Knight R (2011) UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27:2194–2200. doi:10.1093/bioinformatics/btr381
Enochs IC, Manzello DP, Wirshing HH, Carlton R, Serafy J (2016) Micro-CT analysis of the Caribbean octocoral Eunicea flexuosa subjected to elevated pCO2. ICES J Mar Sci 73:910–919. doi:10.1093/icesjms/fsv159
Finney JC, Pettay DT, Sampayo EM, Warner ME, Oxenford HA, LaJeunesse TC (2010) The relative significance of host–habitat, depth, and geography on the ecology, endemism, and speciation of coral endosymbionts in the genus Symbiodinium. Microb Ecol 60:250–263. doi:10.1007/s00248-010-9681-y
Gabay Y, Fine M, Barkay Z, Benayahu Y (2014) Octocoral tissue provides protection from declining oceanic pH. PLoS One 9:e91553. doi:10.1371/journal.pone.0091553.t001
Gardner TA, Côté IM, Gill JA, Grant A, Watkinson AR (2003) Long-term region-wide declines in Caribbean corals. Science 301:958–960. doi:10.1126/science.1086050
Gerhart DJ (1983) The chemical systematics of colonial marine animals: an estimated phylogeny of the order Gorgonacea based on terpenoid characters. Biol Bull 164:71–81. doi:10.2307/1541191
Gerhart DJ (1990) Fouling and gastropod predation: consequences of grazing for a tropical octocoral. Mar Ecol Prog Ser 62:103–108
Gnaiger E, Bitterlich G (1984) Proximate biochemical composition and caloric content calculated from elemental CHN analysis: a stoichiometric concept. Oecologia 62:289–298. doi:10.1007/BF00384259
Goldberg WM (1973) The ecology of the coral-octocoral communities off the southeast Florida coast: geomorphology, species composition, and zonation. Bull Mar Sci 23:465–488
Gómez CE, Paul VJ, Ritson-Williams R, Muehllehner N, Langdon C, Sánchez JA (2015) Responses of the tropical gorgonian coral Eunicea fusca to ocean acidification conditions. Coral Reefs 34:451–460. doi:10.1007/s00338-014-1241-3
Goulet TL, Cook CB, Goulet D (2005) Effect of short-term exposure to elevated temperatures and light levels on photosynthesis of different host-symbiont combinations in the Aiptasia pallida/Symbiodinium symbiosis. Limnol Oceanogr 50:1490–1498. doi:10.4319/lo.2005.50.5.1490
Grajales A, Aguilar C, Sánchez JA (2007) Phylogenetic reconstruction using secondary structures of Internal Transcribed Spacer 2 (ITS2, rDNA): finding the molecular and morphological gap in Caribbean gorgonian corals. BMC Evol Biol 7:90. doi:10.1186/1471-2148-7-90
Gray MA, Stone RP, McLaughlin MR, Kellogg CA (2011) Microbial consortia of gorgonian corals from the Aleutian Islands. FEMS Microbiol Ecol 76:109–120. doi:10.1111/j.1574-6941.2010.01033.x
Hall ER, DeGroot BC, Fine M (2015) Lesion recovery of two scleractinian corals under low pH conditions: implications for restoration efforts. Mar Pollut Bull 100:321–326. doi:10.1016/j.marpolbul.2015.08.030
Henry L-A, Hart M (2005) Regeneration from injury and resource allocation in sponges and corals—a review. Int Rev Hydrobiol 90:125–158. doi:10.1002/iroh.200410759
Hewson I, Brown JM, Burge CA, Couch CS, LaBarre BA, Mouchka ME, Naito M, Harvell CD (2011) Description of viral assemblages associated with the Gorgonia ventalina holobiont. Coral Reefs 31:487–491. doi:10.1007/s00338-011-0864-x
Hoegh-Guldberg O, Mumby PJ, Hooten AJ, Steneck RS, Greenfield P, Gomez E, Harvell CD, Sale PF, Edwards AJ, Caldeira K, Knowlton N, Eakin CM, Iglesias-Prieto R, Muthiga N, Bradbury RH, Dubi A, Hatziolos ME (2007) Coral reefs under rapid climate change and ocean acidification. Science 318:1737–1742. doi:10.1126/science.1152509
Holm JB, Heidelberg KB (2016) Microbiomes of Muricea californica and M. fruticosa: comparative analyses of two co-occurring eastern Pacific octocorals. Front Microbiol 7:917. doi:10.3389/fmicb.2016.00917
Horwitz R, Fine M (2014) High CO2 detrimentally affects tissue regeneration of Red Sea corals. Coral Reefs 33:819–829. doi:10.1007/s00338-014-1150-5
Hothorn T, Bretz F, Westfall P (2008) Simultaneous inference in general parametric models. Biom J 50:346–363. doi:10.1002/bimj.200810425
Hughes AD, Grottoli AG, Pease TK, Matsui Y (2010) Acquisition and assimilation of carbon in non-bleached and bleached corals. Mar Ecol Prog Ser 420:91–101. doi:10.3354/meps08866
Hunt LR, Smith SM, Downum KR, Mydlarz LD (2012) Microbial regulation in gorgonian corals. Mar Drugs 10:1225–1243. doi:10.3390/md10061225
Iglesias-Prieto R, Beltrán VH, LaJeunesse TC, Reyes-Bonilla H, Thomé PE (2004) Different algal symbionts explain the vertical distribution of dominant reef corals in the eastern Pacific. Proc R Soc Lond Ser B 271:1757–1763. doi:10.1098/rspb.2004.2757
Imbs AB, Yakovleva IM, Dautova TN, Bui LH, Jones P (2014) Diversity of fatty acid composition of symbiotic dinoflagellates in corals: evidence for the transfer of host PUFAs to the symbionts. Phytochemistry 101:76–82. doi:10.1016/j.phytochem.2014.02.012
Inoue S, Kayanne H, Yamamoto S, Kurihara H (2013) Spatial community shift from hard to soft corals in acidified water. Nat Clim Change 3:683–687. doi:10.1038/nclimate1855
Jackson JBC, Donovan MK, Cramer KL, Lam V (2014) Status and trends of Caribbean coral reefs: 1970–2012. Global Coral Reef Monitoring Network, IUCN, Gland
Jackson CR, Stone BWG, Tyler HL (2015) Emerging perspectives on the natural microbiome of fresh produce vegetables. Agriculture 5:170–187. doi:10.3390/agriculture5020170
Jensen P, Harvell CD, Wirtz K, Fenical W (1996) Antimicrobial activity of extracts of Caribbean gorgonian corals. Mar Biol 125:411–419. doi:10.1007/BF00346321
Jones RJ, Yellowlees D (1997) Regulation and control of intracellular algae (=zooxanthellae) in hard corals. Philos Trans R Soc Lond Ser B 352:457–468
Jordán-Dahlgren E (1989) Gorgonian community structure and reef zonation patterns on Yucatan coral reefs. Bull Mar Sci 45:678–696
Kanwisher JW, Wainwright SA (1967) Oxygen balance in some reef corals. Biol Bull 133:378–390. doi:10.2307/1539833
Kellogg CA, Lisle JT, Galkiewicz JP (2009) Culture-independent characterization of bacterial communities associated with the cold-water coral Lophelia pertusa in the northeastern Gulf of Mexico. Appl Environ Microbiol 75:2294–2303. doi:10.1128/AEM.02357-08
Kim K, Kim PD, Alker AP, Harvell CD (2000) Chemical resistance of gorgonian corals against fungal infections. Mar Biol 137:393–401. doi:10.1007/s002270000333
Kinzie RA III (1973) Zonation of West Indian gorgonians. Bull Mar Sci 23:93–155
Knowlton N, Rohwer F (2003) Multispecies microbial mutualisms on coral reefs: the host as a habitat. Am Nat 162:S51–S62. doi:10.1086/378684
Kopp C, Domart-Coulon I, Escrig S, Humbel BM, Hignette M, Meibom A (2015) Subcellular investigation of photosynthesis-driven carbon assimilation in the symbiotic reef coral Pocillopora damicornis. mBio 6:e02299-02214. doi:10.1128/mBio.02299-14
Kozich JJ, Westcott SL, Baxter NT, Highlander SK, Schloss PD (2013) Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl Environ Microbiol 79:5112–5120. doi:10.1128/AEM.01043-13
Kurahashi M, Yokota A (2007) Endozoicomonas elysicola gen. nov., sp. nov., a γ-proteobacterium isolated from the sea slug Elysia ornata. Syst Appl Microbiol 30:202–206. doi:10.1016/j.syapm.2006.07.003
Kuznetsova A, Brockhoff PB, Bojesen Christensen RH (2014) lmerTest: tests for random and fixed effects for linear mixed effect models (lmer objects of lme4 package). R package version 2.0
La Rivière M, Roumagnac M, Garrabou J, Bally M (2013) Transient shifts in bacterial communities associated with the temperate gorgonian Paramuricea clavata in the northwestern Mediterranean Sea. PLoS One 8:e57385. doi:10.1371/journal.pone.0057385.s001
La Rivière M, Garrabou J, Bally M (2015) Evidence for host specificity among dominant bacterial symbionts in temperate gorgonian corals. Coral Reefs 34:1087–1098. doi:10.1007/s00338-015-1334-7
Lang da Silveira F, Van’t Hof T (1977) Regeneration in the gorgonian Plexaura flexuosa (Cnidaria, Octocorallia). Bijdr Dierkd 47:98–108
Lasker HR (1985) Prey preferences and browsing pressure of the butterflyfish Chaetodon capistratus on Caribbean gorgonians. Mar Ecol Prog Ser 21:213–220
Lasker HR, Coffroth MA, Fitzgerald L (1988) Foraging patterns of Cyphoma gibbosum on octocorals: the roles of host choice and feeding preference. Biol Bull 174:254–266. doi:10.2307/1541952
Leal-Bautista RM, Hernández-Zárate G, Jaime MNA, Cuevas RG, Velázquez Oliman G (2011) Pathogens and pharmaceuticals pollutants as indicators of contamination at the northeastern aquifer of Quintana Roo. Trop Subtrop Agroecosyst 13:211–219
Lee OO, Yang J, Bougouffa S, Wang Y, Batang Z, Tian R, Al-Suwailem A, Qian P-Y (2012) Spatial and species variations in bacterial communities associated with corals from the Red Sea as revealed by pyrosequencing. Appl Environ Microbiol 78:7173–7184. doi:10.1128/AEM.01111-12
Lema KA, Willis BL, Bourne DG (2014) Amplicon pyrosequencing reveals spatial and temporal consistency in diazotroph assemblages of the Acropora millepora microbiome. Environ Microbiol 16:3345–3359. doi:10.1111/1462-2920.12366
Lenihan HS, Edmunds PJ (2010) Response of Pocillopora verrucosa to corallivory varies with environmental conditions. Mar Ecol Prog Ser 409:51–63. doi:10.3354/meps08595
Lenz EA, Bramanti L, Lasker HR, Edmunds PJ (2015) Long-term variation of octocoral populations in St. John, US Virgin Islands. Coral Reefs 34:1099–1109. doi:10.1007/s00338-015-1315-x
Lewis JB, Post EE (1982) Respiration and energetics in West Indian Gorgonacea (Anthozoa, Octocorallia). Comp Biochem Physiol A 71:457–459. doi:10.1016/0300-9629(82)90434-0
Mann WT, Beach-Letendre J, Mydlarz LD (2014) Interplay between proteases and protease inhibitors in the sea fan—Aspergillus pathosystem. Mar Biol 161:2213–2220. doi:10.1007/s00227-014-2499-2
McCauley EP, Haltli B, Correa H, Kerr RG (2016) Spatial and temporal investigation of the microbiome of the Caribbean octocoral Erythropodium caribaeorum. FEMS Microbiol Ecol 92:fiw147. doi:10.1093/femsec/fiw147
Meesters EH, Pauchli W, Bak RPM (1997) Predicting regeneration of physical damage on a reef-building coral by regeneration capacity and lesion shape. Mar Ecol Prog Ser 146:91–99. doi:10.3354/meps146091
Meyer JL, Paul VJ, Teplitski M (2014) Community shifts in the surface microbiomes of the coral Porites astreoides with unusual lesions. PLoS One 9:e100316. doi:10.1371/journal.pone.0100316.s002
Morrow KM, Moss AG, Chadwick NE, Liles MR (2012) Bacterial associates of two Caribbean coral species reveal species-specific distribution and geographic variability. Appl Environ Microbiol 78:6438–6449. doi:10.1128/AEM.01162-12
Morrow KM, Bourne DG, Humphrey C, Botte ES, Laffy P, Zaneveld J, Uthicke S, Fabricius KE, Webster NS (2015) Natural volcanic CO2 seeps reveal future trajectories for host-microbial associations in corals and sponges. ISME J 9:894–908. doi:10.1038/ismej.2014.188
Mouchka ME, Hewson I, Harvell CD (2010) Coral-associated bacterial assemblages: current knowledge and the potential for climate-driven impacts. Integr Comp Biol 50:662–674. doi:10.1093/icb/icq061
Murdock GR (1978a) Circulation and digestion of food in the gastrovascular system of gorgonian octocorals (Cnidaria; Anthozoa). Bull Mar Sci 28:363–370
Murdock GR (1978b) Digestion, assimilation, and transport of food in the gastrovascular cavity of a gorgonian octocoral (Cnidaria; Anthozoa). Bull Mar Sci 28:354–362
Mydlarz LD, Harvell CD (2007) Peroxidase activity and inducibility in the sea fan coral exposed to a fungal pathogen. Comp Biochem Physiol A 146:54–62. doi:10.1016/j.cbpa.2006.09.005
Mydlarz LD, Palmer CV (2011) The presence of multiple phenoloxidases in Caribbean reef-building corals. Comp Biochem Physiol A 159:372–378. doi:10.1016/j.cbpa.2011.03.029
Mydlarz LD, Holthouse SF, Peters EC, Harvell CD (2008) Cellular responses in sea fan corals: granular amoebocytes react to pathogen and climate stressors. PLoS One 3:e1811. doi:10.1371/journal.pone.0001811
Neave MJ, Apprill A, Ferrier-Pagès C, Voolstra CR (2016) Diversity and function of prevalent symbiotic marine bacteria in the genus Endozoicomonas. Appl Microbiol Biotechnol. doi:10.1007/s00253-016-7777-0
Olano C, Bigger C (2000) Phagocytic activities of the gorgonian coral Swiftia exserta. J Invert Pathol 76:176–184. doi:10.1006/jipa.2000.4974
Oren U, Benayahu Y, Loya Y (1997) Effect of lesion size and shape on regeneration of the Red Sea coral Favia favus. Mar Ecol Prog Ser 146:101–107. doi:10.3354/meps146101
Oren U, Benayahu Y, Lubinevsky H, Loya Y (2001) Colony integration during regeneration in the stony coral Favia favus. Ecology 82:802–813. doi:10.1890/0012-9658(2001)082[0802:cidrit]2.0.co;2
Palmer CV, McGinty ES, Cummings DJ, Smith SM, Bartels E, Mydlarz LD (2011a) Patterns of coral ecological immunology: variation in the responses of Caribbean corals to elevated temperature and a pathogen elicitor. J Exp Biol 214:4240–4249. doi:10.1242/jeb.061267
Palmer CV, Traylor-Knowles NG, Willis BL, Bythell JC (2011b) Corals use similar immune cells and wound-healing processes as those of higher organisms. PLoS One 6:e23992. doi:10.1371/journal.pone.0023992.g012
Penn K, Wu D, Eisen JA, Ward N (2006) Characterization of bacterial communities associated with deep-sea corals on Gulf of Alaska seamounts. Appl Environ Microbiol 72:1680–1683. doi:10.1128/AEM.72.2.1680-1683.2006
Perkins D, Ciereszko L (1973) The environmental toxicity of crassin acetate using Tetrahymina pyriformis as a model. Hydrobiologia 42:77–84. doi:10.1007/BF00014147
Petes LE, Harvell CD, Peters EC, Webb MAH, Mullen KM (2003) Pathogens compromise reproduction and induce melanization in Caribbean sea fans. Mar Ecol Prog Ser 264:167–171. doi:10.3354/meps264167
Pike RE, Haltli B, Kerr RG (2013) Description of Endozoicomonas euniceicola sp. nov. and Endozoicomonas gorgoniicola sp. nov., bacteria isolated from the octocorals Eunicea fusca and Plexaura sp., and an emended description of the genus Endozoicomonas. Int J Syst Evol Microbiol 63:4294–4302. doi:10.1099/ijs.0.051490-0
Porporato EMD, Lo Giudice A, Michaud L, De Domenico E, Spanò N (2013) Diversity and antibacterial activity of the bacterial communities associated with two Mediterranean sea pens, Pennatula phosphorea and Pteroeides spinosum (Anthozoa: Octocorallia). Microb Ecol 66:701–714. doi:10.1007/s00248-013-0260-x
Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, Peplies J, Glöckner FO (2013) The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res 41:D590–D596. doi:10.1093/nar/gks1219
R Core Team (2015) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Ramsby BD, Shirur KP, Iglesias-Prieto R, Goulet TL (2014) Symbiodinium photosynthesis in Caribbean octocorals. PLoS One 9:e106419. doi:10.1371/journal.pone.0106419
Razin S, Yogev D, Naot Y (1998) Molecular biology and pathogenicity of mycoplasmas. Microbiol Mol Biol Rev 62:1094–1156. doi:10.1007/b113360
Ribes M, Coma R, Gili J-M (1998) Heterotrophic feeding by gorgonian corals with symbiotic zooxanthella. Limnol Oceanogr 43:1170–1179. doi:10.4319/lo.1998.43.6.1170
Robertson V, Haltli B, McCauley E, Overy D, Kerr R (2016) Highly variable bacterial communities associated with the octocoral Antillogorgia elisabethae. Microorganisms 4:23. doi:10.3390/microorganisms4030023
Roder C, Arif C, Bayer T, Aranda M, Daniels C, Shibl A, Chavanich S, Voolstra CR (2014) Bacterial profiling of white plague disease in a comparative coral species framework. ISME J 8:31–39. doi:10.1038/ismej.2013.127
Roder C, Bayer T, Aranda M, Kruse M, Voolstra CR (2015) Microbiome structure of the fungid coral Ctenactis echinata aligns with environmental differences. Mol Ecol 24:3501–3511. doi:10.1111/mec.13251
Rodríguez A (1995) The natural product chemistry of West Indian gorgonian octocorals. Tetrahedron 51:4571–4618. doi:10.1016/0040-4020(95)00216-U
Ruesink J, Harvell CD (1990) Specialist predation on the Caribbean gorgonian Plexaurella spp. by Cyphoma signatum (Gastropoda). Mar Ecol Prog Ser 65:265–272
Ruiz-Diaz CP, Toledo-Hernández C, Sabat AM, Marcano M (2013) Immune response to a pathogen in corals. J Theor Biol 332:141–148. doi:10.1016/j.jtbi.2013.04.028
Rutherford ST, Bassler BL (2012) Bacterial quorum sensing: its role in virulence and possibilities for its control. Cold Spring Harb Perspect Med 2:a012427. doi:10.1101/cshperspect.a012427
Ruzicka R, Colella M, Porter J, Morrison J, Kidney J, Brinkhuis V, Lunz K, Macaulay K, Bartlett L, Meyers M, Colee J (2013) Temporal changes in benthic assemblages on Florida Keys reefs 11 years after the 1997/1998 El Niño. Mar Ecol Prog Ser 489:125–141. doi:10.3354/meps10427
Sabine AM, Smith TB, Williams DE, Brandt ME (2015) Environmental conditions influence tissue regeneration rates in scleractinian corals. Mar Pollut Bull 95:253–264. doi:10.1016/j.marpolbul.2015.04.006
Sánchez JA, McFadden CS, France SC, Lasker HR (2003) Molecular phylogenetic analyses of shallow-water Caribbean octocorals. Mar Biol 142:975–987. doi:10.1007/s00227-003-1018-7
Schloss PD, Westcott SL, Ryabin T, Hall JR, Hartmann M, Hollister EB, Lesniewski RA, Oakley BB, Parks DH, Robinson CJ, Sahl JW, Stres B, Thallinger GG, Van Horn DJ, Weber CF (2009) Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl Environ Microbiol 75:7537–7541. doi:10.1128/AEM.01541-09
Schloss PD, Gevers D, Westcott SL (2011) Reducing the effects of PCR amplification and sequencing artifacts on 16S rRNA-based studies. PLoS One 6:e27310. doi:10.1371/journal.pone.0027310.t004
Shirur KP, Ramsby BD, Iglesias-Prieto R, Goulet TL (2014) Biochemical composition of Caribbean gorgonians: implications for gorgonian—Symbiodinium symbiosis and ecology. J Exp Mar Biol Ecol 461:275–285. doi:10.1016/j.jembe.2014.08.016
Sunagawa S, DeSantis TZ, Piceno YM, Brodie EL, DeSalvo MK, Voolstra CR, Weil E, Andersen GL, Medina M (2009) Bacterial diversity and white plague disease-associated community changes in the Caribbean coral Montastraea faveolata. ISME J 3:512–521. doi:10.1038/ismej.2008.131
Sunagawa S, Woodley CM, Medina M (2010) Threatened corals provide underexplored microbial habitats. PLoS One 5:e9554. doi:10.1371/journal.pone.0009554.g001
Tanaka Y, Grottoli AG, Matsui Y, Suzuki A, Sakai K (2015) Partitioning of nitrogen sources to algal endosymbionts of corals with long-term 15N-labelling and a mixing model. Ecol Model 309–310:163–169. doi:10.1016/j.ecolmodel.2015.04.017
Thompson JR, Rivera HE, Closek CJ, Medina M (2015) Microbes in the coral holobiont: partners through evolution, development, and ecological interactions. Front Cell Infect Microbiol 4:176. doi:10.3389/fcimb.2014.00176
Thornhill DJ, Rotjan RD, Todd BD, Chilcoat GC, Iglesias-Prieto R, Kemp DW, LaJeunesse TC, Reynolds JM, Schmidt GW, Shannon T, Warner ME, Fitt WK (2012) A connection between colony biomass and death in Caribbean reef-building corals. PLoS One 6:e29535. doi:10.1371/journal.pone.0029535
Toledo-Hernández C, Ruiz-Diaz CP (2014) The immune responses of the coral. Invertebr Surviv J 11:319–328
Toledo-Hernández C, Zuluaga-Montero A, Bones-González A, Rodríguez JA, Sabat AM, Bayman P (2008) Fungi in healthy and diseased sea fans (Gorgonia ventalina): is Aspergillus sydowii always the pathogen? Coral Reefs 27:707–714. doi:10.1007/s00338-008-0387-2
Tracy AM, Koren O, Douglas N, Weil E, Harvell CD (2015) Persistent shifts in Caribbean coral microbiota are linked to the 2010 warm thermal anomaly. Environ Microbiol Rep 7:471–479. doi:10.1111/1758-2229.12274
van de Water JAJM, Ainsworth TD, Leggat W, Bourne DG, Willis BL, van Oppen MJH (2015) The coral immune response facilitates protection against microbes during tissue regeneration. Mol Ecol 24:3390–3404. doi:10.1111/mec.13257
van Woesik R (1998) Lesion healing on massive Porites spp. corals. Mar Ecol Prog Ser 164:213–220. doi:10.3354/meps164213
Vezzulli L, Pezzati E, Huete-Stauffer C, Pruzzo C, Cerrano C (2013) 16SrDNA pyrosequencing of the Mediterranean gorgonian Paramuricea clavata reveals a link among alterations in bacterial holobiont members, anthropogenic influence and disease outbreaks. PLoS One 8:e67745. doi:10.1371/journal.pone.0067745
Villamizar E, Díaz MC, Rützler K, de Nóbrega R (2014) Biodiversity, ecological structure, and change in the sponge community of different geomorphological zones of the barrier fore reef at Carrie Bow Cay, Belize. Mar Ecol 35:425–435. doi:10.1111/maec.12099
Voss GL (1956) Protective coloration and habitat of the shrimp Tozeuma carolinensis Kingsley, (Caridea: Hippolytidae). Bull Mar Sci 6:359–363
Vreeland HV, Lasker HR (1989) Selective feeding of the polychaete Hermodice carunculata Pallas on Caribbean gorgonians. J Exp Mar Biol Ecol 129:265–277. doi:10.1016/0022-0981(89)90108-1
Wahle CM (1980) Detection, pursuit, and overgrowth of tropical gorgonians by milleporid hydrocorals: perseus and medusa revisited. Science 209:689–691. doi:10.1126/science.209.4457.689
Wahle CM (1983) Regeneration of injuries among Jamaican gorgonians: the roles of colony physiology and environment. Biol Bull 165:778–790. doi:10.2307/1541478
Wahle CM (1985) Habitat-related patterns of injury and mortality among Jamaican gorgonians. Bull Mar Sci 37:905–927
Waters CM, Bassler BL (2005) Quorum sensing: cell-to-cell communication in bacteria. Annu Rev Cell Dev Biol 21:319–346. doi:10.1146/annurev.cellbio.21.012704.131001
Westfall PH (1997) Multiple testing of general contrasts using logical constraints and correlations. J Am Stat Assoc 92:299–306. doi:10.1080/01621459.1997.10473627
Wirshing HH, Messing CG, Douady CJ, Reed J, Stanhope MJ, Shivji MS (2005) Molecular evidence for multiple lineages in the gorgonian family Plexauridae (Anthozoa: Octocorallia). Mar Biol 147:497–508. doi:10.1007/s00227-005-1592-y
Yang C-S, Chen M-H, Arun AB, Chen CA, Wang J-T, Chen W-M (2010) Endozoicomonas montiporae sp. nov., isolated from the encrusting pore coral Montipora aequituberculata. Int J Syst Evol Microbiol 60:1158–1162. doi:10.1099/ijs.0.014357-0
Yue JC, Clayton MK (2005) A similarity measure based on species proportions. Commun Stat Theory Methods 34:2123–2131. doi:10.1080/STA-200066418
Ziegler M, Roik A, Porter A, Zubier K, Mudarris MS, Ormond R, Voolstra CR (2016) Coral microbial community dynamics in response to anthropogenic impacts near a major city in the central Red Sea. Mar Pollut Bull 105:629–640. doi:10.1016/j.marpolbul.2015.12.045
Acknowledgments
We thank the staff, students, and Roberto Iglesias-Prieto at the Unidad Académica de Sistemas Arrecifales, Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México for their assistance, and B. Ramsby, H. Pearson and M. McCauley for help in the field. We appreciate the help of M. Slattery, S. Ankisetty and W. Mann’s with the biochemical assays, J. Hoeksema and S. Brewer for their statistical guidance, and T. LaJeunesse with Symbiodinium naming. Work performed through the UMMC Molecular and Genomics Facility was partially supported by the National Institute of General Medical Sciences of the National Institutes of Health, including Mississippi INBRE (P20GM103476), Center for Psychiatric Neuroscience (CPN)-COBRE (P30GM103328) and Obesity, Cardiorenal and Metabolic Diseases-COBRE (P20GM104357). The FlowCAM particle analyzer used was funded by the National Science Foundation (NSF) MRI Grant No. 1126379. Funding for this work was provided by the Graduate Student Council and Department of Biology at the University of Mississippi to K.P.S. and the NSF under Grant No. IOS 0747205 and REU supplement to T.L.G. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the authors and do not necessarily reflect the views of the National Science Foundation.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflict of interest. Gorgonian corals were sampled with a collection permit.
Additional information
Responsible Editor: L. D. Mydlarz.
Reviewed by Undisclosed experts.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Shirur, K.P., Jackson, C.R. & Goulet, T.L. Lesion recovery and the bacterial microbiome in two Caribbean gorgonian corals. Mar Biol 163, 238 (2016). https://doi.org/10.1007/s00227-016-3008-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-016-3008-6