
Abstract
Pole walking (PW) has received attention not only as a whole-body exercise that can be adapted for elderly people with poor physical fitness but also as a possible intervention for the restoration of gait function in normal walking without the use of poles (i.e., conventional walking CW). However, the characteristics of PW, especially how and why PW training affects CW, remain unclear. The purpose of this study was to examine the characteristics of locomotor adaptation in PW from the perspective of kinematic variables. For this purpose, we compared the locomotor adaptation in PW and CW to that when walking on a split-belt treadmill in terms of spatial and temporal coordination. The result showed that adaptations to the split-belt treadmill in PW and CW were found only in interlimb parameters (step length and double support time ratios (fast/slow limb)), not in intralimb parameters (stride length and stance time ratios). In these interlimb parameters, the movement patterns acquired through split-belt locomotor adaptations (i.e., the aftereffects) were transferred between CW and PW regardless of whether the novel movement patterns were learned in CW or PW. The aftereffects of double support time and step length learned in CW were completely washed out by the subsequent execution in PW. On the other hand, the aftereffect of double support time learned in PW was not completely washed out by the subsequent execution in CW, whereas the aftereffect of step length learned in PW was completely washed out by the subsequent execution in CW. These results suggest that the neural mechanisms related to controlling interlimb parameters are shared between CW and PW, and it is possible that, in interlimb coordination, temporal coordination is preferentially stored in adaptation during PW.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Pole walking (PW) is a form of locomotion in which a person holds a pole in each hand and touches one of the poles to the ground simultaneously with the contralateral lower limb heel contact. PW has received attention as a whole-body exercise that can be adapted for elderly or disabled persons with poor physical fitness (Figard-Fabre et al. 2010; Parkatti et al. 2012; Monteiro et al. 2017; Fukusaki et al. 2018). Moreover, recent studies have proposed the usefulness of PW as a possible intervention for restoration of gait function in normal walking without the use of poles (i.e., conventional walking: CW) for stroke and Parkinson’s disease patients (Shin et al. 2015; Warlop et al. 2017; Gougeon et al. 2017; Obata et al. 2017). In the elderly and patients with Parkinson’s disease, it has been reported that locomotor training that uses PW is more effective to restore walking ability (i.e., walking speed) and standing balance than other trainings (Reuter et al. 2011; Figueiredo et al. 2013). However, the characteristics of PW, especially how and why PW training affects bipedal conventional walking (CW), remain unknown.
Using locomotor adaptation paradigms that investigate the transfer of adaptation between PW and CW would be an effective method to reveal the characteristics of PW training. In previous studies, using a split-belt adaptation paradigm, partial transfer between forward and backward walking (Choi and Bastian 2007), between walking and running (Ogawa et al. 2012, 2015), and among various walking (Vasudevan and Bastian 2010) and running speeds (Ogawa et al. 2018) has been reported. These results suggest that the neural control underlying human locomotion has specificity, and the aftereffect (i.e., the training effect) is not necessarily shared between two locomotor conditions, even if the same muscle groups were recruited. This is a potential issue for gait rehabilitation using PW as well as other walking aids. It is known that activity-dependent plasticity is ubiquitous in the central nervous systems (CNS) (Wolpaw 2007). If the neural controls underlying PW and bipedal CW are different, the repetitive use of walking poles may impede the reacquisition of CW through the modification of locomotor centers in the central nervous system.
To reveal the difference in neural control between PW and CW, we recently investigated the transfer between those two gait modes after split-belt walking adaptation (Obata et al. 2019). The results showed that the degrees of the aftereffects in CW and PW were not different, regardless of whether split-belt walking adaptation was induced in CW or PW, suggesting that the neural controls of PW and CW are not independent. However, it is still unknown which kinematic variables are related to these findings, because only the anterior component of the ground reaction force was adopted as the index of locomotor adaptation in the previous study. The difference between spatial and temporal gait parameters is especially important, both for understanding neural control and for applying CW as a gait rehabilitation. Spatial and temporal controls for locomotor adaptation have been suggested to take place independently, and temporal control would be more automatic and more heavily dependent on subcortical circuits (Malone and Bastian 2010; Malone et al. 2012). In children who have undergone a hemispherectomy, deficits of locomotor adaptation have been reported only in temporal gait parameters (Choi et al. 2009). Insight into the characteristics of PW from the perspective of kinematic variables would give us important information to develop gait rehabilitation targeting either temporal or spatial gait parameters.
In this study, we investigated split-belt walking adaptation in CW and PW and transfers between these two gait modes in terms of kinematic variables. From the aspect of spatial versus temporal gait parameters, we compared the difference in neural control mechanisms between CW and PW.
Methods
Subjects
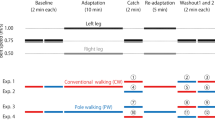
Eleven able-bodied male subjects (age: 26.2 ± 4.7 years old; height: 173.7 ± 4.2 cm; weight: 68.3 ± 6.6 kg; mean ± SD) with no known history of neurological disorders participated in this study. Each subject was tested in two experimental protocols (Fig. 1). The difference between these protocols was the gait mode used during adaptation periods (CW in Experiment 1 and PW in Experiment 2). The gait modes in washout periods and their combinations were also different between these protocols. The order of participation in these experiments was randomized across subjects. Two experiments were tested at intervals of at least 24 h. All participants gave informed written consent prior to participating in the study. The experimental procedures were conducted in accordance with the Declaration of Helsinki and were approved by the human ethics committee of the University of Tokyo, Japan.

Time course of the experimental protocol. Subjects were given a split-belt walking-adaptation task of either conventional walking (CW) (Exp. 1) or pole walking (PW) (Exp. 2). The adaptation and re-adaptation periods were 10 and 5 min, respectively, on an asymmetrically driven treadmill (the left belt was set to 1.00 m/s and the right to 0.50 m/s). The Baseline, Catch, and washout periods (Washout 1 and Washout 2) were each 2 min on a symmetrically driven treadmill (the left and right belts were set to 0.75 m/s)
Experimental protocols
In this study, subjects were asked to walk on a split-belt treadmill (Bertec, USA) that is composed of two separate belts. The treadmill belts can be controlled independently and driven either at the same velocity (i.e., tied belt) or at different velocities (i.e., split belt). During two baseline periods, the treadmill belts were operated at the same velocity, 0.75 m/s (Baseline; 2 min each, tied belt). Subsequently, during the adaptation period, the treadmill was operated at different velocities (Adaptation; 10 min, split belt): the left belt was set to 1.00 m/s (defined as fast limb) and the right to 0.50 m/s (defined as slow limb). After the adaptation period, the treadmill belts were returned to the tied-belt mode (Catch; 2 min) and then operated in split-belt mode (Re-adaptation; 5 min). Finally, the treadmill belts were again returned to the tied-belt mode (Washout 1 and Washout 2; 2 min). For safety, subjects were asked to stand to the outside of the belts before these changes and step onto the belts using their right foot first after the required belt velocity was reached. Subjects were also instructed to walk naturally and refrain from looking down at the treadmill belts to avoid receiving any visual information about velocity.
During PW, subjects were asked to place one of the Nordic poles vertically on either split belt with the contralateral lower limb heel contact at each stride. The length of the poles was adjusted so that the elbow joint angles were approximately 90° when subjects stood upright with the poles and kept them naturally in front of them. During CW, subjects were asked to swing their arms naturally.
Data recordings and analysis
An optical motion capture system (OptiTrack FLEX V100 R2, Natural Point, USA) composed of 12 infrared cameras was used to examine the step and stride length of both legs, and the arm swing. Reflective markers (5 mm in diameter) were placed bilaterally on the ankle (lateral malleolus), the back of the hand, and between the ulnar and radial styloids. Triaxial ground reaction forces (GRFs) were measured using two force plates mounted beneath each treadmill belt to examine foot contacts. Marker positions were recorded at a sampling rate of 100 Hz and synchronized with GRFs using OptiTrack software (Motive Version 2.0.2; Natural Point, USA). GRFs were recorded at a sampling rate of 1000 Hz (PowerLab; AD Instruments, Australia) and stored on a computer for later offline analysis.
Three-dimensional marker positions and force data were low-pass filtered at 6 Hz. Custom software written in MATLAB (R2019a; Mathworks, USA) was used for all analyses. Stride time was determined by detecting the moment of foot contact at which the vertical GRFs were more than 2.5 N and is defined as the period from one foot contact to the next for the same limb.
In this study, four kinematic variables, stride length, percent stance time, step length, and percent double support time, were calculated according to the previous study (Reisman et al. 2005). (1) Stride length is defined as the intralimb and spatial parameters calculated as the distance traveled by the ankle marker in the anterior–posterior direction from initial contact to the liftoff of one limb. (2) Stance time is defined as the intralimb and temporal parameters calculated as the duration of stance phase expressed as a percentage of the stride time. (3) Step length is defined as the interlimb and spatial parameters calculated as the anterior–posterior distance between the ankle marker of each leg at foot contact of the leading leg. (4) Double support time is defined as the interlimb and temporal parameters calculated as the duration that both feet were in contact with the split-treadmill, expressed as a percentage of the stride time for each leg. Double support time of the fast leg is the time from the slow leg foot contact to the fast leg liftoff, and vice versa for the double support time of the slow leg. These parameters were expressed as the ratio of the fast limb to the slow limb to assess asymmetry.
Arm swing ratio, which is the ratio of the left arm swing angle at the right heel contact and the right swing angle at the left heel contact, were also calculated in 8 of 11 subjects. Arm swing was defined as the angle between a vertical line and the vector from the acromion to the wrist on the sagittal plane: it was positive when the wrist was in front of the acromion. Each arm swing angle was calculated from the marker of the wrist and the acromion on both sides.
Averages of the last ten strides of the Baseline periods as well as the first (Initial) and last ten (Final) strides of the Adaptation, Catch, Washout 1, and Washout 2 periods were calculated for statistical comparisons.
Statistical analyses
Statistical analyses were performed for the number of steps taken during each period, the ratio of the fast limb to the slow limb in the four kinematic variables, and arm swing ratio using a commercially available software package (SPSS 21.0; SPSS, USA).
A paired t test was performed to compare the number of steps taking during Adaptation, Catch, Re-adaptation and Washout between CW and PW.
To compare the degree of adaptation and re-adaptation, a two-way analysis of variance (ANOVA) with repeated measures (factors: gait mode (CW or PW) and time points (Baseline, Initial, and Final strides)) was performed. In addition, to compare the degree of adaptation and re-adaptation among time points, a one-way ANOVA with repeated measures (factor: time points) was performed for each gait mode. When the one-way ANOVA showed a significant difference, Tukey’s post hoc comparisons were performed to test for differences among time points.
To compare the degree of transfer of the aftereffect (i.e., (1), (2), (4), (5) and in Fig. 1), a three-way ANOVA with repeated measures (factors: gait mode in the adaptation period (CW or PW), gait mode in the de-adaptation period (CW or PW), and time points (Initial or Final strides)) was performed. When the three-way ANOVA showed significant interactions, Tukey’s post hoc comparisons were performed.
To compare the degree of washout (i.e., (3), (6) and in Fig. 1), a two-way ANOVA with repeated measures (factors: gait mode (CW or PW) and time points (Initial or Final strides)) was performed. When the two-way ANOVA showed significant interactions, Tukey’s post hoc comparisons were performed.
The number steps were presented as the mean and standard deviation (mean ± SD). Data of kinematics variables were presented as the mean and standard error (mean ± SE). Statistical differences were accepted as significant when P < 0.05. For the results of ANOVA, the effect size is reported as η2.
Result
The number of steps taking during each period were not different between CW and PW. A paired t test showed no significant difference between these two gait modes during Adaptation (CW vs. PW, 971.3 ± 147.0 vs. 924.3 ± 120.9 steps, P = 0.27), Cath (198.0 ± 28.2 vs. 191.3 ± 20.8 steps, P = 0.30), Re-adaptation (494.7 ± 68.7 vs. 473.5 ± 59.4 steps, P = 0.21), Washout 1 (198.8 ± 22.7 vs. 190.0 ± 18.9 steps, P = 0.11) and Washout 2 (201.3 ± 24.3 vs. 197.2 ± 18.8 steps, P = 0.25).
Group means of the time series changes of the four kinematic variables are shown in Fig. 2. During the Baseline, where the belt conditions were tied, the ratio of each kinematic variable was approximately equal to 1 in exps. 1 and 2, which means the differences between the fast (left) and slow (right) limbs were very small. With exposure to split-belt conditions (i.e., Adaptation), the differences in step length and double support time were prominent for the first minute, whereas the differences did not diminish through the Adaptation periods in stride length and stance time in exps. 1 and 2. With a return to the tied-belt condition (i.e., Catch and Washout 1), the differences in step length and double support time between the fast and slow limbs were prominent for the first minute, whereas the differences in stride length and stance time were not clear in exps. 1 and 2. After the washout of a stored movement pattern (i.e., Washout 2), the difference in double support time between the fast and slow limbs still existed in Exp. 2. In the following section, the statistical results in each period are described.

Group mean of the time series changes in four kinematic parameters. Each plot was averaged over stride cycles in 10-s bins. Red circles and blue circles represent CW and PW, respectively
The degrees of adaptation were not different between CW and PW (i.e., exps. 1 and 2) (Fig. 3a). In stride length, stance time, step length, and double support ratios, a two-way repeated-measure ANOVA showed no significant interaction between the time points and gait mode, whereas the main effect of the time points was significant (stride length: F(2,20) = 481.735, P < 0.01, η2 = 0.98; stance time: F(2,20) = 36.218, P < 0.01, η2 = 0.78; step length: F(2,20) = 16.108, P < 0.01, η2 = 0.62; double support time: F(2,20) = 19.120, P < 0.01, η2 = 0.66). A one-way repeated-measure ANOVA showed the main effect of the time points in all four ratios of CW (stride length: F(2,20) = 248.190, P < 0.01, η2 = 0.96; stance time: F(2,20) = 23.115, P < 0.01, η2 = 0.70; step length: F(2,20) = 19.747, P < 0.01, η2 = 0.66; double support time: F(2,20) = 12.904, P < 0.01, η2 = 0.56) and PW (stride length: F(2,20) = 444.652, P < 0.01, η2 = 0.98; stance time: F(2,20) = 35.064, P < 0.01, η2 = 0.78; step length: F(2,20) = 10.970, P < 0.01, η2 = 0.52; double support time: F(2,20) = 13.870, P < 0.01, η2 = 0.58), and significant differences were found between the Baseline and Initial stride (P < 0.05). On the other hand, the difference between Initial and Final strides was found only in step length and double support time ratios in CW and PW (P < 0.05), indicating that locomotor adaptation occurred in these two parameters.

Comparison of adaptation (a) and re-adaptation (b) in an acquired movement pattern at different timepoints. Each bar is the average of the first ten strides (Initial) or last ten strides (Baseline and Final) during the Adaptation and Baseline periods. Red bars and blue bars represent CW and PW, respectively. Error bars represent the standard error of the mean. An asterisk indicates a significant difference from Baseline (*P < 0.05) or Initial (#P < 0.05)
The degrees of re-adaptation were also not different between CW and PW (Fig. 3b). In stride length, stance time, step length, and double support ratios, a two-way repeated-measure ANOVA showed no significant interaction between the time points and gait mode, whereas the main effect of the time points was significant (stride length: F(2,20) = 128.716, P < 0.01, η2 = 0.93; stance time: F(2,20) = 39.011, P < 0.01, η2 = 0.80; step length: F(2,20) = 35.192, P < 0.01, η2 = 0.78; double support time: F(2,20) = 7.777, P < 0.01, η2 = 0.44). A one-way repeated-measure ANOVA showed the main effect of the time points in all four ratios of CW (stride length: F(2,20) = 39.294, P < 0.01, η2 = 0.80; stance time: F(2,20) = 19.586, P < 0.01, η2 = 0.66; step length: F(2,20) = 24.436, P < 0.01, η2 = 0.71; double support time: F(2,20) = 6.527, P < 0.01, η2 = 0.40) and PW (stride length: F(2,20) = 267.628, P < 0.01, η2 = 0.96; stance time: F(2,20) = 61.276, P < 0.01, η2 = 0.86; step length: F(2,20) = 35.491, P < 0.01, η2 = 0.78; double support time: F(2,20) = 7.751, P < 0.01, η2 = 0.44), and significant differences were found between the Baseline and Initial stride (P < 0.05). The difference between Initial and Final strides was found only in step length and double support time ratios in CW and PW (P < 0.05).
The degrees of transfer to PW and CW were different among the four kinematic variables (Fig. 4). For the stride length and stance ratios, a three-way repeated-measure ANOVA showed no significant main effects or interactions. For the step length ratio, a three-way repeated-measure ANOVA showed the main effect of time points (F(1,10) = 61.720, P < 0.01, η2 = 0.86), interaction between gait modes in adaptation and de-adaptation periods (F(1,20) = 38.778, P < 0.01, η2 = 0.80), and interaction among the gait mode in the adaptation period, gait mode in the de-adaptation period, and time points (F(1,10) = 20.148, P < 0.01,, η2 = 0.67). For the double support ratio, a three-way repeated-measure ANOVA showed the main effect of time points (F(1,10) = 33.144, P < 0.01, η2 = 0.77), the main effect of gait mode in the adaptation period (F(1,10) = 6.967, P < 0.05, η2 = 0.41), interaction between the gait mode in the de-adaptation period and time points (F(1,20) = 9.652, P < 0.05, η2 = 0.49), and interaction between gait modes in the adaptation and de-adaptation periods (F(1,20) = 37.929, P < 0.01, η2 = 0.79). Post hoc testing of the step length and double support ratios revealed a significant difference between the Initial and Final strides in each combination of the gait modes in the adaptation and de-adaptation periods (P < 0.05). These ratios also revealed a significant difference between the gait mode in the adaptation period in CW and PW when transferred to PW at the Initial stride (P < 0.05). These results indicate that aftereffect of split-belt adaptation was observed only in step length and double support time, and the aftereffects in PW were stronger when a new movement pattern was acquired in PW than when it was done in CW.

Comparison of the transfer of the acquired movement pattern at different timepoints. Each bar is the average of the first ten strides (Initial) or last ten strides (Final). Red bars represent de-adaptation by CW after adaptation to CW (Catch in Exp. 1, filled red bars) and after adaptation to PW (Washout 1 in Exp. 2, open red bars). Blue bars represent de-adaptation by PW after adaptation to PW (Catch in Exp. 2, filled blue bars) and after adaptation to CW (Washout 1 in Exp. 1, open blue bars). Error bars represent the standard error of the mean. An asterisk indicates a significant difference between Initial and Final (*P < 0.05) or between the gait modes of Adaptation (#P < 0.05)
For double support time, the gait mode in which a new movement pattern was stored affected the washout of the aftereffect by another gait mode, whereas such a difference was not found in other kinematic variables (Fig. 5). A two-way repeated-measure ANOVA showed the main effects of the gait mode (F(1,10) = 5.120, P < 0.05, η2 = 0.34) and time points (F(1,10) = 37.985, P < 0.01, η2 = 0.79) as well as the interaction of gait modes and time points in the double support ratio (F(1,10) = 7.792, P < 0.05, η2 = 0.44). Post hoc testing revealed that the double support ratio washed out by CW was significantly larger than that by PW at the Initial stride (P < 0.05), and it was significantly lager at the Initial stride than that at the Final stride (P < 0.05). On the other hand, in the stride length and step length ratio, no statistical difference was found in a two-way repeated-measure ANOVA. In the stance time ratio, a two-way repeated-measure ANOVA showed the interaction of gait modes and time points (F(1,10) = 8.111, P < 0.05, η2 = 0.45), but no significant difference was found in post hoc testing. These results indicate that the aftereffect of PW that was related to double support movement could not be completely washed out by CW.

Comparison of the washout of the stored movement pattern at different timepoints. Red bars are the average of the first ten strides (Initial) or last ten strides (Final) during CW after washout by PW (Washout 2 in Exp. 1). Blue bars are the average of the first ten strides (Initial) or last ten strides (Final) during PW after washout by CW (Washout 2 in Exp. 2). Error bars represent the standard error of the mean. An asterisk indicates a significant difference between Initial and Final (*P < 0.05) or between the gait modes of Washout 2 (#P < 0.05)
For the arm swing ratio, the degrees of adaptation, transfer, and washout were not different between Exps. 1 and 2 (Fig. 6). Adaptation (Fig. 6a): A two-way repeated-measure ANOVA showed no significant main effect of the gait mode (F(1,14) = 0.088, P = 0.78, η2 = 0.01) and interaction between the time points and gait modes (F(2,14) = 0.042, P = 0.96, η2 = 0.01), whereas the main effect of the time points was significant (F(2,14) = 5.242, P < 0.05, η2 = 0.43). Transfer (Fig. 6b): A three-way repeated-measure ANOVA showed no significant main effects (time points: F(1,7) = 0.000, P = 0.99, η2 = 0.00; gait mode in the adaptation period: F(1,7) = 4.393, P = 0.07, η2 = 0.39; gait mode in the de-adaptation period: F(1,7) = 0.038, P = 0.85, η2 = 0.01) or interactions (time points × gait mode in the adaptation period × gait mode in the de-adaptation period, F(1,7) = 1.351, P = 0.28, η2 = 0.16; time × gait mode in the adaptation period, F(1,7) = 1.074, P = 0.34, η2 = 0.13; time points × gait mode in the de-adaptation period, F(1,7) = 3.736, P = 0.10, η2 = 0.35; gait mode in the adaptation period × gait mode in the de-adaptation, F(1,7) = 1.400, P = 0.28, η2 = 0.17). Washout (Fig. 6c): Two-way repeated-measure ANOVA showed no significant main effects (time points: F(1,7) = 1.779, P = 0.22, η2 = 0.20; gait mode of washout: F(1,7) = 0.000, P = 0.99, η2 = 0.00) or interactions (F(1,7) = 1.147, P = 0.32, η2 = 0.14).

Arm swing ratio in exps. 1 and 2. Comparison of adaptation in an acquired movement pattern (a), the transfer of the acquired movement pattern (b), and the washout of the stored movement pattern at different timepoints (c). Each bar is the average of the first ten strides (Initial) or last ten strides (Baseline and Final)
Discussion
The purpose of this study was to examine the characteristics of locomotor adaptation in PW from the perspective of kinematic variables. The results demonstrated that (1) no difference between PW and CW was found in split-belt walking adaptation, that is, the adaptation was observed in interlimb parameters (step length and double support ratios) but not in intralimb parameters; (2) the aftereffect size of interlimb temporal (step length ratio) and spatial parameters (double support ratio) was larger in PW than in CW when the novel movement was learned in PW, whereas no difference between PW and CW was found when it was learned in CW; and (3) the aftereffect of interlimb temporal parameter learned in PW was not completely washed out by the subsequent execution in CW, whereas the aftereffect of the interlimb spatial parameter learned in PW was completely washed out by the subsequent execution in CW. These results suggest unique characteristics of PW as compared to those of CW.
Spatial coordination is preferentially restored in split-belt adaptation
The aftereffect size of interlimb parameters in PW was larger when the novel movement pattern was learned in PW than when it was learned in CW, whereas the aftereffect size in CW was not different, regardless of whether the novel movement pattern was learned in PW or CW. These results suggest that storage of the aftereffect in PW is not the same as that in CW. In addition, the present results showed an asymmetrical washout: the aftereffect of the interlimb temporal parameter in CW was washed out by the subsequent execution in PW, whereas the pattern in PW was not completely washed out by the subsequent execution in CW. These results are consistent with those in our previous study (Obata et al. 2019). In that study, from an observation of the anterior component of the ground reaction force (braking force), we showed that the aftereffect in CW was washed out by the subsequent execution in PW, whereas the aftereffect in PW was not completely washed out by the subsequent execution in CW. Based on these results, we suggested that the neural mechanisms of PW and CW are not independent and that the neural process of CW was a subset of that for PW. The present results for the interlimb spatial and temporal parameters during de-adaptation periods (Catch and Washout 1) and the interlimb temporal parameter in Washout 2 support the notion that this neural structure between PW and CW exists.
Concerning the interlimb spatial parameter, the effect of the washout was symmetry: the aftereffect of the interlimb temporal parameter in CW was washed out by the subsequent execution in PW, and the pattern in PW was washed out by the subsequent execution in CW. These results suggest that, in PW, interlimb temporal coordination is stored more preferentially than interlimb spatial coordination. During PW, subjects are required to touch one pole to the ground at the same time as the contralateral heel contact. Therefore, subjects would be paying attention to their temporal gait coordination rather than their spatial gait coordination. This characteristic of PW may affect a significant aftereffect of interlimb temporal coordination. In a previous study, Malone and Bastian (2010) demonstrated that a conscious correction of spatial gait coordination, in response to instruction on how to step and intermittent visual feedback on stepping during adaptation, affected the adaptation speed for spatial but not temporal parameters. Likewise, in the present study, the use of a pole may have led to subjects’ awareness of temporal coordination, and adaptation to temporal coordination may be enhanced as compared to that for spatial coordination.
In interlimb coordination, previous studies have demonstrated the independence of the neural mechanisms for spatial and temporal coordination in focusing attention during adaptation (Malone and Bastian 2010), time course over development (Musselman et al. 2011; Vasudevan et al. 2011), and brain injury (Choi et al. 2009). Specifically, Choi et al. (2009) demonstrated that hemispherectomy affects interlimb temporal gait coordination, whereas the adaptation of interlimb spatial coordination was intact. They suggest that temporal modification requires a more complicated process involving the cerebrum, while spatial modification is a simpler adjustment. These differences in neural mechanisms would underlie the difference between spatial and temporal parameters during split-belt adaption in PW.
Adaptation process
In both PW and CW, intralimb parameters (stride length and stance time ratios) showed an almost immediate change during split-belt walking adaptation and no aftereffects, whereas interlimb parameters showed a gradual change and aftereffects. These results of difference among kinematic variables are consistent with the previous study of Reisman et al. (2005). In many studies of split-belt walking adaptation, predictive (feedforward) adjustment was found in interlimb gait parameters (e.g., step length), and reactive (feedback) adjustment was found in intralimb parameters (e.g., stride length) (Reisman et al. 2005, 2010; Morton and Bastian 2006; Vasudevan and Bastian 2010). The difference between these gait parameters during the split-belt adaptation has been suggested to reflect independent neural control of intra- versus interlimb coordination during human bipedal gait (Reisman et al. 2005, 2010). Pathological and neurological studies have shown that the cerebellum plays an essential role in feedforward adjustment (Morton and Bastian 2006; Jayaram et al. 2011, 2012), whereas the cerebellum and the cerebrum are not absolutely necessary for feedback adjustment (Morton and Bastian 2006; Choi et al. 2009).
During PW, a person touches one of the poles to the ground simultaneously with the contralateral lower limb heel contact. Thus, a new interlimb coordination between one upper limb and the contralateral lower limb would be formed during locomotor adaptation of PW. However, no kinematic variables were affected by this upper–lower interlimb coordination. In our previous study, we demonstrated that the ground reaction force of the anterior–posterior direction was not different between CW and PW (Obata et al. 2019). Therefore, one possible explanation of no difference between PW and CW during the adaptation period may be that the force generated by the upper–lower interlimb coordination is too small to induce kinematic changes as compared with the force exerted by intralimb and interlimb coordination in the lower limb. Another explanation is that the basic pattern of upper–lower interlimb coordination is kept between PW and CW, and a specific upper–lower interlimb coordination is not formed during locomotor adaptation of PW despite rather different kinetics. Supporting this possibility, no difference was found in the degree of adaptation of arm swing ratio between PW and CW.
Conclusion
In conclusion, the present results demonstrate that interlimb temporal coordination, as compared to interlimb spatial coordination, is preferentially stored through split-belt walking adaptation. The present result provides important information for gait rehabilitation: PW can modify a temporal walking pattern and would be beneficial for interventions targeting temporal walking deficits.
References
Choi JT, Bastian AJ (2007) Adaptation reveals independent control networks for human walking. Nat Neurosci 10(8):1055–1062
Choi JT, Vining EPG, Reisman DS, Bastian AJ (2009) Walking flexibility after hemispherectomy: split-belt treadmill adaptation and feedback control. Brain 132(3):722–733
Figard-Fabre H, Fabre N, Leonardi A, Schena F (2010) Physiological and perceptual responses to Nordic walking in obese middle-aged women in comparison with the normal walk. Eur J Appl Physiol 108(6):1141–1151
Figueiredo S, Finch L, Mai J, Ahmed S, Huang A, May NE (2013) Nordic walking for geriatric rehabilitation: a randomized pilot study. Disabil Rehabil 35(12):968–975
Fukusaki C, Leetawesup K, Kato N, Kadokura Y, Nakazawa K, Yano H, Ishii N (2018) Effects of defensive style Nordic walking intervention in patients with lumbar and lower-limb osteoarthritis. J Nov Physiother 8(2):383. https://doi.org/10.4172/2165-7025.1000383
Gougeon MA, Zhou L, Nantel J (2017) Nordic walking improves trunk stability and gait spatial-temporal characteristics in people with Parkinson disease. NeuroRehabilitation 41(1):205–210
Jayaram G, Galea JM, Bastian AJ, Celnik P (2011) Human locomotor adaptive learning is proportional to depression of cerebellar excitability. Cereb Cortex 21(8):1901–1909
Jayaram G, Tang B, Pallegadda R, Vasudevan EV, Celnik P, Bastian AJ (2012) Modulating locomotor adaptation with cerebellar stimulation. J Neurophysiol 107(11):2950–2957
Malone LA, Bastian AJ (2010) Thinking about walking: effects of conscious correction versus distraction on locomotor adaptation. J Neurophysiol 103(4):1954–1962
Malone LA, Bastian AJ, Torres-Oviedo G (2012) How does the motor system correct for errors in time and space during locomotor adaptation? J Neurophysiol 108(2):672–683
Monteiro EP, Franzoni LT, Cubillos DM, de Oliveira FA, Carvalho AR, Oliveira HB, Pantoja PD, Schuch FB, Rieder CR, Martinez FG, Peyré-Tartaruga LA (2017) Effects of Nordic walking training on functional parameters in Parkinson’s disease: a randomized controlled clinical trial. Scand J Med Sci Sports 27(3):351–358
Morton SM, Bastian AJ (2006) Cerebellar contributions to locomotor adaptations during split-belt treadmill walking. J Neurosci 26(36):9107–9116
Musselman KE, Patrick SK, Vasudevan EV, Bastian AJ, Yang JF (2011) Unique characteristics of motor adaptation during walking in young children. J Neurophysiol 105(5):2195–2203
Obata H, Ogawa T, Hoshino M, Fukusaki C, Masugi Y, Kobayashi H, Yano H, Nakazawa K (2017) Effects of aquatic pole walking on the reduction of spastic hypertonia in a patient with hemiplegia: a case study. Int J Phys Med Rehabil 5:401. https://doi.org/10.4172/2329-9096.1000401
Obata H, Ogawa T, Nakazawa K (2019) Unique controlling mechanisms underlying walking with two handheld poles in contrast to those of conventional walking as revealed by split-belt locomotor adaptation. Exp Brain Res 237(12):3175–3183
Ogawa T, Kawashima N, Ogata T, Nakazawa K (2012) Limited transfer of newly acquired movement patterns across walking and running in humans. PLoS ONE 7(9):e46349
Ogawa T, Kawashima N, Obata H, Kanosue K, Nakazawa K (2015) Mode-dependent control of human walking and running as revealed by split-belt locomotor adaptation. J Exp Biol 218(Pt 20):3192–3198
Ogawa T, Obata H, Yokoyama H, Kawashima N, Nakazawa K (2018) Velocity-dependent transfer of adaptation in human running as revealed by split-belt treadmill adaptation. Exp Brain Res 236(4):1019–1029
Parkatti T, Perttunen J, Wacker P (2012) Improvements in functional capacity from Nordic walking: a randomized-controlled trial among elderly people. J Aging Phys Act 20(1):93–105
Reisman DS, Block HJ, Bastian AJ (2005) Interlimb coordination during locomotion: what can be adapted and stored? J Neurophysiol 94:2403–2415
Reisman DS, Bastian AJ, Morton SM (2010) Neurophysiologic and rehabilitation insights from the split-belt and other locomotor adaptation paradigms. Phys Ther 90(2):187–195
Reuter I, Mehnert S, Leone P, Kaps M, Oechsner M, Engelhardt M (2011) Effects of a flexibility and relaxation programme, walking, and Nordic walking on Parkinson’s disease. J Aging Res 2011:232473. https://doi.org/10.4061/2011/232473
Shin JH, Kim CB, Choi JD (2015) Effects of trunk rotation induced treadmill gait training on gait of stroke patients: a randomized controlled trial. J Phys Ther Sci 27(4):1215–1217
Vasudevan EV, Bastian AJ (2010) Split-belt treadmill adaptation shows different functional networks for fast and slow human walking. J Neurophysiol 103(1):183–191
Vasudevan EV, Torres-Oviedo G, Morton SM, Yang JF, Bastian AJ (2011) Younger is not always better: development of locomotor adaptation from childhood to adulthood. J Neurosci 31(8):3055–3065
Warlop T, Detrembleur C, Buxes Lopez M, Stoquart G, Lejeune T, Jeanjean A (2017) Does Nordic walking restore the temporal organization of gait variability in Parkinson’s disease? J Neuroeng Rehabil 14(1):17. https://doi.org/10.1186/s12984-017-0226-1
Wolpaw JR (2007) Spinal cord plasticity in acquisition and maintenance of motor skills. Acta Physiol 189(2):155–169
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Francesco Lacquaniti.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Obata, H., Ogawa, T., Yokoyama, H. et al. Spatiotemporal characteristics of locomotor adaptation of walking with two handheld poles. Exp Brain Res 238, 2973–2982 (2020). https://doi.org/10.1007/s00221-020-05954-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-020-05954-0