
Abstract
Rolling walkers are common walking aids for individuals with poor physical fitness or balance impairments. There is no doubt that rolling walkers are useful in assisting locomotion. On the other hand, it is arguable that walking with rolling walkers (WW) is effective for maintaining or restoring the nervous systems that are recruited during conventional walking (CW). This is because the differences and similarities of the neural control of these locomotion forms remain unknown. The purpose of the present study was to compare the neural control of WW and CW from the perspective of a split-belt adaptation paradigm and reveal how the adaptations that take place in WW and CW would affect each other. The anterior component of the ground reaction (braking) forces was measured during and after walking on a split-belt treadmill by 10 healthy subjects, and differences in the peak braking forces between the left and right sides were calculated as the index of the split-belt adaptation (the degree of asymmetry). The results demonstrated that (1) WW enabled subjects to respond to the split-belt condition immediately after its start as compared to CW; (2) the asymmetry movement pattern acquired by the split-belt adaptation in one gait mode (i.e., CW or WW) was less transferable to the other gait mode; (3) the asymmetry movement pattern acquired by the split-belt adaptation in CW was not completely washed out by subsequent execution in WW and vice versa. The results suggest unique control of WW and the specificity of neural control between WW and CW; use of the walkers is not necessarily appropriate as training for CW from the perspective of neural control.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Walking aids are widely used as assistive devices for individuals with poor physical fitness or balance impairments (Bateni and Maki 2005; Bertrand et al. 2017). There are many types of walking aids, such as canes, crutches, walkers, rolling walkers (four-wheeled walkers), and their variations (Bradley and Hernandez 2011). These walking aids have a variety of uses, such as in facilities that provide medical facilities (i.e., hospitals and rehabilitation centers) and care for the aged, as well as assisting the frail elderly in their activities of daily living. There are two purposes for using walking aids. One is as an alternate means of ensuring mobility instead of conventional walking (CW), and the other is a training assistive device for the re-acquisition of CW.
There is no doubt that walking aids are useful in assisting locomotion by distributing the user’s weight on walking aids and reducing loads on the lower limbs. On the other hand, it is arguable that walking with walking aids is effective for maintaining and restoring CW. This is due to the specificity of the neural control underlying human locomotion (Choi and Bastian, 2007; Vasudevan and Bastian 2010) and plasticity of the central nervous system (CNS) (Wolpaw 2007). In the previous studies, the specificity of the neural control has been reported between forward and backward walking (Choi and Bastian 2007), between walking and running (Ogawa et al. 2012, 2015), and between pulling forces during walking (Ogawa et al. 2023). These results suggest that, even if the same muscles are recruited in similar walling modes, it is possible that the neural controls in those walking modes are not the same. This is critical when walking aids are used for CW training. If the neural controls between CW and walking with walking aids are different and the use of such walking aids is extended for a prolonged period of time, the neural circuits related to locomotion could be modified to optimize walking with walking aids and may interfere with CW because of the plasticity in the CNS. These risks would become apparent when walking aids are used in daily life, whereas it would not matter as long as their use is limited to a short period (such as a rehabilitation period after some injuries). Therefore, it is important to know whether and to what extent the neural control of walking with walking aids is shared with that of CW, especially in walking aids used by the elderly on a daily basis.
To reveal characteristics of the neural control of walking with walking aids, a split-belt adaptation paradigm would be an effective method (Reisman et al. 2005). This paradigm promotes adaptation to treadmill walking in which the left and right belt speeds are different in one walking mode and investigates the transfer of the adaptation to the other walking mode. The aforementioned specificity of locomotion has been investigated using this paradigm (Choi and Bastian 2007, 2015, 2023; Ogawa et al. 2012). Recently, we compared the neural control of CW and pole walking (PW) using this paradigm; we suggested that the neural control of PW and CW are not independent, and the neural control of PW is built upon that of CW (Obata et al. 2019). As in this case, if the neural control is almost shared, a walking aid would be useful for maintaining and restoring CW. If it is not so, a walking aid would not be suitable for those purposes.
In the present study, we focused on rolling walkers because these walking aids are very common in senior-living communities (Liu 2009; Geravand et al. 2017; Costamagna et al. 2019). The purpose of the present study was to compare the neural control of CW and walking with a rolling walker (WW) from the perspective of a split-belt adaptation paradigm and to reveal how locomotor adaptation that takes place in WW and CW would affect each other. To reveal similarities and differences in the neural controls underlying these two walking modes, it was also tested whether adaptation in one walking mode was washed out by the subsequent execution of the other. Knowledge from this study provides evidence-based data on the use of walking aids and gives clues for developing a new intervention and device for maintaining walking function in the frail elderly who use walking aids in daily life.
Methods
Subjects
Ten able-bodied male subjects (age: 24.8 ± 1.5 years old; height: 173.7 ± 4.6 cm; weight: 69.7 ± 8.3 kg; mean ± SD) with no known history of neurological disorders participated in this study. Each subject was tested in two experimental protocols. The order of participation in these experiments was randomized across subjects. All subjects gave written informed consent prior to participating in the study. The experimental procedures were conducted in accordance with the Declaration of Helsinki and were approved by the human ethics committee of the University of Tokyo, Japan (ethics number: 701–4).
Experimental protocols
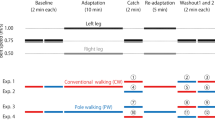
Two experiments were adapted to investigate the relationships between CW and walking with a rolling walker (Nissin, Japan) in locomotor adaptation and de-adaptation using a split-belt treadmill (Bertec, Columbus, OH, USA). The gait modes (CW and WW) in each period and their combination were different between these experiments (Fig. 1). The experiments were tested at intervals of at least one week. During CW, subjects were instructed to walk and swing their arms naturally. During WW, subjects were asked to put their weight onto the forearm support of the walker naturally and grip the handlebars. The walker was fixed on the split-belt treadmill by strings from external frames of the outside of the treadmill. The height between the forearm support of the walker and the surface of treadmill was 100.0 cm.

Time course of the experimental protocols. Subjects were given a split-belt adaptation task of either conventional walking (CW) (Exp. 1) or walking with a rolling walker (WW) (Exp. 2). The adaptation (Adaptation) and re-adaptation periods (Re-adaptation) were 10 and 5 min, respectively, on an asymmetrically driven treadmill (the left belt was set to 1.00 m/s and the right to 0.50 m/s). The baseline (Baseline), catch (Catch), and washout periods (Washout 1 and Washout 2) were 2 min each on a symmetrically driven treadmill (at 0.75 m/s)
Subjects were asked to walk on a split-belt treadmill composed of two separate belts. The treadmill belts can be controlled independently and driven either at the same velocity (i.e., tied belt; 0.75 m/s) or at different velocities (i.e., split belt; left: 1.00 m/s; right: 0.50 m/s). During two baseline periods, the treadmill belts were operated at the same velocity (Baseline 1 and Baseline 2; 2 min each, tied belt). Subsequently, during the adaptation period, the treadmill was operated at different velocities (Adaptation; 10 min, split belt). After the adaptation period, the treadmill belts were returned to the same velocity (Catch; 2 min, tied belt) and then operated at different velocities (Re-adaptation; 5 min, split belt). Finally, the treadmill belts were again returned to the same velocity (Washout 1 and Washout 2; 2 min each, tied belt). For safety, subjects were asked to stand outside the belts before these speed changes and step onto the belts using their right foot first after the required belt velocity was reached.
Data recordings and analysis
Triaxial ground reaction forces (GRFs) were measured using two force plates mounted beneath each treadmill belt. The force data were digitized at a sampling frequency of 1 kHz using an analog-to-digital converter and low-pass filtered at 8 Hz (PowerLab, AD Instruments, Sydney, Australia). Stride cycles were determined by detecting the moment of foot contact at which the vertical ground reaction forces were more than half of a subject’s weight. Given that gait speed is quotient of length (spatial) and time (temporal factors), subject could potentially employ complex strategies to adapt the split-belts: walking with spatially symmetrical with temporally asymmetrical movement patters, temporally symmetrical with spatially asymmetrical movement patterns, or changing the both parameter (Ogawa et al. 2012). Therefore, in this study, peak absolute values of the anterior (braking) component of the GRF were employed as the overall indicator to estimate symmetry and asymmetry induced by the split-belt adaptation and calculated for each stride cycle in accordance with our previous study (Ogawa et al. 2012, 2023; Obata et al. 2019). Next, differences in the peak forces between the left and right strides were calculated as the index of the split-belt adaptation (the right peak braking force subtracted from the left force; the degree of asymmetry). The data for the first stride cycle of each period were excluded from later analysis since gaits were remarkably perturbed when subjects started walking on the moving belts.
To estimate the unloading of the subject’s body weight due to the use of the walker, the following signal processing was performed. First, the vertical component of the GRFs of the final 10 steps during baseline periods was normalized to gait cycle [%] based on the left and right heel contact, and the additive mean waveforms were calculated for each gait mode and each side of the left and right legs. Then, the mean forces from 70 to 90% of gait cycle, which corresponded to the center of the swing phase, were calculated and averaged in Exp.1 and Exp.2. The left and right mean forces were averaged because the mean forces on the left and right sides were not statistically different in each gait mode, and the mean force in WW was subtracted from that in CW. The calculated value reflects the estimated unloading on one side, and the doubled value is the total unloaded body weight with use of the walker.
Statistical analysis
Statistical analyses were performed using a commercially available software package (SPSS 21.0, SPSS, Chicago, IL, USA). Parametric tests were used because the Shapiro–Wilk tests showed that calculated values in each period of two experiments were normally distributed.
Two-way analysis of variance (ANOVA) with repeated measures was used to test for statistical difference in the peak braking forces in the anterior component of GRFs and the mean forces in the vertical component of GRFs during baseline periods (factors: gait mode (CW or WW) and side (left or right leg)). A paired Student’s t test was performed to compare the difference in the degree of asymmetry between CW and WW.
To compare the degree of asymmetry during adaptation (i.e., Adaptation) or washout (i.e., Washout 2), a two-way ANOVA with repeated measures (factors: gait mode (CW or WW) and time point (average of the first or last 10 strides)) was performed. The number of first and last strides was referred to in our previous study (Obata et al. 2019).
To compare the degree of asymmetry during de-adaptation (i.e., Catch and Washout 1), a three-way ANOVA with repeated measures (factors: gait mode of adaptation (CW or WW), gait mode of de-adaptation (CW or WW), and time point (average of the first or last 10 strides)) was performed. When the three-way ANOVA gave significant interactions, a two-way ANOVA with repeated measures (factors: gait mode of adaptation (CW or WW) and time point (average of the first or last 10 strides)) was performed in each gait mode of de-adaptation. A one-way ANOVA was also performed to compare the degree of transfer over the first 10 strides in each condition.
When the two- or one-way ANOVA revealed significant results, Tukey’s post-hoc comparisons were performed to identify significant differences between variables. Data are presented as the mean and standard error (mean ± SE). Statistical differences were accepted as significant when P < 0.05. For the results of ANOVA, the effect size was reported as η2.
Results
Baseline
To confirm how using the walker affected the anteroposterior and vertical components of GRFs, representative values were calculated in these components. Figure 2(A) shows comparisons of the typical wave forms of the anteroposterior component of GRFs, the average of braking forces, and the average of the degree of asymmetry between CW and WW. As shown in the typical waveforms, the amplitude of the anteroposterior component of GRFs was smaller in WW than that in CW. The averages of the peak braking force over the last 10 strides were compared between CW and WW. A two-way repeated-measure ANOVA showed only the main effect of gait mode (F(1,9) = 141.426, P < 0.05, η2 = 0.94). Post-hoc testing revealed significant difference between the CW and WW in both sides (P < 0.05). Because the main effect of side was not significant (F(1,9) = 0.631, P = 0.45, η2 = 0.07), the left and right braking forces were averaged, and the averages of the degree of asymmetry were compared between CW and WW. A paired Student’s t-test showed no statistical difference between these walking modes. These results indicate that use of the walker reduced the peak braking forces of left and right sides but did not affect their asymmetry.

Comparison of the anteroposterior (A) and vertical GRFs (B) between CW and WW during baseline periods. A Representative waveforms of the anteroposterior GRFs (average of the left and right sides, and Exps. 1 and 2) (left panel), average of the peak braking force (average of Exps. 1 and 2) on the left and right sides (middle panel), and average of the degree of asymmetry between CW and WW (i.e., peak braking force differences) (right panel). B Representative waveforms of the vertical GRFs (average of the left and right sides, and Exps. 1 and 2) (left panel), average of the mean force from 70 to 90% gait cycle (i.e., stance phase) (average of Exps. 1 and 2) on the left and right sides (middle panel), and average of estimated unloading on one side with use of a walker (right panel). Red lines and bars represent conventional walking (CW), and blue lines and bars represent walking with a walker (WW). Error bars represent the standard error of the mean. An asterisk indicates a significant difference between CW and WW (*P < 0.05)
Figure 2(B) shows comparisons of the typical wave forms of the vertical component of GRFs, average of the mean force from 70 to 90% gait cycle, and average of estimated unloading by use of the walker. The waveforms show that the stance phase was approximately from 0 to 60% of gait cycle, and the swing phase was approximately from 60 to 100%. In the waveforms, during the stance phase, it was shown that the vertical GRF in WW was smaller than that in CW, indicating the use of the walker reduced lower limbs loading during this phase. During the swing phase, the vertical GRF in WW was larger than that in CW. This was because the load on the walker caused by unloading subject’s weight was added on the vertical GRF during this phase. It should be noted that the vertical GRF found in CW during stance phase was not zero. This was due to the walker always being on the split-belt and force plates. To quantify the mean force during the swing phase, averages of the mean force from 70 to 90% of gait cycle were calculated. A two-way repeated-measure ANOVA showed only the main effect of gait mode (F(1,9) = 311.147, P < 0.05, η2 = 0.97). Post-hoc testing revealed significant difference between the CW and WW in both sides (P < 0.05, respectively). Because the main effect of side was not significant (F(1,9) = 2.594, P = 0.14, η2 = 0.22), the left and right mean forces were averaged, and the values in WW were subtracted from those in CW. The calculated value reflects estimated unloading in one side (17.5 ± 1.0% of body weight, right panel), and the doubled value is the estimated total unloaded body weight by use of the walker.
Adaptation
During Adaptation periods, the peak braking forces of both sides and the degrees of asymmetry reached steady-state values over time in CW (Fig. 3(A), Adaptation in Exp. 1, upper and lower panels), whereas adaptation curve was not clear in WW (Fig. 3(B), Adaptation in Exp. 2, upper and lower panels). In addition, it should be noted that when subjects adapted the split-belt condition in WW, the initial values of the degrees of asymmetry were closer to the final values than those when subjects adapted the split-belt condition in CW. In Fig. 4, the averages of the degrees of asymmetry at the initial (the first 10 strides) and final time points (the last 10 strides) during Adaptation periods were compared between CW and WW. A two-way repeated-measure ANOVA showed significant interaction (gait mode × time point, F(1,9) = 9.377, P < 0.05, η2 = 0.51). Post-hoc testing revealed significant differences between the initial and final time points in both gait modes (P < 0.05). It also revealed that the degree of asymmetry at the initial time point in WW was significantly larger than that in CW (P < 0.05).

Typical examples of stride-to-stride profiles of the peak braking force (upper panels) and the degree of asymmetry (lower panels) in Exp. 1 (A) and Exp. 2 (B) for a single subject. Red circles and blue circles represent the peak braking force in CW and WW, respectively. Filled circles and open circles represent the fast (left) and slow (right) sides, respectively. Red triangles and blue triangles represent the differences in the peak braking force between the fast and slow sides (i.e., degree of asymmetry) in CW and WW, respectively

Comparison of the degree of asymmetry between CW and WW at the initial and final time points during adaptation periods. Each bar is the average of the first ten strides (Initial) or last ten strides (Final) during adaptation periods. Red bars and blue bars represent CW (①) and WW (⑤), respectively. Error bars represent the standard error of the mean. Asterisks indicate significant differences between Initial and Final or between CW and WW (*P < 0.05)
De-adaptation
The degree of transfer to CW or WW was complicated depending on the combination between the gait modes of adaptation and de-adaptation (Fig. 3(A) and (B), Catch and Washout 1, lower panels). The mean data of the degrees of asymmetry at the first and last 10 strides were compared in the Catch and Washout 1 periods (Fig. 5). A three-way repeated-measure ANOVA showed significant interaction (gait mode of adaptation × gait mode of de-adaptation × time point, F(1,72) = 38.280, P < 0.05, η2 = 0.81). Therefore, to simplify the interpretation of the results, the effects of the gait mode of adaptation and time points were compared in the de-adaptation by CW (Fig. 5 (A)) and WW (Fig. 5 (B)). In the former comparison, a two-way repeated-measure ANOVA showed significant interaction (gait mode of adaptation × time point, F(1,9) = 22.283, P < 0.05, η2 = 0.81). Post-hoc testing revealed a significant difference between the initial and final time points only after adaptation to CW (P < 0.05). In the latter comparison, a two-way repeated-measure ANOVA showed the main effects of the gait mode of adaptation (F(1,9) = 23.684, P < 0.05, η2 = 0.73) and time point (F(1,9) = 16.233, P < 0.05, η2 = 0.64) but did not show significant interaction (F(1,9) = 2.539, P = 0.15, η2 = 0.22). Post-hoc testing revealed significant difference between the initial and final time points both after learning WW and CW (P < 0.05). At the initial time points, the effects of combining gait modes of adaptation and de-adaptation were also compared (Fig. 5 (C)). A one-way repeated-measure ANOVA showed the main effect (F(3,27) = 49.348, P < 0.05, η2 = 0.85). Post-hoc testing revealed a significant difference between conditions ② (CW → CW) and ⑥ (WW → WW), ② and ⑦ (WW → CW), ② and ③ (CW → WW), ③ and ⑥, and ⑥ and ⑦ (P < 0.05).

Comparison of the degree of asymmetry during Catch and Washout 1 periods by CW (A) and WW (B) at the Initial and Final time points, and comparison of the degree of asymmetry at the Initial time point in each condition (combination of CW and WW in Adaptation and De-adaptation) (C). Each bar is the average of the first ten strides (Initial) or last ten strides (Final) during Catch or Washout 1 periods. Red bars represent de-adaptation by CW after adaptation to CW (Catch in Exp. 1, filled red bars, ②) and after adaptation to WW (Washout 1 in Exp. 2, open red bars, ⑦). Blue bars represent de-adaptation by WW after adaptation to WW (Catch in Exp. 2, filled blue bars, ⑥) and after adaptation to CW (Washout 1 in Exp. 1, open blue bars, ③). Error bars represent the standard error of the mean. An asterisk indicates a significant difference between Initial and Final or among conditions at Initial (*P < 0.05)
Washout
A new movement pattern that was stored by one gait mode could not be completely washed out by another gait mode (Fig. 3(A) and (B), Washout 2, lower panels). Figure 6 compares the mean data of the degrees of asymmetry between the initial and final time points during Washout 2. A two-way repeated-measure ANOVA showed significant interaction (gait mode × time point, F(1,9) = 26.774, P < 0.05, η2 = 0.75). Post-hoc testing revealed that the degree of asymmetry during the initial time points was significantly larger than that during the final time points both in CW (adapted in CW and washed out by WW, CW → WW → CW) and WW (adapted in WW and washed out by CW, WW → CW → WW) (P < 0.05). It also revealed that the degree of asymmetry during the initial time points was significantly larger in CW (CW → WW → CW) than that in WW (WW → CW → WW) (P < 0.05).

Comparison of the degree of asymmetry during Washout 2 at the initial and final time points. Red bars are the average of the first ten strides (Initial) or last ten strides (Final) during CW after washout by WW (Washout 2 in Exp. 1, ④). Blue bars are the average of the first ten strides (Initial) or last ten strides (Final) during WW after washout by CW (Washout 2 in Exp. 2, ⑧). An asterisk indicates a significant difference between Initial and Final or between CW and WW (*P < 0.05)
Discussion
The purpose of the present study was to compare the neural control of WW and CW from the perspective of a split-belt adaptation paradigm and reveal how locomotor adaptation that took place in WW and CW would affect each other. The present results demonstrate that (1) WW enabled subjects to respond to the split-belt condition immediately after its start as compared to CW; (2) movement patterns acquired by split-belt adaptation were less transferable between WW and CW; (3) the movement pattern acquired by split-belt adaptation in the CW mode was not completely washed out by subsequent execution in the WW mode and vice versa. The results suggest that there is a unique controlling mechanism underlying WW and specificity of neural control between WW and CW.
Locomotor adaptation
In adaptation periods, the degree of asymmetry of the first 10 strides was closer to that of the last 10 strides in WW than in CW. This result could be interpreted as showing that, in WW, subjects could respond or adapt to the split-belt condition immediately after the adaptation period began, as compared to that in CW. The changes in the anterior braking forces during the split-belt adaptation have been suggested to reflect the reactive feedback and predictive feedforward control strategies (Ogawa et al. 2014). The reactive feedback control plays a primary role during the initial part of the adaptation, and the predictive feedforward control takes time to play a role. The cerebellum’s involvement has been suggested to update feedforward control in the previous reports of pathophysiological (Morton and Bastian 2006) and neurophysiological studies (Jayaram et al. 2011, 2012). Therefore, our result suggests that the feedback control was effective and/or the feedforward control was updated early to the split condition in WW.
Human locomotion is controlled from two ways—walking balance and leg movements. In response to these controls, neural resources must be allocated to the CNS. As compared to CW, the use of a walker increases the base of support, and walking stability (balance) is greatly improved (Bateni and Maki 2005). Therefore, the use of a walker could save neural resources required for balance control during walking and focus them on leg movements. In addition, in WW, arm movements are restricted because subjects hold handlebars attached to the walker. The neural resources to control arm movements, including coordination with lower movements, also would be saved in WW. In a few previous studies, dual tasks, which divide neural resources between two tasks, have been reported to impair or delay motor adaptation in a reaching task (Taylor and Thoroughman 2007) and a split-belt walking adaptation (Malone and Bastian 2010). We also demonstrated that during PW, which requires strict upper and lower limb coordination, split-belt adaptation was delayed as compared to that during CW (Obata et al. 2019). Therefore, the features of stable walking and fixed arm movements in WW may affect the feedback and feedforward control of human locomotion through the allocation of the neural resources in the CNS and cause the difference in the degree of asymmetry between CW and WW at the initial points of adaptation.
Neural control
Limited transfer or washout of new movement patterns, which is acquired by split-belt adaptation, has been accepted as indirect evidence of independent control in human studies (Choi and Bastian 2007; Vasudevan and Bastian 2010; Ogawa et al. 2012, 2015). The present results show that the movement pattern acquired in CW was not completely washed out by subsequent execution in WW (Exp. 1, Washout 2), and the movement pattern acquired in WW was not completely washed out by subsequent execution in CW (Exp. 2, Washout 2). At the initial points of de-adaptation (i.e., Exps. 1 and 2, Catch and Washout 1), the degree of asymmetry was significantly smaller when the walking modes were different between the adaptation and de-adaptation periods than when those were the same. These results suggest that the neural control of WW and CW are independent.
When the walking modes were the same for adaptation and de-adaptation, the degrees of transfer (i.e., the degrees of asymmetry at the initial time points during de-adaptation) to WW after adaptation in WW were smaller than those to CW after adaptation in CW. These results imply that the storage, which retains movement patterns acquired by the split-belt adaptation, is small in the WW mode as compared to that in the CW mode. As described in the previous section, Locomotor Adaptation, in human locomotion, the CNS must control walking balance and leg movements. The storage that retains an acquired movement pattern would be a component of the feedforward system that controls these two control variables. In WW, the walker compensated for most of the walking balance, and the feedforward system learned only part of the leg movements for locomotion. On the other hand, in CW, the feedforward system learned both the balance and leg movements for locomotion. This may explain why the storage in WW is small.
In the previous study, handrail holding during split-belt adaptation have reduced the asymmetry of left and right step lengths during the early phase of adaptation and aftereffects (Buurke et al. 2019; Park et al. 2022). Taken together with these previous studies and the present study, external stabilization of walking balance would be important factor for locomotor adaptation. In order to maintain or reacquire conventional walking, balance support during walking should be carefully considered based on the user's physical fitness level and stage of rehabilitation..
Difference between WW and pole walking (PW)
In our previous study, using the same protocol as in the present study, we suggested that the neural control of pole walking (PW, a form of locomotion in which a person holds a pole in each hand) and CW are not independent (Obata et al. 2019, 2020). The results demonstrated that: (1) the degree of transfer to CW and PW was not different regardless of whether subjects adapted to the split belt in CW or PW; (2) PW washout trials completely washed out CW adaptation; (3) CW washout trials did not completely wash out PW adaptation.
As differences in neural controls between WW and PW as compared to CW, differences in posture and movement between these walking modes can be considered. PW is a form of locomotion in which a person holds a pole in each hand and touches one of the poles to the ground simultaneously with contralateral lower limb heel contact. Because the length of the poles was adjusted so that the elbow joint angles were approximately 90° when subjects stood upright with the poles (Obata et al. 2019), subjects could maintain a posture that is not different from that with CW, indicating that, at least in the lower limb, sensory inputs are also not different from those in CW. On the other hand, during WW, subjects are in a bending posture, as they put their weight on the rollator. Such a posture when using the walker would produce different sensory inputs between CW and PW.
Considering the plasticity of the locomotor neural circuit induced by sensory inputs, increased hip and knee flexions (Boyer et al. 2017), lower limb unloading (Costamagna et al. 2019), and reduced lower-limb muscle activities in WW (Suica et al. 2016) are likely to have a major impact on the specificity of the neural control between WW and CW. In gait rehabilitation, sensory inputs produced by the loading of the lower extremities and the extension of the hip joint have been reported to be two important factors in enhancing the locomotor circuit in the spinal cord (Pearson 2004; Dietz 2009). These differences in sensory inputs from lower limb loading and joint movements may cause the distinct neural control between CW and WW and the overlap of neural control between CW and PW.
Clinical significance (transfer to CW)
The difference in the degree of asymmetry between the initial and final points in Catch or Washout 1 inherently reflects that the de-adaptation of split-belt adaption occurs. However, in the transfer to CW after adaptation in WW (WW → CW), the degree of asymmetry at the initial points was not different between that at the final points. The result of no significant difference in the symmetry at the initial and final time points indicates that the transfer was very small or did not occur in this condition (WW → CW). Moreover, as compared to the degrees of asymmetry at the initial time points, the transfer to CW after adaptation in WW (WW → CW) was very small as compared to that to CW after adaptation in CW (CW → CW). Taken together, these results suggest that the effect of the split adaptation in WW is hardly retained in CW. When the adaptation is replaced with gait rehabilitation, the results imply that gait training with a walker is not effective for maintaining or restoring conventional bipedal walking.
Conclusion
The present results have shown that the effects of the split-belt adaptation were less transferable between CW and WW. The results suggest that rolling walkers are not necessarily appropriate as training for conventional walking because their neural control is different, whereas they are common walking aids for frail elderly people and useful for supporting their locomotion.
Data availability statement
The experimental data used for figures are available as open data via Kyushu Institute of Technology online data repository http://hdl.handle.net/10228/0002000389.
References
Bateni H, Maki BE (2005) Assistive devices for balance and mobility: benefits, demands, and adverse consequences. Arch Phys Med Rehabil 86(1):134–145
Bertrand K, Raymond MH, Miller WC, Ginis KAM, Demers L (2017) Walking aids for enabling activity and participation: a systematic review. Am J Phys Med Rehabil 96(12):894–903
Bradley SM, Hernandez CR (2011) Geriatric assistive devices. Am Fam Physician 84(4):405–411
Buurke TJW, Lamoth CJC, van der Woude LHV, den Otter R (2019) Handrail holding during treadmill walking reduces locomotor learning in able-bodied persons. IEEE Trans Neural Syst Rehabil Eng 27(9):1753–1759
Choi JT, Bastian AJ (2007) Adaptation reveals independent control networks for human walking. Nat Neurosci 10(8):1055–1062
Costamagna E, Thies SB, Kenney LPJ, Howard D, Lindemann U, Klenk J, Baker R (2019) Objective measures of rollator user stability and device loading during different walking scenarios. PLoS ONE 14(1):e0210960
Dietz V (2009) Body weight supported gait training: from laboratory to clinical setting. Brain Res Bull 78(1):1–6
Geravand M, Korondi PZ, Werner C, Hauer K, Peer A (2017) Human sit-to-stand transfer modeling towards intuitive and biologically-inspired robot assistance. Auton Robot 41(3):575–592
Jayaram G, Galea JM, Bastian AJ, Celnik P (2011) Human locomotor adaptive learning is proportional to depression of cerebellar excitability. Cereb Cortex 21(8):1901–1909
Jayaram G, Tang B, Pallegadda R, Vasudevan EV, Celnik P, Bastian A (2012) Modulating locomotor adaptation with cerebellar stimulation. J Neurophysiol 107(11):2950–2957
Liu HH (2009) Assessment of rolling walkers used by older adults in senior-living communities. Geriatr Gerontol Int 9(2):124–130
Malone LA, Bastian AJ (2010) Thinking about walking: effects of conscious correction versus distraction on locomotor adaptation. J Neurophysiol 103(4):1954–1962
Morton SM, Bastian AJ (2006) Cerebellar contributions to locomotor adaptations during splitbelt treadmill walking. J Neurosci 26(36):9107–9116
Obata H, Ogawa T, Nakazawa K (2019) Unique controlling mechanisms underlying walking with two handheld poles in contrast to those of conventional walking as revealed by split-belt locomotor adaptation. Exp Brain Res 237(7):1699–1707
Obata H, Ogawa T, Yokoyama H, Kaneko N, Nakazawa K (2020) Spatiotemporal characteristics of locomotor adaptation of walking with two handheld poles. Exp Brain Res 238(12):2973–2982
Ogawa T, Kawashima N, Ogata T, Nakazawa K (2012) Limited transfer of newly acquired movement patterns across walking and running in humans. PLoS ONE 7(9):e46349
Ogawa T, Kawashima N, Ogata T, Nakazawa K (2014) Predictive control of ankle stiffness at heel contact is a key element of locomotor adaptation during split-belt treadmill walking in humans. J Neurophysiol 111(4):722–732
Ogawa T, Kawashima N, Obata H, Kanosue K, Nakazawa K (2015) Distinct motor strategies underlying split-belt adaptation in human walking and running. PLoS ONE 10(3):e0121951
Ogawa T, Obata H, Yokoyama H, Kawashima N, Nakazawa K (2023) Different functional networks underlying human walking with pulling force fields acting in forward or backward directions. Sci Rep 13(1):1909. https://doi.org/10.1038/s41598-023-29231-6
Park S, Finley JM (2022) Manual stabilization reveals a transient role for balance control during locomotor adaptation. J Neurophysiol 128(4):808–818
Pearson KG (2004) Generating the walking gait: role of sensory feedback. Prog Brain Res 143:123–129
Reisman DS, Block HJ, Bastian AJ (2005) Interlimb coordination during locomotion: what can be adapted and stored? J Neurophysiol 94(4):2403–2415
Suica Z, Romkes J, Tal A, Maguire C (2016) Walking with a four wheeled walker (rollator) significantly reduces EMG lower-limb muscle activity in healthy subjects. J Bodyw Mov Ther 20(1):65–73
Taylor JA, Thoroughman KA (2007) Divided attention impairs human motor adaptation but not feedback control. J Neurophysiol 98(1):317–326
Vasudevan EV, Bastian AJ (2010) Split-belt treadmill adaptation shows different functional networks for fast and slow human walking. J Neurophysiol 103(1):183–191
Wolpaw JR (2007) Spinal cord plasticity in acquisition and maintenance of motor skills. Acta Physiol (oxf) 189(2):155–169
Acknowledgements
This work is supported by the Grant-in-Aid for Scientific Research (B) Grant Number JP21H03854 from Japan Society for the Promotion of Science (JSPS).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Additional information
Communicated by Francesco Lacquaniti.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Obata, H., Ogawa, T., Kaneko, N. et al. Distinct locomotor adaptation between conventional walking and walking with a walker. Exp Brain Res 242, 1861–1870 (2024). https://doi.org/10.1007/s00221-024-06863-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-024-06863-2