
Abstract
We addressed a hypothesis that changes in indices of finger interaction during maximal force production (MVC) tasks are accompanied by changed coordination of fingers in multi-finger accurate force production tasks. To modify relative involvement of extrinsic and intrinsic hand muscles, the subjects produced force by pressing either at their distal phalanges or at their proximal phalanges. As in earlier studies, in MVC trials, the elderly subjects showed a greater force decline when pressing at the proximal phalanges as compared to pressing at the distal phalanges. Two methods were applied to analyze finger coordination during the task of four-finger force production from zero to 30% of MVC over 5 s, at the level of finger forces (performance variables) and at the level of modes (control variables). Our previous observations of higher indices of variability during the ramp task in elderly subjects have been generalized to both sites of force application. An index of finger force covariation (the difference between the variance of the total force and the sum of the variances of individual finger forces) revealed small age related differences, which did not depend on the site of the force application. In contrast, analysis of covariation of force modes within the uncontrolled manifold (UCM) hypothesis showed much better stabilization of the time profile of the total force by young subjects. The UCM hypothesis was also used to test stabilization of the pronation/supination moment during the ramp task. Young subjects showed better moment stabilization than elderly. Age related differences in both force- and moment-stabilization effects were particularly strong during force application at the proximal phalanges. We conclude that the drop in MVC is accompanied in elderly subjects with worse coordination of control signals to fingers in multi-finger tasks. The UCM analysis was more powerful as compared to analysis of force variance profiles in revealing significant differences between the groups. This general result underscores the importance of efforts to analyze motor coordination using control rather than performance variables.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Aging leads to changes at many levels of the neuromotor hierarchy that participate in everyday hand activities. In particular, there is a progressive loss of the number of motor units in hand muscles accompanied by processes of reinnervation leading to the emergence of larger and slower motor units (Grabiner and Enoka 1995; Larsson and Ansved 1995). These processes are also accompanied by a drop in the muscle force and a general deterioration of the hand motor function (Christou et al. 2003; Cole et al. 1999; Ranganathan et al. 2001). Recent studies have suggested that age-related changes in the neuromotor apparatus are accompanied by adaptive changes in the control strategies that help alleviate the detrimental effects (DeVita and Hortobagyi 2000; Shinohara et al. 2003a).
Our recent studies (Shinohara et al. 2003a, 2003b) of finger interaction in tasks involving the production of maximal voluntary contraction (MVC) forces have shown that aging leads not only to a drop in MVC force but also to changes in indices of finger interaction such as force deficit (lower peak finger forces in multi-finger tasks as compared to single-finger tasks; Kinoshita et al. 1996; Li et al. 1998) and enslaving (force production by fingers that are not instructed to produce force; Kilbreath and Gandevia 1994; Li et al. 1998; Zatsiorsky et al. 2000; Slobounov et al. 2002). Elderly subjects showed lower peak forces, greater force deficit, and lower enslaving as compared to young subjects.
In one of those studies (Shinohara et al. 2003a), we performed analysis of the profiles of finger force variances computed over repetitions of a task that required accurate ramp force production. We were particularly interested in the relations between the sum of the variances of individual finger forces (∑VarF i ) and the variance of the total force (VarF TOT ). If the fingers deviate from their preferred (average) profiles independently of each other, one could expect ∑VarF i = VarF TOT (cf. Bienaymé equality theorem; Loève 1977). If, however, ∑VarF i >VarF TOT , one may conclude that there is predominantly negative covariation among finger forces such that they partly compensate for each other’s errors with respect to the total force profile (cf. Latash et al. 1998, 2001; Scholz et al. 2002). Both young and elderly subjects show no error compensation among the fingers at low forces and significant error compensation (in a sense that ∑VarF i >VarF TOT ) at higher forces: Independently of age and MVC, the subjects started to show error compensation when the total force reached about 5 N (Shinohara et al. 2003a). We hypothesized that this finding reflected the emergence of multi-finger synergies based on similar everyday tasks, such as drinking from the cup or eating with the spoon, that involve manipulation of objects whose weight is typically not scaled to the person’s age or force producing abilities.
There are two potentially important issues that remained unexplored in the previous study. First, finger forces are known to be dependent on each other because of the phenomenon of enslaving. Hence, interpretation of relations between ∑VarF i and VarF TOT is not trivial since some of them could be due to the enslaving, not to particular features of multi-finger control. As mentioned earlier, enslaving changes with age (it becomes weaker; Shinohara et al. 2003a, 2003b). Hence, the contribution of enslaving to the apparent relations between the two indices of finger force variance is likely to change with age. To overcome this problem, we have decided to use a method based on the uncontrolled manifold (UCM) hypothesis (Scholz and Schoöner 1999; reviewed in Latash et al. 2002c). According to the UCM approach, the controller acts in a state space of control variables and selects in that space a sub-space (a manifold, UCM) corresponding to a value of a performance variable that needs to be stabilized. Then, it arranges covariations among the control variables such that their variance has relatively little effect on the selected performance variable, i.e. it is mostly confined to the UCM.
The UCM hypothesis has been applied to analysis of finger interaction in force-producing tasks (Latash et al. 2001, 2002b; Scholz et al. 2002). That analysis was based on a set of hypothetical control variables, force modes (cf. Danion et al. 2003) accounting for enslaving among the fingers. In the current study, we compare the “more traditional” analysis that compares finger force variance profiles with UCM analysis of the same data. We hypothesized that there could be age-related differences in finger interaction during multi-finger tasks that the previous analysis failed to reveal because it was based on a set of variables (finger forces) that were mutually dependent.
Finger interaction has been discussed as resulting from both peripheral factors, such as shared muscles and muscle compartments and inter-digit tendinous connections (Leijnse et al. 1993; Kilbreath and Gandevia 1994), and from a neural organization of multi-finger control (Rouiller 1996). Finger flexion forces are produced by contraction of the finger-specific intrinsic hand muscles which attach to proximal phalanges and the extrinsic hand muscles with multi-digit tendons that attach to medial and distal phalanges (Landsmeer and Long 1965; Long 1965). This makes it possible to vary the relative involvement of the intrinsic and extrinsic hand muscles by changing the site of force production along the fingers, at the distal or at the proximal phalanges (Li et al. 2000; Latash et al. 2001). Earlier studies have shown larger indices of finger interaction during MVC force production at the proximal phalanges, speaking in favor of the mostly central nature of these phenomena (Latash et al. 2002a; Shinohara et al. 2003b). On the other hand, the study by Shinohara et al. (2003b) has shown that aging leads to a larger force loss in the intrinsic hand muscles as compared to the extrinsic muscles. There have been no studies comparing finger interaction during accurate force production trials at the distal and proximal phalanges. We hypothesized, therefore, that the differential effects of aging on the two muscle groups might result in changed finger interaction during submaximal accurate force production tasks, which might be viewed as more relevant to the everyday hand function.
Materials and methods
Subjects
Twelve young (28.9±4.4 years old) and 12 elderly (82.1±8.0 years old) subjects participated in the experiment. Each age group consisted of six males and six females. Within the present study, we did not analyze gender-related differences and concentrated on age-related changes common across the genders. All the subjects were healthy and right-handed according to their preferential use of the hand during writing, drawing, and eating. None of the subjects had a history of long-term involvement in activities such as typing and playing musical instruments. Elderly subjects were recruited from a local retirement community and passed the screening process that involved cognition test (Mini-mental status exam ≥24 points), depression test (Beck depression inventory ≤20 points), quantitative sensory test (monofilament ≤3.22) and general neurological examination. The height and body mass of the subjects were 169.1±6.9 cm and 65.1±14.4 kg in young subjects and 166.6±9.3 cm and 73.5±13.0 kg in elderly subjects, respectively. The length of the middle finger (from the MCP joint to the fingertip) and that of the proximal phalange of the middle finger (from the MCP joint to the proximal interphalangeal joint) was 10.2±0.7 cm and 5.4±0.3 cm in young subjects and 10.7±1.2 cm and 5.1±0.6 cm in elderly subjects, respectively. All the subjects gave informed consent according to the procedures approved by the Office for Regulatory Compliance of the Pennsylvania State University.
Apparatus
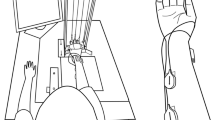
Four unidirectional piezoelectric sensors (Model 208C02, PCB Piezotronics, Inc., Depew, NY) were used to measure the forces produced by individual fingers. The sensors were each connected in series with wire cables that were suspended by swivel attachments from slots in the top plate of the inverted U-shaped frame of the experimental device (see Fig. 1 for a schematic illustration of the set-up). The slots were placed 30 mm apart from each other in the mediolateral direction and allowed fore-aft adjustment of the wires to accommodate individual subject’s differences in finger length. The fingers applied force to rubber-coated loops located at the bottom of each wire. These loops could be placed either against the middle of the distal phalanges (DP site) or against the middle of the proximal phalanges (PP site). Changes in the position of the loops were expected to change the relative contributions of the intrinsic and extrinsic hand muscles to force production (Li et al. 2000).

A schematic drawing of the setup for measuring finger flexion forces. The forearm was placed on the supporting surface, the hand was clamped by the fixation device, and the loops were positioned at the distal or proximal phalanges
Analog output signals from the sensors were sent to separate AC/DC conditioners (M482M66, PCB Piezotronics, Inc., Depew, NY). The signal conditioners operated in a DC-coupled mode, utilizing the sensor’s discharge time constant as established by the built-in microelectronic circuits within the sensors. As such, the time constant of the sensor was ≥500 s. The system involved approximately 1% error over the typical epoch of recording of a constant signal. A 16-bit A/D board (DAQ Card-AI-16XE-50, National Instruments, Austin, TX) was used to digitize the signals at 1,000 Hz. A Dell laptop computer controlled the experiment and was used for data acquisition and processing.
The subject was seated in a chair facing the testing table and a monitor with his/her right upper arm at approximately 45° of abduction in the frontal plane and approximately 45° of flexion in the sagittal plane, and the elbow at approximately 135° of flexion. The forearm was secured by Velcro straps flat on the supporting surface, that was at the same height as the support point of the hand fixation device. The hand fixation device was located at the bottom of the frame and was used to stabilize the palm of the hand and to ensure a constant hand configuration throughout the experiment. The wrist was at 20° of extension (hand and forearm aligned correspond to 0°). The MCP joints were at 20°. The thumb was positioned under the bar on which the palms rested. Due to the experimental procedure, all four finger forces were parallel to each other. All the precautions were taken to avoid motion of the forearm or hand during the tests without compromising the subjects’ comfort.
Procedure
Three types of tests were run. First, the subjects produced a brief maximal force with one finger at a time and with all four fingers together within a 2 s time interval shown on the screen (MVC tests). These tests were used to adjust the target in the ramp force production tasks. Then, the subjects were required to press with one finger such that the force signal shown on the screen followed a ramp line from zero to the peak force the finger showed in its MVC test within 5 s (Ramp-1 test). Results of these tests were used to calculate the enslaving matrix for the UCM analysis (see later). In each trial, all the fingers were in the loops and the subjects were explicitly instructed not to lift “uninvolved” fingers off the loops. The subjects were asked not to pay attention to possible force generation by those fingers as long as the force by the instructed finger followed the ramp line and reached its maximum. Both MVC and single-finger ramp tests were performed with the finger force application at both DP and PP sites. The order of the sites and the order of the finger(s) were balanced across subjects. The intervals between successive trials were 1 min, while the intervals between the conditions were about 5 min.
The main test involved pressing with all four fingers such that the total force followed a ramp template shown on the screen as closely as possible (Ramp-4 test). An oblique yellow line was shown on the screen corresponding to the force increase from zero to 30% of the four-finger MVC over 5 s, and the task was to trace this line in time with the cursor representing the total finger force. The forces of individual fingers were not shown on the screen, but they were recorded. This test was repeated 12 times for each site of force application, DP and PP (the order of sites was balanced across subjects). The intervals between successive trials were 15 s. Subjects never reported fatigue.
Data processing
The data were processed off-line. For the MVC tests, the peak force attained by the instructed finger(s) was determined.
For each Ramp-4 series, a normalization procedure was performed for all subjects and all conditions. The force ramp was plotted in Matlab and the time at which the force began to deviate from the baseline continuously toward its maximum was determined as the force onset. The time at which the force ramp reached its maximum was then chosen as the ramp termination. The force data within this period of time was then normalized to 100%, and average profiles of individual finger forces [F i (t)] and of the combined force [F TOT (t)] across the 12 trials were computed. Time profiles of the variances of the individual finger forces and of the total force across the 12 trials were also computed, ∑VarF i (t) and VarF TOT (t), respectively. Further, VarF i (t) time series were summed up to produce ∑VarF i (t).
The ramp task was split into five 1-s segments, and ∑VarF i (t) and VarF TOT (t) were averaged over each 1-s segment of the ramp for each subject separately. For across subjects comparisons, these indices were expressed in relative units after being divided by the square of the peak force value achieved by each subject in the four-finger MVC test. The difference between the two variance indices, ΔVar(t)=∑VarF i (t)–VarF TOT (t), was computed to assess the predominance of positive or negative covariations among individual finger forces. Note that if ΔVar(t) >0, negative covariations among finger forces dominate revealing (partial) compensation of errors introduced by individual fingers in separate trials. If ΔVar(t) <0, positive covariations dominate revealing amplification of errors introduced by individual fingers. For each subject, ΔVar(t) was averaged over each 1-s segment of the ramp and divided by the corresponding average value of ∑VarF i (t).
UCM analysis
Individual finger forces cannot be considered independent control variables manipulated by the central nervous system (CNS) because of the phenomenon of enslaving (Zatsiorsky et al. 1998). Hence, comparing the sum of variances of individual finger forces to the variance of the total force can lead to spurious effects, suggesting finger force covariation in the absence of covariation of control signals to the fingers. In this study, similarly to previous studies of multi-finger coordination using the UCM approach (Latash et al. 2002a, 2002b; Scholz et al. 2002), we transformed the force data into another set of variables, modes using enslaving matrices. The enslaving matrices were computed based on the Ramp-1 tests.
For each Ramp-1 trial, the onset of the ramp and time at which 40% of MVC was reached were determined. The latter value was chosen to ensure that the relations between individual finger forces and the total force were linear within the actual range of forces used in the four-finger ramp trials. The change of total force and of each individual finger force was determined over the defined time interval. A 4-by-4 one-hand enslaving matrix (ENSL) was then constructed:
where Δf j,k and ΔF k are the changes of individual finger force j [j = index (i), middle (m), ring (r), and little (l)] and the change of total force, respectively, produced during the ramp when finger k (k = i, m, r, and l) was the instructed master finger.
We will use a term “mode” for a hypothetical control variable corresponding to combinations of individual finger forces observed when a person tries to press with only one finger. Relations among changes in individual finger forces within a hand are assumed to be linear within the studied range of forces (cf. Li et al. 1998). Column k in the ENSL matrix describes a set of forces produced by individual fingers when the subject tries to produce the total force of 1 N by pressing only with finger k; i.e., it corresponds to mode-k = 1 N. Multi-finger force production is going to be considered as the process of specifying magnitudes of the modes for each of the explicitly involved fingers. A set of such magnitude factors may be viewed as a vector with the dimensionality equal to the number of explicitly involved fingers; we are going to address it as the m vector. Note that the introduced modes are measured in units of force and are not identical to dimensionless force modes described in earlier studies based on tests with maximal force production by one finger at a time (Danion et al. 2002). We did not take into account the phenomenon of force deficit (Li et al. 1998) in this analysis since it leads to proportional changes in the force of all four fingers in multi-finger tasks.
Our analysis addressed the following questions: Does covariation of mode magnitudes selectively stabilize a particular (average) time profile of the total force produced by the fingers? Does covariation of mode magnitudes selectively stabilize a particular (average) time profile of the total moment produced by the hand in the frontal plane? We will refer to these as force-control and moment-control hypotheses respectively. Ignoring the effects of enslaving, the following formulation of the relationship between variations in individual finger forces within a hand and changes in a selected performance variable (PV) would apply:
where d j are coefficients. For the force-control hypotheses, coefficients d j for the fingers were unity. For the moment-control hypothesis, these values represented lever arms for each finger force with respect to the longitudinal functional axis of the hand/forearm. The enslaving effect, however, induces a structure in the variability of individual finger forces that may be unrelated to a particular task variable. To eliminate such enslaving-induced correlations, we transform the individual finger forces into a set of mode magnitudes, m:
Change in a performance variable can now be expressed as a function of these variables:
We tested hypotheses that the average profile of the total force or moment was stabilized against fluctuations in finger forces. The analysis was performed at every 1% of the normalized force ramp duration. Some of the subjects, particularly elderly, showed considerable variations in the duration of their actually produced ramp profiles. Therefore, prior to further analysis, the time of the actual ramp for each trial was considered to be 100%. At each time, we assumed that the mean value of a performance variable across all trials represented the value that the CNS tried to stabilize. These values were calculated from the means of the individual finger forces across all task repetitions at each percent of the ramp duration. The mean values of the individual mode magnitudes constitute the reference m configurations for each interval of analysis. The linearized model accounting for effects of enslaving is described by Eq. 5.
An uncontrolled manifold (UCM) was computed in the space of the mean-free mode magnitudes. It represents combinations of mode magnitudes that are consistent with a stable value of a performance variable. The manifold is approximated linearly by the null space spanned by basis vectors, e i , solving the following equation:
Each hypothesis accounts for one dimension in the mode space, such that the null space is three-dimensional. The basis, e i , of the null space was computed numerically at each percentage of the ramp duration using MATLAB. The vector of individual mean-free mode magnitudes, obtained at each sample of the force ramp, was resolved into projection onto the null space:
and the component perpendicular to the null space:
The amount of variance per DOF within the UCM was estimated as:
where \( {\left| {f_{{{\text{II}}}} } \right|}^{2} \) is the squared length of the deviation vector lying within the linearized UCM, and n and d are the DOFs of the m vector and the performance variable, respectively.
Analogously, the amount of variance per DOF perpendicular to the UCM was estimated as:
The primary dependent variables used in subsequent analyses are \( \sigma ^{{\text{2}}}_{{{\text{II}}}} \;{\text{and}}\;\sigma ^{2}_{ \bot } \) per degree of freedom, and are referred to, respectively, as variance per DOF within the UCM (VarUCM) and orthogonal to it (VarORT). Further, a variable (ΔV) reflecting the difference between the variance within the UCM and orthogonal to the UCM was computed as:
where all variance indices are computed per degree of freedom; VarTOT means the total variance (Scholz et al. 2003).
Taking into account the phenomenon of enslaving is a major difference between the two approaches, the comparison of the sum of finger force variances to the variance of the total force vs. the UCM approach. If enslaving did not exist, the two types of analysis would produce identical results with respect to total force stabilization (force-control hypothesis) since force modes would lead to force production only by the targeted (master) fingers. In addition, the UCM approach allows stabilization of other performance variables to be addressed, such as the total moment produced by the fingers (moment-control hypothesis).
Statistics
Standard descriptive statistics and ANOVAs with repeated measures were used. Factors were chosen based on particular comparisons. Factors included age (elderly and young), site (PP and DP), performance variable (force or moment), and ramp segment. Level of significance was set at p = 0.05. The Newman-Keuls test was used for post hoc analysis. The data are presented as means and standard deviations in the text and as means and standard errors in the figures.
Results
This section is structured in the following way. First, we present results of tests with maximal force production (MVC) at the two sites of force application. These results are not novel but they form the basis for further analyses. In the second section, we describe analyses of the time profiles of variances of finger forces. These results are novel since they compare, for the first time, force variances during tests with force production at the two sites, DP and PP. The third section describes the results of the UCM analysis of finger forces using a set of hypothetical control variables, modes. Such analysis has not been done to compare the performance of young and elderly persons. Comparison of the results described in the second and third sections forms the core for further discussion.
MVC analysis
During four-finger trials with maximal force production (MVC trials), the peak total force was higher when the subjects pressed at their proximal phalanges (PP site) than when they pressed at the distal phalanges (DP site). Averaged across subjects, the peak force was 77.9±33.4 N at PP and 69.9±27.1 N at DP. Elderly subjects produced lower forces than young ones. Age related differences in MVC were site dependent. In young subjects, the average peak force was 85.9±36.7 N at PP while it was 72.7±30.2 N at DP. In elderly, the average peak force was 69.9±29.1 N at PP and 67.1±24.7 N at DP. In other words, the reduction in maximal force in elderly subject was more prominent at the PP site (19%) as compared to the DP site (8%). These findings were confirmed with a two-way age × site ANOVA with repeated measures. There were significant main effects of both factors (F (1,20)>13.0, p<0.01) and a significant age × site interaction (F (1,20)=5.75, p<0.05).
Analysis of force variance profiles
During the multi-finger force ramp production, errors in the performance were quantified using root mean square (RMS) from the target force over the time of the ramp averaged across trials. For comparison across subjects, the RMS index was normalized to the maximal force in the four-finger MVC test. The normalized RMS index for the elderly subjects was about twice as large as for the young subjects (0.84±0.48 vs. 0.43±0.03 respectively) independently of the site of force application. This difference was confirmed by a main effect of age (F (1,20)=9.13, p<0.01) in a two-way age × site ANOVA with repeated measures.
Two indices of force variance were computed for each set of ramp trials. The variance of total force (VarF TOT ) reflects the overall quality of performance in the task. The sum of the variances of the individual finger forces (∑VarF i ) reflects the total variance in the force space, which is expected to be equal to VarF TOT if all the fingers act as independent force generators. These indices were averaged over every 1-s segment of the force ramp and further divided by the maximal force squared to express them in dimensionless units.
The average values of these indices are shown in Fig. 2 for the young (open symbols) and elderly (filled symbols) subjects. Data for force production at PP are shown with circles, while data for trials at DP are shown with squares. VarF TOT was higher in elderly subjects than in young subjects, and at PP than at DP (Fig. 2A). These observations were confirmed by main effects of age (F (1,20)=8.89, p<0.01) and site (F (1,20)=19.55, p<0.01) in a three-way age × site × ramp segment ANOVA with repeated measures. Elderly subjects showed significantly larger VarF TOT values at PP, as compared to young subjects, as confirmed by the age × site interaction (F (1,20)=5.87, p<0.05). There was a main effect of ramp segment (F (4,80)=6.68, p<0.01), reflecting the greater VarF TOT for the last segment as compared to other segments (p<0.01).

VarF tot (A) and ∑VarF i (B) for the young (open symbols) and elderly (solid symbols) subjects at DP (squares) and PP (circles) are presented after being divided by the corresponding maximal force squared (the units are dimensionless multiplied by 10−4). C The difference between VarF tot and ∑VarF i (ΔVar) is presented, after being divided by ∑VarF i . The symbols are the same as in A and B. D ΔVar at DP (open circles) and PP (filled circles) are plotted against the actual mean force in each ramp segment for all four subject groups. Values are mean ± standard errors for A–C. The best-fit logarithmic curve is shown in panel D
The other index of variance, ∑VarF i , illustrated in panel B of Fig. 2, reached much higher magnitudes as compared to VarF TOT , particularly later in the ramp duration. ∑VarF i was significantly greater at PP as compared to DP, confirmed by the main effect of site (F (1,20)=7.70, p<0.05) in the three-way ANOVA with repeated measures. Age related differences did not reach the level of significance. ∑VarF i increased over the ramp duration (main effect of ramp segment, F (4,80)=57.91, p<0.001).
As described in the “Materials and methods,” comparison of the two indices of variance, VarF TOT and ∑VarF i , allows for assessing predominance of positive or negative covariations among individual finger forces across trials. The difference (ΔVar) between VarF TOT and ∑VarF i was analyzed after being divided by ∑VarF i to allow for across-subjects comparisons. The values of ΔVar averaged across subjects are shown in Fig. 2C. Over the first segment of the ramp, ΔVar was typically less than zero and then it increased over the ramp duration and became positive for the following segments. This was confirmed by the main effect of ramp segment (F (4,80)=222, p<0.001) in a three-way ANOVA with repeated measures. Elderly subjects had somewhat smaller ΔVar as compared to young subjects, confirmed by the main effect of age (F (1,20)=6.95, p<0.05). In contrast to the higher values for both VarF TOT and ∑VarF i at PP than at DP, ΔVar was not influenced by site of force application.
Since the young and elderly subject showed different MVC values, they produced the ramp task over different force ranges. Figure 2D shows the data for each subject plotted against the mean actual force produced over each of the ramp segments. The figure shows that negative values of ΔVar turn into positive values at approximately the same critical force value of about 3–4 N for both subject groups (3.17±2.83 N vs. 4.28±2.41 N for the young and elderly subjects respectively, non-significant). The data points for the ramp force production at the two sites, DP and PP, show similar distributions and turn from negative to positive values at about the same total force value. Since the subjects performed ramp force profiles over different magnitudes of the total force (scaled to their MVC), they reached similar critical force values over different times. The difference between the young and elderly subjects in their MVC translated into a significant difference in the time it took the subjects to reach the critical force level, 1.53±0.58 s vs. 1.95±0.5 s for the young and elderly subjects respectively (F (1,20)=7.19, p<0.05).
UCM analysis
UCM analysis was performed using modes, which unlike finger forces are assumed to be independently manipulated by the CNS. For the hypotheses of stabilization of the total force and total moment produced by the fingers, mode variance was partitioned into two components, within the UCM (VarUCM) and orthogonal to it (VarORT), each per degree of freedom of the corresponding subspaces. Further, the difference between the two variance indices was quantified using a normalized index ΔV. As mentioned in the “Materials and methods,” elderly subjects tended to show relatively high variability of the duration of the actual force ramps. Hence, the total duration of each trial was assumed to be 100%. For further analysis, average indices of variance were computed separately for each 20% (approximating five 1-s segments) of the normalized ramp duration.
Figure 3 illustrates averaged across subjects ΔV indices for each control hypothesis separately. In general, the moment-control hypothesis was not supported. The ΔV index was typically close to or below zero, except at the earliest segment of the force ramp. With respect to the force-control hypothesis, the subjects generally showed positive values of ΔV along the force ramp except for elderly subjects at the earliest segment of the ramp. Positive ΔV values suggest stabilization of the total force by covariations of modes to individual fingers.

The index of covariation of finger modes (ΔV) was computed with respect to the force-control (circles) and moment-control (triangles) hypotheses and averaged over each 1-s segment of the ramp separately. Further, these data were averaged across young (filled symbols) and elderly (open symbols) subjects. Young subjects show significantly higher values of ΔV for both hypotheses. Standard error bars are shown
In Fig. 3, young subjects (filled symbols) showed larger ΔV indices as compared to elderly subjects (open symbols) over the whole ramp duration and for both force-control (circles) and moment-control (triangles) hypotheses. The difference between young and elderly subjects was more pronounced early in the ramp for force-control and later in the ramp for moment-control. It is also obvious from Fig. 3 that ΔV values for the force-control hypothesis were higher than for the moment-control hypothesis at all ramp segments with the exception of the initial one.
These results were confirmed with a three-way age × hypothesis × ramp segment ANOVA, which showed significant effects of hypothesis (F (1,19)=50.9, p<0.001) and ramp segment (F (4,76)=67.7, p<0.0001) as well as significant age × ramp segment interactions (F (4,76)=3.1, p<0.05).
There were no significant main effects of the site of force application; hence, Fig. 3 shows data averaged over the PP and DP sites. However, there were significant age × site interactions (F (1,19)>5.4, p<0.05) for both force- and moment-control hypotheses, reflecting the fact that young subjects had higher ΔV than elderly subjects during force production at the PP site, but not at the DP site (p<0.01). This result is illustrated in Fig. 4 for both force-control and moment-control hypotheses. Note that both hypotheses could be satisfied simultaneously, particularly in young subjects during force application at the PP site.

The index of covariation of finger modes (ΔV) computed for the force-control and moment-control hypotheses for the two sites of force application, DP and PP. Young subjects (filled bars) show higher values of ΔV as compared to elderly subjects during force application at PP but not at DP. Mean values with standard error bars are shown
Typically, subjects showed negative ΔV values early in the ramp, which turned positive at higher forces. For each subject, we computed the percentage of the normalized ramp time when ΔV changed its sign. We will refer to this value as the critical time (T CR ). For a few cases, when ΔV was positive from the very beginning of the ramp, T CR was assumed to be zero. Figure 5A shows values of T CR averaged across subjects of each of the four mini-groups for force application at DP and at PP separately. Young subjects showed smaller values of T CR (0.075±0.021 s) as compared to elderly subjects (0.180±0.033 s; F (1,20)=10.5, p<0.01), while males (0.088±0.026) showed smaller T CR than females (0.160±0.031 s; F (1,20)=7.4, p<0.05), independent of the site of force application.

Along the ramp, ΔV index for the force-control hypothesis typically was negative early in the ramp and then became positive. A The percentage of the ramp when this happened is shown separately for the elderly and young subjects for the force application at the DP site (hatched bars) and at the PP site (solid bars). Elderly subjects took more time than young subjects to show positive ΔV values. B Similar differences were seen when the critical forces when ΔV reached zero were compared across the subject groups and sites of force application. Standard error bars are shown
For comparison with data presented in Fig. 2D, we computed absolute values of force at which ΔV changed its sign (Fig. 5B). Since all subjects performed ramp tasks scaled to their MVC values over the same time (5 s), they produced forces at different rates. When critical force levels were compared between the young and elderly subjects, significantly higher values were seen for the elderly (3.09±1.46 N vs. 1.62±1.44 N; p<0.05). There were no significant differences in these values across the two sites of force production.
Discussion
We applied two methods of analysis to study age related differences in finger interaction during accurate, slow, multi-finger force production tasks. The results have demonstrated that the UCM analysis was superior to analysis of time profiles of force variance in its ability to reveal significant differences across the subject groups. In particular, analysis of the variance profiles allowed the detection of significant age related effects in the behavior of an index (ΔVar) that reflected predominance of positive or negative covariation among finger forces (Fig. 2C, D), but it failed to detect differences between the proximal and distal sites of force application. In contrast, the UCM analysis revealed significant site related differences between the elderly and young subjects in their ability to organize covariation of control variables (modes) that stabilize a required time profile of the total force. The UCM analysis also allowed to address the stabilization of another potentially important variable, the total pronation/supination moment using the same set of data.
Age effects on the ability to coordinate finger action in a multi-finger task
Performance of our subjects in maximal force production (MVC) tests corroborated earlier findings on the lower peak forces produced by elderly subjects as compared to young subjects (Cole et al. 1999; Ranganathan et al. 2001; Shinohara et al. 2003a; Christou et al. 2003). This general loss of muscle force was accompanied in the elderly by a significantly worse performance in the accurate force production task: Their error indices (RMS) were nearly twice as high as those in the young subjects. Analysis of force variance profiles has also revealed that the elderly showed higher magnitudes of both variance indices, VarF TOT and ∑VarF i . There were smaller age related effects on the difference between the two indices, ΔVar, an index that reflected predominance of positive or negative covariations among individual finger forces: All subjects showed predominantly negative values of ΔVar at forces under 4 N and predominantly positive ΔV values at higher forces (also see Shinohara et al. 2003a).
The UCM analysis, however, revealed significant differences between the young and elderly subjects with respect to their ability to stabilize such performance variables as total force and total pronation/supination moment by a covariation of control signals (modes) to individual fingers. To assess this covariation, an index (ΔV) was computed representing normalized difference between the amount of mode variance per degree-of-freedom within the UCM and orthogonal to it (Scholz et al. 2003). Young subjects showed significantly higher values of ΔV for UCMs computed with respect to both total force and total moment corresponding to better stabilization of both performance variables. These results suggest that the ability of elderly subjects to stabilize important performance variables by covariation of control variables is lower than that of young subjects.
In the analysis of force variance profiles, the magnitude of total force when negative values of ΔVar turned into positive values was about the same (about 4 N) across the subject groups and sites of force production. Within the UCM analysis, however, an additional age-related difference has been revealed between the subjects. Elderly subjects took more time and reached higher forces before they were able to covary modes to stabilize the total force. Taken together, these results are the first of their kind in demonstrating an age-related deficit in the coordination of multi-effector actions using control variables rather than performance variables (cf. Ikeda et al. 1991; Cavanaugh et al. 1999; Cole and Rotella 2002).
Differential effects of age on instrinsic and extrinsic muscles
In earlier studies (S. Li et al. 2003; Shinohara et al. 2003b) we have suggested that aging is associated with more pronounced weakening of intrinsic hand muscles as compared to extrinsic muscles. This suggestion has followed a somewhat controversial hypothesis that proximal muscles lose less force with age as compared to distal muscles (Viitasalo et al. 1985; Christ et al. 1992; Era et al. 1992; Rice and Cunningham 2002). In the current study, we also observed a larger drop with age of the peak finger forces when the forces were applied at the PP site as compared to the DP site. At the DP site, the focal finger flexor is an extrinsic muscle, flexor digitorum profundis, while at the PP site, focal flexion force is produced by intrinsic digit-specific muscles (Long 1965). Hence, the larger drop in MVC during force production at PP may be interpreted as pointing at a larger loss of force with age of the intrinsic muscles as compared to extrinsic muscles.
In the UCM analysis of finger coordination during ramp tasks, there were no main effects of the site of force production. However, there was an interaction effect suggesting that the age related differences in peak forces of the intrinsic and extrinsic muscles are accompanied by worse stabilization of important performance variables. When the subjects produced forces at the fingertips (DP site), there were no differences between the young and elderly subjects in the index of stabilization computed for the total force or for the pronation/supination moment. In contrast, at the PP site, when intrinsic hand muscles were expected to be main focal force and moment generators, young subjects showed covariation of modes that stabilized both total force and total pronation/supination moment (ΔV >0), while elderly subjects showed worse force stabilization and failed to stabilize the moment (ΔV ≤0). Hence, one may conclude that age is associated with both a larger loss of muscle force by the intrinsic hand muscles and by their diminished ability to stabilize important performance variables during accurate force production. Although moment stabilization was not an explicit task component in the current study, it may be viewed as an important component for most everyday tasks that involve manipulation of hand-held objects (cf. Latash et al. 2001; Scholz et al. 2002).
Comparison of the results of the two types of analysis suggests that the drop in enslaving with age may be viewed as an adaptive factor helping to stabilize total force produced by the fingers. Indeed, total force stabilization requires predominantly negative covariation among finger forces. High enslaving is likely to result in positive covariation among the forces. In young subjects, modes to individual fingers needed to show strong negative covariation to overcome the effects of enslaving and to result in a net force stabilizing effect. If, as our results suggest, elderly subjects are deficient in their ability to organize adequate negative mode covariation, a drop in the enslaving may be viewed as another method by which to reach the net effect of negative covariation at the level of finger forces. Note, however, that enslaving may contribute to stabilization of the pronation/supination moment (cf. Zatsiorsky et al. 2000). As such, it may be viewed as a positive factor for moment stabilization resulting in better moment control in young subjects.
Multi-finger synergies and the structure of force variability
Multi-digit synergies have commonly been studied as correlated changes in the kinematic or force time patterns produced by a set of digits in response to a perturbation, over the time course of the task, or over manipulations of task parameters (Cole and Abbs 1987, 1988; Santello and Soechting 2000; Baud-Bovy and Soechting 2001, 2002; Reilmann et al. 2001; Rearick and Santello 2002; Gentilucci et al. 2003). Our approach differs in two major aspects. First, we focus on deviations of individual finger forces from their average performance over a series of trials. Second, the UCM analysis is always specific to a particular control hypothesis, i.e. a hypothesis on a performance variable that is supposed to be stabilized by covariation of control signals to a set of digits. As a result, our analysis addresses not a question of whether there is a multi-finger synergy but rather whether there is a synergy that stabilizes a particular potentially important variable.
Using motor variability as a window into motor coordination has a long history (for a review of earlier studies see Newell and Corcos 1993). However, until recently, progress in this direction has been slowed down because of the lack of a method to use indices of motor variability to quantify the cooperative efforts of a set of elements, for example fingers, towards a common goal. We view the introduction of the UCM hypothesis and its associated toolbox as a breakthrough, which allows, in particular, performance of quantitative analyses of motor synergies in special populations including elderly as in the current study.
A recent series of studies of the effects of aging on the structure of force variability during the isometric submaximal force production have shown that age leads to both an increase in the variability and a change in the timing structure of the force signal (Vaillancourt and Newell 2003; Vaillancourt et al. 2003). These studies, however, did not address coordination of force outputs across a set of digits. It is a challenge to understand relations between age-related changes in the structure of the motor variability of the outputs of individual elements and changes in the covariation of such outputs in a task-specific fashion.
Analysis of coordination using performance and control variables
Bernstein (1947, 1967) was arguably the first to emphasize that the central nervous system cannot control movements using performance variables such as forces, displacements, or levels of muscle activation. However, control variables have been notoriously hard to quantify and even to define. Any such attempts have induced heated debates (Latash 1993; Gottlieb 1998; Feldman et al. 1998). The temptation to use performance variables, which can be so readily recorded and related to task variables, has been too much to overcome, and an overwhelming majority of studies of motor coordination have used performance variables to detect and quantify motor synergies (among others, mea culpa, Scholz et al. 2000). Only recently, a few experimental studies of multi-effector coordination using hypothetical control variables have emerged (Latash et al. 2001; Scholz et al. 2002; Krishnamoorthy et al. 2003a, 2003b). One should certainly keep in mind that the control variables used in these studies, as well as the force modes used in the current study, remain hypothetical and not independently observable. As such, they are in an obvious disadvantage as compared to performance variables. Nevertheless, we view this as a temporary problem that may be overcome in future by progress in neurophysiological methods.
The different outcomes of the two methods of analysis used in this study illustrate the important differences between the two approaches. The methods differ in a number of important aspects. In particular, comparison of variance profiles assumes that finger forces potentially are independently controlled variables. However, the phenomenon of enslaving suggests that this is not true: When a subject tries to produce force with only one finger, all fingers show force changes (Li et al. 1998; Zatsiorsky et al. 1998, 2000). As such, the enslaving potentially contributes to positive covariation of finger forces independently of the task. This contribution of enslaving to the apparent covariation of finger forces depends on the strength of the enslaving effects. Since enslaving has been shown to be lower in elderly than in young persons (Shinohara et al. 2003a, 2003b), its contribution to force covariation can be expected to be age specific.
The UCM analysis uses a different set of variables for analysis, modes, which are assumed to be independent of each other (Latash et al. 2001; Scholz et al. 2002). They are computed based on finger forces taking into account the actual enslaving effects seen in each particular subject. Any observed covariation of modes may, therefore, be interpreted as reflecting task-specific features of control. The UCM analysis was more powerful as compared to analysis of force variance profiles in revealing significant differences between the subjects groups. This general result underscores the importance of approaches to analysis of motor coordination using control rather than performance variables.
References
Baud-Bovy G, Soechting JF (2001) Two virtual fingers in the control of the tripod grasp. J Neurophysiol 86:604–615
Baud-Bovy G, Soechting JF (2002) Factors influencing variability in load forces in a tripod grasp. Exp Brain Res 143:57–66
Bernstein NA (1947) On the construction of movements (in Russian). Medgiz, Moscow
Bernstein NA (1967) The co-ordination and regulation of movements. Pergamon Press, Oxford
Cavanaugh JT, Shinberg M, Ray L, Shipp KM, Kuchibhatla M, Schenkman M (1999) Kinematic characterization of standing reach: comparison of younger vs. older subjects. Clin Biomech 1999 14:271–279
Christ CB, Boileau RA, Slaughtr MH, Stillman RJ, Cameron JA, Massey BH (1992) Maximal voluntary isometric force production characteristics of six muscle groups in women aged 25 to 74 years. Am J Hum Biol 4:537–545
Christou EA, Shinohara M, Enoka RM (2003) Fluctuations in acceleration during voluntary contractions lead to greater impairment of movement accuracy in old adults. J Appl Physiol 95:373–384
Cole KJ, Abbs JH (1987) Kinematic and electromyographic responses to perturbation of a rapid grasp. J Neurophysiol 57:1498–1510
Cole KJ, Abbs JH (1988) Grip force adjustments evoked by load force perturbations of a grasped object. J Neurophysiol 60:1513–1522
Cole KJ, Rotella DL (2002) Old age impairs the use of arbitrary visual cues for predictive control of fingertip forces during grasp. Exp Brain Res 143:35–41
Cole KJ, Rotella DL, Harper JG (1999) Mechanisms for age-related changes of fingertip forces during precision gripping and lifting in adults. J Neurosci 19:3238–3247
Danion F, Schöner G, Latash ML, Li S, Scholz JP, Zatsiorsky VM (2003) A force mode hypothesis for finger interaction during multi-finger force production tasks. Biol Cybern 88:91–98
DeVita P, Hortobagyi T (2000) Age causes a redistribution of joint torques and powers during gait. J Appl Physiol 88:1804–1811
Era P, Lyyra AL, Viitasalo JT, Heikkinen E (1992) Determinants of isometric muscle strength in men of different ages. Eur J Appl Physiol Occup Physiol 64:84–91
Feldman AG, Ostry DJ, Levin MF, Gribble PL, Mitnitski AB (1998a) Recent tests of the equilibrium-point hypothesis (λ model). Motor Control 2:189–205
Gentilucci M, Caselli L, Secchi C (2003) Finger control in the tripod grasp. Exp Brain Res 149:351–360
Gottlieb GL (1998) Rejecting the equilibrium-point hypothesis. Motor Control 2:10–12
Grabiner MD, Enoka RM (1995) Changes in movement capabilities with aging. Exerc Sport Sci Rev 23:65–104
Ikeda ER, Schenkman ML, Riley PO, Hodge WA (1991) Influence of age on dynamics of rising from a chair. Phys Ther 71:473–481
Kilbreath SL, Gandevia SC (1994) Limited independent flexion of the thumb and fingers in human subjects. J Physiol 479:487–497
Kinoshita H, Murase T, Bandou T (1996) Grip posture and forces during holding cylindrical objects with circular grips. Ergonomics 39:1163–1176
Krishnamoorthy V, Goodman SR, Latash ML, Zatsiorsky VM (2003a) Muscle synergies during shifts of the center of pressure by standing persons: Identification of muscle modes. Biol Cybern 89:152–161
Krishnamoorthy V, Latash ML, Scholz JP, Zatsiorsky VM (2003b) Muscle synergies during shifts of the center of pressure by standing persons. Exp Brain Res 152:281–292
Landsmeer JM, Long C (1965) The mechanism of finger control, based on electromyograms and location analysis. Acta Anat 60:330–347
Larsson L, Ansved T (1995) Effects of ageing on the motor unit. Prog Neurobiol 45:397–458
Latash ML (1993) Control of human movement. Human Kinetics, Urbana, IL
Latash ML, Li Z-M, Zatsiorsky VM (1998) A principle of error compensation studied within a task of force production by a redundant set of fingers. Exp Brain Res 122:131–138
Latash ML, Scholz JP, Danion F, Schöner G (2001) Structure of motor variability in marginally redundant multi-finger force production tasks. Exp Brain Res 141:153–165
Latash ML, Li S, Danion F, Zatsiorsky VM (2002a) Central mechanisms of finger interaction during one- and two-hand force production at distal and proximal phalanges. Brain Res 924:198–208
Latash ML, Scholz JF, Danion F, Schöner G (2002b) Finger coordination during discrete and oscillatory force production tasks. Exp Brain Res 146:412–432
Latash ML, Scholz JP, Schöner G (2002c) Motor control strategies revealed in the structure of motor variability. Exer Sport Sci Rev 30:26–31
Leijnse JN, Snijders CJ, Bonte JE, Landsmeer JM, Kalker JJ, Van der Meulen JC, Sonneveld GJ, Hovius SE (1993) The hand of the musician: the kinematics of the bidigital finger system with anatomical restrictions. J Biomech 26:1169–1179
Li Z-M, Latash ML, Zatsiorsky VM (1998) Force sharing among fingers as a model of the redundancy problem. Exp Brain Res 119:276–286
Li Z-M, Zatsiorsky VM, Latash ML (2000) Contribution of the extrinsic and intrinsic hand muscles to the moments in finger joints. Clin Biomech 15:203–211
Li S, Latash ML, Yue GH, Siemionow V, Sahgal V (2003) The effects of stroke and age on finger interaction in multi-finger force production tasks. Clin Neurophysiol 114:1646–1655
Loève M (1977) Probability theory I, 4th edn. Springer-Verlag, New York
Long C (1965) Intrinsic-extrinsic muscle control of the fingers. J Bone Joint Surg Am 50:973–984
Newell KM, Corcos DM (eds) (1993) Variability in motor control. Human Kinetics, Urbana, IL
Ranganathan VK, Siemionow V, Sahgal V, Yue GH (2001) Effects of aging on hand function. J Am Geriatr Soc 49:1478–1484
Rearick MP, Santello M (2002) Force synergies for multifingered grasping: effect of predictability in object center of mass and handedness. Exp Brain Res 144:38–49
Reilmann R, Gordon AM, Henningsen H (2001) Initiation and development of fingertip forces during whole-hand grasping. Exp Brain Res 140:443–452
Rice CL, Cunningham DA (2002) Aging of the neuromuscular system: influences of gender and physical activity. In: Shephard RJ (ed) Gender, physical activity, and aging. CRC Press, Boca Raton, FL, pp 121–150
Santello M, Soechting JF (2000) Force synergies for multifingered grasping. Exp Brain Res 133:457–467
Scholz JP, Schöner G (1999) The uncontrolled manifold concept: identifying control variables for a functional task. Exp Brain Res 126:289–306
Scholz JP, Schöner G, Latash ML (2000) Identifying the control structure of multijoint coordination during pistol shooting. Exp Brain Res 135:382–404
Scholz JP, Danion F, Latash ML, Schöner G (2002) Understanding finger coordination through analysis of the structure of force variability. Biol Cybern 86:29–39
Scholz JP, Kang N, Patterson D, Latash ML (2003) Uncontrolled manifold analysis of single trials during multi-finger force production by persons with and without Down syndrome. Exp Brain Res 153:45–58
Shinohara M, Li S, Kang N, Zatsiorsky VM, Latash ML (2003a) Effects of age and gender on finger coordination in maximal contractions and submaximal force matching tasks. J Appl Physiol 94:259–270
Shinohara M, Latash ML, Zatsiorsky VM (2003b) Age effects on force production by the intrinsic and extrinsic hand muscles and finger interaction during maximal contraction tasks. J Appl Physiol 95:1361–1369
Slobounov S, Chiang H, Johnston J, Ray W (2002) Modulated cortical control of individual fingers in experienced musicians: an EEG study. Clin Neurophysiol 113:2013–2024
Vaillancourt DE, Newell KM (2003) Aging and the time and frequency structure of force output variability. J Appl Physiol 94:903–912
Vaillancourt DE, Larsson L, Newell KM (2003) Effects of aging on force variability, single motor unit discharge patterns, and the structure of 10, 20, and 40 Hz EMG activity. Neurobiol Aging 24:25–35
Viitasalo JT, Era P, Leskinen AL, Heikkinen E (1985) Muscular strength profiles and anthropometry in random samples of men aged 31–35, 51–55 and 71–75 years. Ergonomics 28:1563–1574
Zatsiorsky VM, Li Z-M, Latash ML (1998) Coordinated force production in multi-finger tasks. Finger interaction, enslaving effects, and neural network modeling. Biol Cybern 79:139–150
Zatsiorsky VM, Li Z-M, Latash ML (2000) Enslaving effects in multi-finger force production. Exp Brain Res 131:187–195
Acknowledgements
This study was supported in part by NIH grants AG-018751, NS-35032, AR 048563, and M01 RR10732 and NSF grant IBN-0078127. The screening process of the elderly subjects was conducted by Candace Kugel and Elizabeth Abraham under the supervision by Harold Bassett at the General Clinical Research Center (The Pennsylvania State University). The authors are grateful to the staff and participants at the Foxdale Village (State College, PA) for their cooperation.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Shinohara, M., Scholz, J.P., Zatsiorsky, V.M. et al. Finger interaction during accurate multi-finger force production tasks in young and elderly persons. Exp Brain Res 156, 282–292 (2004). https://doi.org/10.1007/s00221-003-1786-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-003-1786-9