
Abstract
Rationale
Paired associates learning (PAL) has been suggested to be predictive of functional outcomes in first episode psychosis and of conversion from mild cognitive impairment to Alzheimer’s disease. An automated touch screen-based rodent PAL (rPAL) task has been developed and is sensitive to manipulations of the dopaminergic and glutamatergic system. Accordingly, rPAL when used with pharmacological models of schizophrenia, like NMDA receptor blockade with MK-801 or dopaminergic stimulation with amphetamine, may have utility as a translational model of cognitive impairment in schizophrenia.
Objective
The purpose of this study was to determine if amphetamine- and MK-801-induced impairment represent distinct models of cognitive impairment by testing their sensitivity to common antipsychotics and determine the relative contributions of D1 versus D2 receptors on performance of PAL.
Method
Rats were trained in rPAL and were then treated with MK-801, amphetamine, risperidone, haloperidol, quinpirole, SK-82958, or SCH-23390 alone and in combination.
Results
While both amphetamine and MK-801 caused clear impairments in accuracy, MK-801 induced a profound “perseverative” type behavior that was more pronounced when compared to amphetamine. Moreover, amphetamine-induced impairments, but not the effects of MK-801, could be reversed by antipsychotics as well as the D1 receptor antagonist SCH-23390, suggesting a role for both the D1 and D2 receptor in the amphetamine impairment model.
Conclusions
These data suggest that amphetamine and MK-801 represent dissociable models of impairment in PAL, dependent on different underlying neurobiology. The ability to distinguish dopaminergic versus glutamatergic effects on performance in rPAL makes it a unique and useful tool in the modeling of cognitive impairments in schizophrenia.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
A difficulty in the study of cognitive disorders in general, and schizophrenia specifically, has been the paucity of meaningful and predictive pre-clinical models of the cognitive deficits associated with the disease. A common criticism of pre-clinical research is that the methods and measures used bare so little resemblance to those used in a clinical setting that they fail as predictive measures (Markou et al. 2009). Yet, it may be possible to improve predictive reliability by using a “translational “approach, testing rodents in a more human-like fashion and using human batteries that can be more effectively modeled in the rodent (Talpos and Steckler 2013).
A good example of the iterative translational approach is work being performed with tests of paired associates learning (PAL). PAL, as part of the Cambridge Neurological Test Battery (CANTAB), is sensitive to first episode psychosis and may be predictive of functional outcomes (Barnett et al. 2005, 2010). As part of the CANTAB battery, participants are shown a series of items placed in distinct locations on a computer monitor (sample phase). These same objects are then shown in the center of the monitor, and participants are asked to select where the object was previously located (Blackwell et al. 2004). Talpos and colleagues developed a rodent object-in-place PAL task (rPAL) that, despite being slowly learned, is sensitive to glutamatergic manipulations of the hippocampus, while a very closely related control task that could have been solved via complex conditional discrimination was not (2009). In rPAL, rats or mice are presented with 2 of 3 stimuli displayed in 2 of 3 possible locations and are required to learn that a certain object is only ever correct when displayed in a specific location in a two-choice discrimination task (Talpos et al. 2009). More recently, rPAL has also been adapted for use in humans, allowing near identical tests to be used in human and rodent research (as reported in Nithianantharajah et al. 2013). In part, because of the translational nature of rPAL, it has been recommended as useful paradigm for research into cognition in schizophrenia by a consortium on Cognitive Neuroscience Treatment Research to Improve Cognition in Schizophrenia (CNTRICS; Bussey et al. 2013). Certainly, there are differences between rPAL and PAL as part of CANTAB. Most notable of these is that the object-location association is built very slowly in rPAL, but may be established in as little as one pairing in CANTAB PAL. However, what the tasks do share in common is the requirement for the recall of an association made across modalities (spatial and visual).
Recently, the effect of pharmacological agents used to mimic or induce specific aspects of disease in rPAL has been established (Talpos et al. 2014). These included popular pharmacological models of dopaminergic or glutaminergic dysfunction in schizophrenia such as acute administration of amphetamine or NMDA antagonists. Interestingly, amphetamine was capable of robustly decreasing correct responses while having minimal effects on response latency. In contrast, PCP caused only a small decrease in accuracy whereas ketamine had little or no effect on accuracy. Yet, both NMDA receptor antagonists preferentially increased response latencies over collection latencies. The more pronounced effect on response as opposed to collection latency may suggest that NMDA receptor blockade leads to cognitive slowing as opposed to non-cognitive behavioral changes. A specific role of the NMDA receptors in the modulation of cognition in PAL is further bolstered by findings from Ballard et al. (2013), showing that MK-801 disrupted cognitive measures in the task (percent correct responses), while the effects of MK-801 on a simple visual discrimination were more limited, primarily affecting responsivity. These findings are in line with the results of the first rPAL study which showed that direct administration of MK-801 into the dorsal hippocampus disrupted percent correct in PAL (Talpos et al. 2009). Accordingly, it appears that amphetamine, and to varying degrees NMDA receptor antagonists, can be used as models of cognitive impairment in rPAL. The use of amphetamine or an MK-801 challenge to induce cognitive deficits in rPAL is in line with the two primary neurochemical theories of schizophrenia that view the disease as primarily a disorder of the dopaminergic, or glutamatergic, systems.
While the dopamine and glutamate hypotheses of schizophrenia are often presented as two independent central dogmas to understanding schizophrenia (Abi-Dargham 2004; Angrist et al. 1974; Brisch et al. 2014; Carlsson 1988; Gilmour et al. 2012; Krystal et al. 2003; Lau et al. 2013; Tamminga 1998), the reality is schizophrenia is a heterogeneous disorder and therefore, it is very likely that elements of both hypotheses are correct. Accordingly, the sensitivity of rPAL to glutamatergic and dopaminergic challenges may be useful in the search for novel pharmacological treatments for schizophrenia, especially if the profiles of these impairments differ as this may allow the modeling of different underlying pathologies and symptom clusters (positive versus cognitive), within the same testing environment. This could be of great benefit in determining the impact of positive symptoms and D2 receptor-based antipsychotics on cognition, as well as providing a new avenue for development of treatments to normalize glutamatergic functioning.
If the effects reported by Ballard et al. (2013) can be replicated, then MK-801 and amphetamine may serve as effective and dissociable models of glutamatergic and dopaminergic cognitive dysfunction in schizophrenia. Here, we attempt to replicate previously reported results and extend these by determining if the effects of amphetamine or MK-801 can be reversed by the commonly used antipsychotics haloperidol and risperidone. We predict that haloperidol and risperidone will be able to partially reverse the effects observed with amphetamine owing to their ability to normalize hyper-dopaminergic function. In contrast, we expect these compounds to have little effect on the dysfunction within the glutamatergic system induced by MK-801 as this may be more related to changes within the hippocampus and not related to the D2 receptor. Furthermore, through the use of compounds selective for different dopamine receptor subtypes (the D1 antagonist SCH-23390, the D1 agonist SKF-82958, and the D2 agonist quinpirole), we attempt to elucidate the relative contributions of the D1 and D2 receptor subtypes to the impairments in accuracy caused by amphetamine. To do this, haloperidol and risperidone, as well as SCH-23390 and quinpirole were tested against amphetamine to see if they could reverse amphetamine-induced deficits. Moreover, agonists SFK-82958 and quinpirole were tested on their own to determine if selective dopamine receptor activation could give similar impairments to that of amphetamine. By performing this work, we hope to determine if MK-801 and amphetamine represent unique models of disease when used in rPAL.
Material and methods
Subjects
All animals were treated in accordance with the European Ethics Committee (decree 86/609/CEE), the Animal Welfare Act (1 USC 2131) and the Guidelines for the Care and Use of animals in Neuroscience and Behavioral Research (National Research Council 2003). The study protocol was approved by the local animal experimental ethical committee at Janssen Research and Development (Beerse, Belgium).
Lister-Hooded rats (Harlan, Netherlands: 180–200 g at arrival, 72 rats in total) were used for all work reported in this manuscript. Upon arrival, animals were housed four per cage in individually ventilated type four cages (1400 cm2). These cages were filled with sawdust, and animals were also given chewing blocks and red plastic tunnels as environmental enrichment. Rats were fed daily after testing and given enough food to maintain them at 85–90 % of free feed body weight (typically 15 g a day). Animals were given free access to water except during testing. Upon arrival, animals were given 1 week to acclimatize to their new setting before being placed on food restriction. Compounds were typically tested on Tuesdays or Fridays with Mondays and Thursdays being used to ensure that the animals’ performance had returned to baseline after the previous treatment, thus a standard study would take 2 or 3 weeks to complete. Forty-eight animals were used for the amphetamine and dopaminergic studies, whereas 24 animals were used for the MK-801 studies. In both instances, experimental sample size was 11 or 12 rats. Animals were reused in multiple experiments. In order to be considered as “trained”, an animal had to perform at least 65 % correct over the three previous sessions, with a standard error of the mean less the 10 % of average performance, and complete at least 67 of 72 trials.
Compounds
All compounds were administered in a volume of 1 mg/kg and given via the sub-cutaneous route. Amphetamine was administered 60 min prior to testing in a saline vehicle (synthesized internally). MK-801 (Tocris Cookson) was administered 30 min prior to testing (saline vehicle) for the dose response studies, but 60 min prior to testing in reversal studies to make the studies more comparable to amphetamine studies. Risperidone and Haloperidol (synthesized internally) were dissolved with tartaric acid and administered in a saline solution 30 min prior to testing. Quinpirole (Sigma) was administered 30 min prior to testing in saline. SKF-82958 (3B Pharmachem) was dissolved with tartaric acid and administered in a 10 % cyclodextrin saline solution. SCH-23390 (Tocris Cookson) was administered 30 min prior to testing in a saline solution. Please see Table 1.
Apparatus
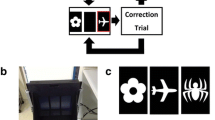
The apparatus used here was identical to that used in Talpos et al. (2014). All experiments were performed in modified Med Associates operant chambers (Med associates Inc. Fairfax, Vermont; 33.5 cm H × 32.5 cm W × 40 cm l). These chambers were equipped with a tone generator, a house light and a pellet receptacle, including a reward light, as well as a pellet dispenser. One wall of the chamber was replaced with a touch-sensitive computer monitor that was partly covered with a mask, restricting responses to three areas on the screen. A “flap” was placed in front of the screen to slow the response of the rat. Screens and boxes were controlled by K-limbic software (version 1.21.3.3; Conclusive Solutions, Sawbridgeworth, UK).
Training
Animals were trained to complete the PAL task in a stepwise manner. Rats are initially encouraged to explore the screen by placing a mixture of peanut butter and reward pellets upon the screen. Once animals reliably ate the pellets from the screen (two sessions), they were trained to associate a tone with the delivery of a food pellet reward. This was done by pairing the delivery of a food pellet with the sounding of a tone and activation of the food receptacle light. Once the pellet was collected, a 30-s inter-trial interval (ITI) started, and at the end of this interval, another pellet was delivered. A session lasted for 60 min or 60 trials, whichever occurred first. Once this had been achieved, rats were trained to touch the illuminated screen (anywhere) to earn a pellet reward. To do this, a trial began with the illumination of the screen. Once a response was made at any part of the screen, the screen would go dark, a tone would sound, the magazine light would turn on, and a food pellet would be delivered. Collection of the pellet would cause the magazine light to turn off and start a short ITI (10 s). Once the ITI had passed, the magazine light would again illuminate. A nose poke at the magazine light would cause it to turn off and the screen to again illuminate. A session lasted for 60 trials or 60 min, whichever occurred first. Once animals had learned to screen-touch, they are trained to respond to a smaller, illuminated portion of the screen to earn a reward (only 1 of 3 areas on the screen was illuminated). Once rats have learned to respond to different areas, depending on whether they were illuminated or not, rats were placed on the full version of the task.
In the full PAL, task rodents must learn that a given image was correct only when displayed in a specific location. On any trial, two out of three images were shown in two of three locations upon the screen. Each image was associated with reward only when paired with a distinct location, and on any given trial, one image was paired with its correct location (where a response resulted in a reward), whereas the other was paired with an incongruent location (where a response resulted in a timeout period before the start of the next trial). Accordingly, a total of 6 trial types were possible (pairs with two correct or incorrect outcomes were not included within this study). A session started with the delivery of a food pellet, the activation of the house and reward lights, as well as the sounding of the reward tone. Once the animal nose poked at the pellet receptacle, the reward light was turned off and two stimuli were displayed on the screen. A correct response at the screen resulted in the delivery of a food pellet with the simultaneous activation of reward magazine light and the sounding of the reward tone (0.5 s). Once the pellet was collected, a 10-s inter-trial interval (ITI) began. At the end of this period, the magazine light was again illuminated and a response to the magazine initiated the next trial. If, however, an incorrect response was made, the images were extinguished from the screen and the house light was turned off, signaling a timeout period (10 s). Once this timeout period had lapsed, the next ITI started. Incorrect trials were followed by a repetition of the previous trial, so-called correction trials, until a correct response was made. These correction trials were not counted towards the total trials completed, nor were they used in accuracy or latency calculations, but merely served as reminder for the correct stimulus/location association. A session was completed after 72 trials or 45 min, whichever occurred first. For a pictorial description of the stimulus location pairings, see Talpos et al. (2009).
Statistical methods
Percent correct ((correct trials/total trials)*100), percent trials completed ((completed trials/trials scheduled per session)*100), response latency (time from onset of stimuli to response in Log 10 msec), and collection latency (time from a correct response to the collection of reward in Log 10 msec) served as the primary measures of performance. Repeated measure ANOVAs were used to detect main effects of drugs. Post hoc analysis was performed with Dunnett’s t tests, with comparisons being made against vehicle or vehicle/challenge as appropriate. In instances where no variance existed within a treatment group (for example, percent trials completed under the vehicle condition), paired sample t tests were used instead of ANOVAs (an analysis of variance cannot be performed without variance). As in our previous study using these methodologies (Talpos et al. 2014), animals completing fewer than 20 trials were excluded from statistical analysis, except for trials completed and comparisons of standard trials versus correction trials. Correction trials were removed from analysis of all measures, except for a comparison of performance on standard trials versus correction trials after MK-801 or amphetamine treatment. In these instances, calculations were performed on the basis group errors, rather than individual animal errors, using a χ 2 approach. While this statistical approach is limited, it circumvented the problem caused by animals that made relatively few errors.
Results
MK-801 dose response
MK-801, given prior to testing in PAL, had a significant effect on percent correct (F(3,66) = 31.55, P < 0.001). Post hoc analysis indicated that all doses significantly lowered percent correct when compared to the vehicle control, (see Fig. 1, Table 2 supplementary material). MK-801 caused a significant decrease in trials completed (F(3,66) = 7.09, P < 0.001). Post hoc analysis indicated that a significant difference existed between only vehicle and the 0.075 mg/kg condition (P < 0.001). Similarly, while MK-801 had a significant effect on response latency (F(3,66) = 13.78, P < 0.001), post hoc analysis indicated that only 0.075 mg/kg was significantly increased when compared to vehicle (P < 0.001). MK-801 also had a significant effect on collection latency (F(3,66) = 15.54, P < 0.001), while the lowest dose (0.025 mg/kg) caused a small decrease in latency (P = 0.025), the highest dose caused a small increase in latency (P < 0.001). A series of χ 2 tests suggested that MK-801 had a preferential effect on correction trials. While no significant differences were seen on total group errors under the vehicle condition (χ 2(1,2005) = 1.09, P = 0.29) or at the 0.025 mg/kg dose (χ 2(1,2122) = 2.02, P = 0.155), dramatic decreases were seen at doses of 0.05 (χ 2(1,2343) = 41.77, P < 0.00001) and 0.075 mg/kg (χ 2(1,2277) = 11.79, P = 0.0006).

MK-801 induced a significant decrease in accuracy as well as trials completed (a). The results of a series χ 2 tests suggest that MK-801 has a greater effect on correction trials than standard trials (b; total group errors). Risperidone (c) and haloperidol (d) were not able to reverse changes induced by MK-801. *P < 0.05; **P < 0.01; ***P < 0.001
Amphetamine dose response
The effects of amphetamine have previously been reported in Talpos et al. (2014; see Fig. 2 and table 2 supplementary material for summary of these data) and are included here only for illustrative purposes. However, an additional analysis was done, comparing performance on standard trials versus correction trials after administration of amphetamine. No difference between the groups was seen under vehicle conditions (χ 2(1,1009) = 1.20, P = 0.27) or after a 0.25 mg/kg dose (χ 2(1,1244) = 2.93, P = 0.08). However, significant differences were observed at 0.5 mg/kg (χ 2(1,1249) = 8.55, P = 0.0035), and 0.75 mg/kg (χ 2(1,846) = 4.39, P = 0.03) of amphetamine, where amphetamine treated animals were worse on correction trials.

Amphetamine induced a significant decrease in accuracy in PAL (a). Amphetamine appeared to have a modest preferential effect on correction trial errors (b; group errors). Risperidone (c) and haloperidol (d) were able to partial reverse this impairment. *P < 0.05; **P < 0.01; ***P < 0.001. The data in Fig. 2a have previously been reported in Talpos et al. 2014
The effects of the D1 agonist SKF-82958 on baseline PAL performance
SKF-82958 caused a near significant decrease in percent correct (F(3,27) = 2.390, P = 0.09). No effect was seen on response latency (F(3,27) = 1.83, P = 0.16) or collection latency (F(3,27) = 0.15, P = 0.928). All animals completed all trials under all conditions (Table 4 supplementary material).
The effects of the D1 antagonist SCH-23390 on baseline PAL performance
SCH-23390 caused a significant reduction on trials completed at all doses tested (0.0025 mg/kg T(11) = 2.257, P = 0.04; 0.005 mg/kg T(11) = 4.562, P < 0.001, 0.01 mg/g T(11) = 3.08, P = 0.01). Because of the low number of trials completed, only the effects on percent correct caused by the lowest dose of SCH-23390 were analyzed by ANOVA. No effect of SCH-23390 on percent correct was seen using this analysis (F(1,11) = 2.7705, P = 0.12). To also allow analysis of the higher doses, a very liberal approach was taken, where each dose was individually compared to vehicle, thus maximizing the number of data points that could be included in analysis. However, no significant difference from vehicle was observed at the 0.005 or the 0.01 mg/kg condition (Table 4 supplementary material).
The effects of the D2 agonist quinpirole on baseline PAL performance
When administered on its own, quinpirole caused a reduction in trials completed at all doses tested (0.125 mg/kg T(11) = 2.45, P = 0.03; 0.25 mg/kg T(11) = 4.73, P < 0.001, 0.5 mg/kg T(11) = 20.16, P < 0.001). A significant effect of quinpirole was detected on percent correct responses (F(3,18) = 7.828, P = 0.002), where the highest dose of quinpirole was associated with a significant decrease in percent correct (P = 0.002). Inclusion of the highest dose in the analysis limited the total numbers of animals that could be included in this study. Accordingly, an additional ANOVA was performed, including the 0.125 and 0.25 mg/kg doses, but not the 0.5 mg/kg dose, to increase sample size. 0.125 and 0.25 mg/kg of quinpirole were not associated with significant changes from vehicle.
Quinpirole had a clear effect on response latency (F(3,18) = 12.50, P < 0.001). 0.125 mg/kg of quinpirole caused a near significant increase in response latency, while 0.25 mg/kg (P = 0.003) and 0.5 mg/kg (P < 0.001) both achieved statistical significance. Quinpirole also had an effect on collection latency (F(3,18) = 10.875, P < 0.001), where the effects of the 0.125 mg/kg dose approached statistical significance (P = 0.06) and effects of 0.25 (P = 0.002) and 0.5 mg/kg (P < 0.001) reached statistical significance compared to vehicle (see Table 4 supplementary material).
Both the typical antipsychotics haloperidol and the atypical antipsychotic risperidone failed to antagonize an MK-801-induced impairment
The effects of MK-801 at a dose of 0.075 mg/kg largely replicated those observed in the previous study: MK-801 caused a highly significant decrease in percent correct (F(1,18) = 40.37, P < 0.001), as well as a decrease in trials completed (t(19) = 4.06, P < 0.001). However, haloperidol had no effect on the MK-801 induced accuracy impairment (F(3,20) = 0.417, P = 0.074). Moreover, haloperidol had no effect on MK-801-induced perseverations (Fig. 1, Table 2 supplementary material).
As in previous studies, MK-801 caused a significant decrease in percent correct (T(12) = 11.74, P < 0.001) and also resulted in a decrease in trials completed (T(17) = 5.78, P < 0.001) in the risperidone experiment. Risperidone had no effect on percent correct responses (F(3,21) = 0.894, P = 0.46) but caused a further decrease in trials completed at the highest dose tested (F(17) = 3.55, P = 0.002). Moreover, no effect of risperidone was seen on MK-801-induced perseverations (See Fig. 1, Table 2 supplementary material).
Both the haloperidol and risperidone attenuated an amphetamine-induced impairment
As before, amphetamine (0.5 mg/kg) caused a significant decrease in percent correct (t(10) = 4.152, P = 0.002). No effect of amphetamine was seen on any other measure. A Dunnett’s one-tailed t test indicated a near significant increase from the vehicle/amphetamine condition at 0.01 mg/kg (P = 0.058) and a significant increase at 0.02 (P = 0.018) mg/kg haloperidol when co-administered with amphetamine. No effect of haloperidol was seen on response (F(3,30) = 0.8, P = 0.50) or collection (F(3,30) = 1.90, P = 0.151) latencies (see Fig. 2, Table 3 supplementary material).
When tested in combination with risperidone, amphetamine (0.5 mg/kg) again caused a significant decrease in percent correct (T(11) = 3.994, P = 0.002). This was associated with a small but significant decrease in response latency (T(11) = 4.22, P = 0.001), while no significant effects were seen on any other measures (Fig. 2, Table 3 supplementary material).
When compared to the vehicle/amphetamine condition, risperidone had a significant effect on percent correct (F(3,33) = 6.728, P = 0.001). Post hoc analysis indicated that all conditions were associated with a significant improvement in percent correct (P < 0.002 for all conditions). Risperidone also altered collection latency (F(3,33) = 9.339, P < 0.001), with 2.0 mg/kg resulting in a longer latency when compared to the vehicle/amphetamine condition. Risperidone also influenced response latency (F(3,33) = 32.77, P < 0.001), where it caused a significant increase at all doses tested (0.5 mg/kg, P = 0.028, 0.1–2 mg/kg P < 0.001).
The D2 agonist quinpirole also attenuated an amphetamine-induced impairment
Again, amphetamine (0.5 mg/kg) caused a significant reduction in percent correct (T(11) = 3.217, P = 0.008). However, in this instance, a small but significant effect of amphetamine was also observed on response latency (T(11) = 3.385, P = 0.006; see Table 5) where animals responded slightly faster. At the highest dose tested, quinpirole caused a substantial decrease in trials completed (T(11. 5.518, P < 0.001). This treatment condition was therefore removed from further analysis, as its inclusion would have resulted in numerous empty cells, limiting the usefulness of the within-subject ANOVA. The lowest dose of quinpirole significantly reduced the amphetamine-induced deficit (0.031 mg/kg; P = 0.017), while no effect was observed at 0.063 mg/kg. While quinpirole did cause an increase in response latency, this did not reach statistical significance (F(2,20) = 2.417, P = 0.11). However, a significant effect of quinpirole was seen on collection latency (F(2,20) = 7.59, P = 0.003). Post hoc analysis indicated a significant increase at 0.063 mg/kg (P = 0.002), while 0.031 mg/kg just failed to reach statistical significance (P = 0.07).
The D1 receptor antagonists SCH-23390 also attenuated an amphetamine-induced impairment
Amphetamine (0.5 mg/kg) caused a significant decrease in percent correct when compared to the vehicle/vehicle condition (T(10) = 5.303, P < 0.001). In this instance, amphetamine had no effect on response and collection latency or trials completed. SCH-23390 significantly improved percent correct at all doses when compared to vehicle (0.005 mg/kg P = 0.005; 0.01 mg/kg P = 0.002; 0.02 mg/kg P < 0.001). Moreover, no effect of SCH-23390 was seen on response latency (F(3,30) = 1.604, P = 0.20) or collection latency (F(3,30) = 1.79, P = 0.17). No significant effect of SCH-23390 was observed on trials completed (Table 5).
Discussion
As previously reported, MK-801 and amphetamine both induced impairments in PAL (Ballard et al. 2013; Talpos et al. 2014). Furthermore, MK-801 had a profound effect on correction trial errors, disproportionately influencing performance on this trial type. While an effect was also observed following treatment with amphetamine on correction trial errors, it was smaller in magnitude (the amphetamine dose–response curve was previously presented in Talpos et al. (2014); however, the perseveration analysis was not included in the previous publication). Whether the effect observed with MK-801 represents true perseveration (bias towards responding at the previously correct location), a failure to attend to the stimuli, or a memory impairment associated with a specific trial type is unclear at this time. However, these impairment models appear dissociable on the basis of pharmacology and behavioral profiles.Footnote 1
Risperidone and haloperidol both reversed the amphetamine-induced deficit in percent correct. Remarkably, risperidone showed a near complete reversal over a dose range from 0.5–2.0 mg/kg. At doses of 1.0–2.0 mg/kg, but not at a lower dose of 0.5 mg/kg, this effect was associated with increases in latencies caused by risperidone. The effects of haloperidol were not as robust, but a clear beneficial effect was also observed. At 0.01 mg/kg, the effects of haloperidol to reverse an amphetamine-induced deficit just failed to reach statistical significance. However, statistical significance was achieved at the 0.02 mg/kg dose. Of note, 0.04 mg/kg did not result in additional improvements when compared to the 0.02 mg/kg dose, suggesting that haloperidol was not capable of completely reversing the amphetamine-induced impairments with the dosing regimen used here, as was the case with risperidone. As a whole, these data suggest that D2 receptor blockade reverses amphetamine-induced impairments in rPAL. However, it is possible that activity at other receptors may also contribute to the effects observed. In contrast, risperidone and haloperidol showed no benefit after an MK-801 challenge. If anything, they slightly exacerbated the effects of MK-801, showing a general trend towards a further decrease in percent correct and an increase in latency. This also made it necessary to use lower doses of risperidone and haloperidol, as higher doses, in combination with MK-801, resulted in large reductions in responding.
Amphetamine has a diverse pharmacology and is known to increase free levels of dopamine at the synaptic cleft, likely via its ability to disrupt function at the plasma membrane and vesicular monoamine transporters (Robertson et al. 2009; Sulzer et al. 2005). Since the effects of amphetamine could be modulated by haloperidol and risperidone, presumably via actions at the D2 receptor, we performed additional studies to further explore the relative contributions of D1 versus D2 receptors to the effects observed here. When tested alone, the D1 agonist SKF-82958 caused a near significant decrease in accuracy, without influencing secondary measures, suggesting that at higher doses, a statistically significant impairment could be detected. Similarly, the D2 agonist quinpirole also caused a significant impairment in PAL when tested alone. However, this effect was only seen with a concomitant decrease in trials completed and an increase in response latency. These results suggest that stimulation of both the D1 and the D2 receptors could contribute to the amphetamine-induced impairment observed in PAL, although D2 agonism may be more associated with non-cognitive effects, as evidenced by effects on responsivity. Furthermore, the finding that both selective D1 or D2 agonists can disrupt performance in PAL opens the possibility that amphetamine may also disrupt PAL performance via unique D1 and D2 receptor-mediated pathways. To further elucidate the importance of D1 and D2 receptors in the amphetamine impairment PAL model, we also tried to reverse the amphetamine-induced impairment with the D1 receptor antagonist SCH-23390. SCH-23390 potently reversed the effects of amphetamine on accuracy at all doses tested. These findings provide further support to the idea that both D1 and D2 receptor activation is involved in the cognitive deficit seen in the PAL amphetamine model. This is an exciting possibility as it would suggest that over activation of the excitatory D1 receptors or the inhibitory D2 receptors could both result in cognitive impairment in rPAL. If so, amphetamine may in fact represent two distinct paths to cognitive impairment. However, it should be noted that owing to the long training time associated with the PAL task, it was necessary to reuse animals. Accordingly, behavioral responses to pharmacological treatments may have been altered as a result of previous drug exposure. External replication of these studies would help to confirm the results reported here.
Counter-intuitively, the D2 receptor agonist quinpirole was also able to reverse the amphetamine-induced deficit when tested at very low doses. This finding is possibly due to a preferential action of quinpirole on pre-synaptic inhibitory D2 auto-receptors. A selective stimulation of these receptors would be expected to decrease dopamine release, potentially improving accuracy as was observed here. In line with this suggestion, the beneficial effects of quinpirole decreased when tested at higher doses, presumably because postsynaptic D2 receptors become increasingly activated—although this remains speculative in the absence of concomitant receptor occupancy data.
It has been previously shown that PAL performance is impaired after direct infusion of MK-801 or CNQX into the hippocampus (Talpos et al. 2009). Accordingly, it seems likely that some of the effects of systemic administration of MK-801 are being driven by the hippocampus. Besides disrupting LTP within the hippocampus (Herron et al. 1986), NMDA receptor antagonists have also been shown to disrupt EEG gamma signals (for a recent review, see Hunt and Kasicki 2013), likely indicating a disconnection between the hippocampus and pre-frontal cortex. Accordingly, the MK-801 impairment model may also be relevant for other disorders where hippocampal-pre-frontal disconnectivity may play a role, such as in Alzheimer’s disease.
The data presented here shows that PAL is sensitive to alterations of dopaminergic function, whereas a visual discrimination task is relatively resistant to an amphetamine challenge (Talpos et al. 2012). We speculate that amphetamine induces an impairment in this task by increasing dopamine levels within the orbital pre-frontal cortex (OFC). OFC dopamine levels have been shown to be critical for numerous aspects of executive function, such as working memory, attentional selection when confronted with a distracter, and maintaining a stimuli representation, as discussed in Roberts et al. (2007). rPAL is unlikely to require working memory per se. However, if it is to be solved using an object-in-place rule, which may require the flexible, simultaneous manipulation of a spatial and a visual representation in order to successfully complete the task, then it would have much in common with the utilization of working memory. While purely speculative, this may also explain why the task remains sensitive to manipulations of the hippocampus after it has been acquired (Talpos et al. 2009, 2014).
While some studies have claimed an improvement in cognitive abilities as a result of antipsychotic treatment in schizophrenia patients (Hagan and Jones 2005), these effects are generally modest and may be confounded by side effects of said treatment. The fact that risperidone and haloperidol can dramatically improve performance on rPAL might be taken as evidence that the amphetamine model lacks predictive validity for a clinical setting. Yet, considering amphetamines primary action is to increase dopamine levels, it would be of serious concern if this effect could not be reversed by a dopamine antagonist. Accordingly, we feel that the impairment induced by amphetamine in PAL may still be of benefit for modeling behavioral abnormalities associated with schizophrenia, especially the potential overlap between positive symptoms and cognitive impairment. Moreover the finding that both the D1 and D2 receptor may be involved in the amphetamine impairment model suggests that rPAL may be of use in untangling the relationship between cognitive symptoms, positive symptoms, and the influence of the D2 antipsychotics on cognition. Unfortunately, hyper-dopaminergic models, whether with amphetamine or a D1 receptor agonists, will be at risk of detecting mechanistic false-positive effects (D2 antagonists). Regardless, rPAL does appear to be exquisitely sensitive to changes within the dopaminergic system. Many models of amphetamine psychosis take advantage of the compounds action as a psychostimulant. For example, in tests of locomotor activity, amphetamine induces a hyperlocomotion that can then be reversed with a wide variety of drugs. However, a shortcoming of this approach is that any drug that non-specifically reduces the expression of behavior (e.g., causes sickness or sedation) will have an “enhancing” profile. Yet, this is not the case in PAL, where such gross changes would be expected to also cause a disruption in PAL. This may make rPAL of specific interest for the broader study of dopaminergic function rather than being utilized solely as a translational model of disease.
Here, we have demonstrated that rPAL is sensitive to two different, but dissociable, pharmacological models of schizophrenia. While amphetamine is generally used to model positive symptoms, and NMDA antagonists cognitive symptoms of schizophrenia, both challenge types cause impairments in rPAL. rPAL is a novel approach to model the impact of glutamatergic or dopaminergic dysfunction on cognition and may also affording a unique means to study the interplay between positive symptoms, dopamine, and cognitive impairment in the development of novel treatments for schizophrenia.
Notes
The amphetamine impairment model has proven to be very consistent. We have employed this challenge in over 20 studies and have only once observed a lack of an amphetamine effect. In contrast, the MK-801 model has been more difficult to employ. 0.05 mg/kg will cause deficits in percent correct with no effects on secondary measures; however, this dose may lack the consistency needed for repeated drug screening. In contrast at 0.075 mg/kg MK-801 did consistently cause a decrease in percent correct, but this same dose will also cause substantial increases in response latencies. Since response latency is preferentially affected by MK-801 over collection latency, this may represent cognitive slowing as opposed to non-specific behavioral changes. However, we cannot rule out the possibility that some of the effects observed on percent correct after an MK-801 challenge are non-cognitive in nature.
References
Abi-Dargham A (2004) Do we still believe in the dopamine hypothesis? New data bring new evidence. Int J Neuropsychopharmacol 7(Suppl 1):S1–S5
Angrist B, Sathananthan G, Wilk S, Gershon S (1974) Amphetamine psychosis: behavioral and biochemical aspects. J Psychiatr Res 11:13–23
Ballard TM, Wyler R, Burns T (2013) Comparison of scopolamine and MK801 effects on touchscreen based cognitive tasks in rats. The annual meeting of the Society for Neuroscience
Barnett JH, Sahakian BJ, Werners U, Hill KE, Brazil R, Gallagher O, Bullmore ET, Jones PB (2005) Visuospatial learning and executive function are independently impaired in first-episode psychosis. Psychol Med 7:1031–1041
Barnett JH, Robbins TW, Leeson VC, Sahakian BJ, Joyce EM, Blackwell AD (2010) Assessing cognitive function in clinical trials of schizophrenia. Neurosci Biobehav Rev 34:1161–1177
Blackwell AD, Sahakian BJ, Vesey R, Sample JM, Robbins TW, Hodges JR (2004) Detecting dementia: novel neuropsychological markers of preclinical Alzheimer's disease. Dement Geriatr Cogn Disord 17:42–48
Brisch R, Saniotis A, Wolf R, Bielau H, Bernstein HG, Steiner J, Bogerts B, Braun AK, Jankowski Z, Kumaritlake J, Henneberg M, Gos T (2014) The role of dopamine in schizophrenia from a neurobiological and evolutionary perspective: old fashioned, but still in vogue. Front Psychiatry 5:47
Bussey TJ, Barch DM, Baxter MG (2013) Testing long-term memory in animal models of schizophrenia: suggestions from CNTRICS. Neurosci Biobehav Rev 37:2141–2148
Carlsson A (1988) The current status of the dopamine hypothesis of schizophrenia. Neuropsychopharmacology 1:179–186
Gilmour G, Dix S, Fellini L, Gastambide F, Plath N, Steckler T, Talpos J, Tricklebank M (2012) NMDA receptors, cognition and schizophrenia—testing the validity of the NMDA receptor hypofunction hypothesis. Neuropharmacology 62:1401–1412
Hagan JJ, Jones DN (2005) Predicting drug efficacy for cognitive deficits in schizophrenia. Schizophr Bull 31:830–853
Herron CE, Lester RA, Coan EJ, Collingridge GL (1986) Frequency-dependent involvement of NMDA receptors in the hippocampus: a novel synaptic mechanism. Nature 322:265–268
Hunt MJ, Kasicki S (2013) A systematic review of the effects of NMDA receptor antagonists on oscillatory activity recorded in vivo. J Psychopharmacol 27:972–986
Krystal JH, D’Souza DC, Mathalon D, Perry E, Belger A, Hoffman R (2003) NMDA receptor antagonist effects, cortical glutamatergic function, and schizophrenia: toward a paradigm shift in medication development. Psychopharmacology (Berl) 169:215–233
Lau CI, Wang HC, Hsu JL, Liu ME (2013) Does the dopamine hypothesis explain schizophrenia? Rev Neurosci 24:389–400
Markou A, Chiamulera C, Geyer MA, Tricklebank M, Steckler T (2009) Removing obstacles in neuroscience drug discovery: the future path for animal models. Neuropsychopharmacology 34:74–89
Nithianantharajah J, Komiyama NH, McKechanie A, Johnstone M, Blackwood DH, St CD, Emes RD, van de Lagemaat LN, Saksida LM, Bussey TJ, Grant SG (2013) Synaptic scaffold evolution generated components of vertebrate cognitive complexity. Nat Neurosci 16:16–24
Roberts AC, Reekie Y, Braesicke K (2007) Synergistic and regulatory effects of orbitofrontal cortex on amygdala-dependent appetitive behavior. Ann N Y Acad Sci 1121:297–319
Robertson SD, Matthies HJ, Galli A (2009) A closer look at amphetamine-induced reverse transport and trafficking of the dopamine and norepinephrine transporters. Mol Neurobiol 39:73–80
Sulzer D, Sonders MS, Galli A (2005) Mechanisms of neurotransmitter release my amphetamines: a review. Prog Neurobiol 75:406–433
Talpos J, Steckler T (2013) Touching on translation. Cell Tissue Res 354:297–308
Talpos JC, Winters BD, Dias R, Saksida LM, Bussey TJ (2009) A novel touchscreen-automated paired-associate learning (PAL) task sensitive to pharmacological manipulation of the hippocampus: a translational rodent model of cognitive impairments in neurodegenerative disease. Psychopharmacology (Berl) 205:157–168
Talpos JC, Fletcher AC, Circelli C, Tricklebank MD, Dix SL (2012) The pharmacological sensitivity of a touchscreen-based visual discrimination task in the rat using simple and perceptually challenging stimuli. Psychopharmacology (Berl) 221:437–449
Talpos JC, Aerts N, Fellini L, Steckler T (2014) A touch-screen based paired-associates learning (PAL) task for the rat may provide a translatable pharmacological model of human cognitive impairment. Pharmacol Biochem Behav 122:97–106
Tamminga CA (1998) Schizophrenia and glutamatergic transmission. Crit Rev Neurobiol 12:21–36
Acknowledgments
NEWMEDS—the research leading to these results has received support from the Innovative Medicines Initiative Joint Undertaking under Grant agreement no. 115008 of which resources are composed of EFPIA in-kind contribution and financial contribution from the European Union’s Seventh Framework Programme (FP7/2007-2013).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Talpos, J., Aerts, N., Waddell, J. et al. MK-801 and amphetamine result in dissociable profiles of cognitive impairment in a rodent paired associates learning task with relevance for schizophrenia. Psychopharmacology 232, 3911–3920 (2015). https://doi.org/10.1007/s00213-015-3934-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-015-3934-x