
Abstract
Cardiomyocyte infarction could lead to high morbidity and mortality worldwide. Recent studies demonstrated that Heme oxygenase-1 (HO-1) could exert cardiac protective effect and arouse attention. However, the detailed mechanism is still unclear. Our study provided evidences of the protective effect of HO-1 overexpression on cardiomyocytes against hypoxia/reoxygenation (H/R). We divided the treatment into four groups: the control group, H/R group, H/R+HO-1 group, and H/R+Null group. Immunofluorescent study was utilized to label the BrdU-positive and LC3-positive cells. Flow cytometry and TUNEL assay were used to examine the cell apoptosis. Protein levels of Bax, Bcl-2, Sirt3, beclin-1, LC3-I, and LC3-II were both measured using western blotting. The results indicated that HO-1 overexpression decreased the cell apoptosis and enhanced the cell proliferation. The level of Sirt3 and autophagy were also increased in H/R+HO-1 group compared with H/R group. However, ZnPP, a HO-1 inhibitor, and SiRNA of Sirt3 are both reversed the decrease of cell apoptosis of HO-1 overexpression. Moreover, ZnPP also decreased the expression of Sirt3 in HO-1 overexpression treatment group. In summary, HO-1 overexpression protects cardiomyocytes against H/R injury via ameliorating cell apoptosis and enhancing cell proliferation and autophagy through Sirt3 signaling pathway.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Myocardial hypoxia/reoxygenation (H/R) injury is one of the main causes of death and disability worldwide (Murphy and Steenbergen 2008). Reperfusion, a rapid recovery of blood flow and oxygen supply, is a protective step of the progression of damage during myocardial ischemia. However, reperfusion-induced injury may lead to irreversible damage to the myocardium, which ultimately causes myocardial cell apoptosis (Fujimoto et al. 2004; Hausenloy and Yellon 2013). However, few therapies are available to cure this injury. Thus, it is imperative to figure out the mechanism underlying myocardial ischemia/reperfusion injury.
Studies have focused on calcium overload, reactive oxygen species (ROS), endoplasmic reticulum stress, ATP depletion, and activation of autophagy and apoptosis (Wei et al. 2016; Du et al. 2017). Among above, the autophagy and apoptosis of myocardial cell during ischemia/reperfusion injury are the main causes. Apoptosis, a major form of cell death, is associated with removing excess, damaged or infected cells throughout life. It is either an essential component of normal development or a response to pathological disease states (Bonavita et al. 2003). Apoptosis in the heart is a major regulator in the process of cardiac failure due to myocardial ischemia/reperfusion (Haunstetter and Izumo 1998). Autophagy could ameliorate mitochondrial dysfunction via autophagosome forming, which is a catabolic process to preserve the mitochondria’s structural and functional integrity (Chen et al. 2016). The sirtuin3 (Sirt3), which belong to the family of NAD+-dependent deacetylase proteins, is a crucial regulator of mitochondrial function controls global acetylation of the organelle. It is a major acetylation enzyme located in mammalian mitochondria. Sirt3 signaling pathway is partially involved in anti-apoptosis effect to exert cardioprotective effect (Du et al. 2017). Moreover, downregulation of Sirt3 could partially prevented the autophagy in endoplasmic reticulum stress–associated neuronal injury conditions (Yan et al. 2018).
Heme oxygenase-1 (HO-1) belongs to the low molecular weight heat shock protein (HSP) family. A series studies reported that induction and expression of HO-1 exerts protective mechanisms in ischemia/reperfusion injury, especially in cardiomyocyte ischemia/reperfusion injury (Maines 1997; Melo et al. 2002; Vulapalli et al. 2002; Paine et al. 2010). HO-1 induction in donor organs in rats has also been revealed to ease ischemia/reperfusion-induced injury, prolong graft survival and improve the long-term function of grafted kidney (Tullius et al. 2002). Studies have demonstrated that HO-1 could exert protective effect through anti-oxidative mechanism, the maintenance of microcirculation, anti-inflammation, anti-arrhythmia, and anti-apoptosis mechanisms (Chen et al. 2016). However, the mechanism underlying HO-1 protective effect against H/R injury in cardiomyocytes is still uncertain.
Thus, the aim of this study was to investigate the possible underlying mechanisms of HO-1 in protecting against myocardial ischemia/reoxygenation injury. In our study, we hypothesize that anti-apoptosis, autophagy, and proliferation were involved in the protective effect.
Materials and methods
Primary cell culture
The H9c2 cardiomyocyte cell line was obtained from neonatal rat cardiomyocytes using the American Type Culture Collection (ATCC, Manassas, VA, USA). The H9c2 cells were cultured in high-glucose DMEM solution with 10% v/v fetal bovine serum (FBS) at 37 °C in a humidified atmosphere containing 5% carbon dioxide. The medium was changed every 2 days, and the cells were subcultured or subjected to experimental procedure when the cell population density reached 70–80% confluence. In all experiments, the cells were treated with serum starvation for 24 h before treatment.
Stimulated ischemia/reperfusion
For the ischemia/reperfusion experiments, the hypoxic model was performed by incubating the cells in the hypoxic chamber equilibrated with 92.5% N2, 5% CO2, and 2.5% O2 at 37 °C for 24 h. Then, the hypoxic cells were recovered at normal culture condition in a humidified atmosphere (5% CO2/95% air) for 18 h. The experiment was divided into four groups: control group, H/R group, H/R+HO-1 group, and H/R+Null group. Cells in H/R/HO-1 group were characterized with HO-1 overexpression and H/R + Null group received the same treatment with control lentivirus. In the ZnPP and SiRNA against Sirt3 experiments, the cells were pretreated for 6 h with the HO-1 inhibitor ZnPP (10 μM, Sigma) and Si-Sirt3 then subjected to hypoxia and reoxygenation.
Lv-HO1 H9c2 cell line construction
Lv-HO1 H9c2 cell line was constructed according to previous study (Chen et al. 2016). Briefly, cells were transfected with Ad-mCherry-LC3-GFP lentivirus (MOI = 50). The transfected H9c2 cells express GFP which can be measured through fluorescence microscope.
Confocal microscopy measurement
The overexpression of HO-1 was determined using confocal microscopy. Cells transfected with Ad-mCherry-LC3-GFP lentivirus were fixed with 4% PFA (paraformaldehyde). After that, the cells were washed with PBS for three times. For detection of autophagy, cells were fixed with 4% PFA. Then, cells were blocked with 5% goat serum in 0.25% Triton-X PBS and incubated with LC3 primary antibody (Abcam, ab48394, 1 μg/ml), followed by incubation of secondary antibody (1: 500). Finally, cells were stained with DAPI to visualize the nuclei. For BrdU incorporation assay, the cells were cultured in 0.1% gelatin-coated glass slides in 12-well plates for 12 h, and then cells were grown in 30 μM BrdU for 2 h, fixed by 3% formaldehyde in PBS. After washing with PBS three times, cells were acid-denatured with 2 M HCl in PBS for 30 min and further blocked by 2% FBS/PBS followed by a primary anti-BrdU antibody incubation for 30 min. Cells were then washed with PBS and incubated with second antibody for 30 min. Finally, cells were counterstained DAPI. Finally, antifade mounting medium was added to all cells and the cells were imaged under a confocal microscope (LSM 780, Carl Zeiss, Jena, Germany).
Western blot assay
The H9c2 cells of four groups were collected. Proteins were extracted from the cells based on previous studies. Briefly, cells were homogenized in lysis buffer with 1% protease inhibitor cocktail. The lysates were centrifuged for 15 min at 12,000 rpm. The concentration of each protein sample was determined using the Bradford Protein Assay Kit (Beyotime Biotechnology, China). Equal amounts of protein sample were subjected to 8–12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), electrotransferred onto polyvinylidene difluoride (PVDF) membranes. After blocking with 5% skimmed milk, the membranes were incubated with primary antibodies for HO-1 (1:1000), Sirt-3 (1:1000), Beclin-1 (1:800), LC3A/B (1:1000), Bax (1:1000), Bcl-2 (1:1000), and β-actin (1:2000, Cell Signaling Technology, USA) overnight at 4 °C. After washing with TBST, the membranes were incubated with secondary antibody (1:10,000) for 2 h. After washing with TBST for three times, protein bands were detected using a chemiluminescence imaging analysis system. The intensity of bands was analyzed using ImageJ, and the results were normalized to the levels of β-actin.
Detection of the rate of apoptosis
Apoptosis was detected using an Annexin-V FITC/PI apoptosis kit according to the manufacturers’s direction (KeyGEN, Nanjing, China). After treatment, the cells (1~5 × 105) were collected and placed in EP tubes. Then, the cells were rinsed with 0.1 mmol/L PBS (PH = 7.4) and digested with 0.25% trypsin EDTA-free. After that, the cells were resuspended in 500 μL of binding buffer, incubated with 5 μL of Annexin-V-FITC in a dark room for 10 min. Then, 5 μL of propidium iodide (PI) was added and kept at room temperature without light for 15 min. Cellular fluorescence were measured by flow cytometer (FACSCalibur; BD Biosciences, Franklin Lakes, NJ, USA).
Cell viability
Cells were cultured in 96-well plates, and cell viability was measured by cell-counting kit-8 (CCK-8, Beyotime). Ten microliters of CCK-8 solution was added to each well, and the plates were incubated for 3 h. The viability of cells was measured at 450 nm using an ELISA reader (Thermo Fisher). The mean optical density (OD) of five wells was used to obtain the percentage of cell viability. The independent experiments were performed three times.
TUNEL assay
Cultured cells were stained with terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling (TUNEL) assay to determine the myocardial apoptosis. TUNEL staining was performed under manufacturer indication by In Situ Cell Death Detection Kit (Roche Diagnostics, Indianapolis, USA). The percentage of TUNEL-positive cells was calculated as the number of TUNEL-positive nuclei divided by the total number of nuclei stained with DAPI.
Statistical analysis
All data were represented as mean ± SEM. GraphPad Prism was used to measure the statistical analysis. Comparison between various groups was analyzed by one-way ANOVA followed by the Student–Newman–Keuls test. The difference was set at p < 0.05.
Results
HO-1 overexpression in H9c2 cells
HO-1 overexpression was constructed through an Lv-HO-1-H9c2 cell line encoding lentivirus. Cells with overexpression of HO-1 showed increasing green fluorescent measured by fluorescent microscopy compared with LV-Null group as shown in our Fig. 1a, b. In consistent with the confocal result, the expression of HO-1 in the Lv-HO-1 group cells was significantly higher than the control group (p < 0.05); however, the Lv-Null group had no change on HO-1 expression (p > 0.05, Fig. 1c, d). These results showed that our method to overexpress HO-1 is effective.

The overexpression of HO-1 in H9c2 cells. a, b Immunohistochemistry staining showed the overexpression of HO-1 in the Lv-HO-1 group and the Lv-Null group in H9c2 cells. c Immunoblot band of HO-1 expression using western blotting method in each groups. c Statistical analysis of HO-1 expression and the result were normalized to the control group (n = 3 wells per group, five fields were randomly selected to obtain the mean intensity in each well). Data were presented as the mean ± SEM. *P < 0.05 relative to the control group
HO-1 overexpression protects H9c2 cells from apoptosis with H/R model
To characterize the effect of HO-1 overexpression following myocardial infarction, a hypoxia/ischemia model was utilized. Research has reported that H/I could induce cell apoptosis in cardiomyocytes (Duan et al. 2016). We also measured the effect of HO-1 on apoptosis in H9c2 cells with H/R model using Annxin V/PI staining. As depicted in Fig. 2a–d, cells in the control group were presented in a round shape with full and integral appearance. H/R injury resulted in cell shrinkage in morphology. However, HO-1 overexpression prevented the morphology change from H/R injury in H9c2 cells, whereas the H/R+Null group had no protective effect. H/R model remarkably increased the apoptosis levels compared to the control group (p < 0.001), which is in consistent with previous studies. Of note, HO-1 overexpression reduced the apoptosis levels which were induced by H/R (p < 0.01). However, H/R+Null group cells showed approximately three times apoptosis than the control group (p < 0.01) and had no protective effect on apoptosis (Fig. 2i). The effect of HO-1 overexpression was confirmed using flow cytometry. H/R injury induced marked apoptosis compared with the control group. HO-1 overexpression ameliorated apoptosis induced by H/R, while H/R+Null group had no effect compare to the H/R group (Fig. 2e, h). Indeed, HO-1 overexpression showed neuronal protective effect against H/R in H9c2 cells.

Effect of HO-1 overexpression on cell survival in H9c2 cells during H/R. a–d Microscope images for nuclei from H9c2 cells. Apoptotic bodies were presented in H/R and H/R+Lv-Null groups. By contrast, they were significantly decreased in HO-1+Lv-H/R group. e–h Representative flow cytometry results were showed in four groups. i Relative apoptosis levels were analyzed from flow cytometry results (n = 5 in each group, experiments were repeated three times to obtain an average value). Data were presented as the mean ± SEM. ***P < 0.001 and **P < 0.01 relative to the control group. ##P < 0.01 relative to the H/R group
HO-1 overexpression increased autophagy in H9c2 cells with H/R injury
Emerging evidence suggests that autophagy plays a protective role in heart disease (Wang et al. 2015b). Research has been reported that HO-1 possesses anti-apoptotic properties through enhancing autophagy in podocytes (Dong et al. 2015). Light chain 3 (LC3) is a specific marker for autolysosomes; thus, we first labeled LC3 and DAPI to find out the co-localization in H9c2 cells. LC3 was scattered in both cytosolic and nuclear in four groups, whereas the H/R+LV-HO-1 groups showed highest expression of LC3 signals (Fig. 3a, d). We then measured the protein levels of beclin-1, LC3-I, and LC3-II by western blotting assay. Our results showed that the H/R did not change the expression of beclin-1, LC-I, and LC-II (p > 0.05). However, HO-1 overexpression upregulated the protein levels of beclin-1, LC-I, and LC-II (p < 0.01, Fig. 3e–g). These results indicated that HO-1 overexpression in H9c2 cells could enhance the autolysosome formation which is mediated by beclin-1 during H/R injury.

HO-1 overexpression enhanced the H9c2 cells autophagy during H/R injury. a–d Double-staining images of LC3 (red) and DAPI (blue) were presented in different groups. e Western blotting immunoblot bands of beclin-1, LC3-I, and LC3-II were displayed. f, g Quantitative analysis of beclin-1 and LC3-II/LC3-I were shown. Beclin-1 was presented as normalized to β-actin (n = 5 in each group, experiments were repeated three times to obtain an average value). Data were presented as the mean ± SEM. #P < 0.05 and ##P < 0.01 relative to the H/R group
HO-1 overexpression enhanced proliferation, cell viability, and anti-apoptosis in H9c2 cells during H/R injury
The effects of HO-1 overexpression on cell proliferation were detected by a BrdU assay. We measured the co-localization of BrdU and DAPI, and cells with BrdU-positive signal were considered in proliferation. We also measured the BrdU-positive cell ratio in four groups. H/R injury remarkably reduced the BrdU-positive cells compared with the control group (p < 0.001). HO-1 overexpression significantly enhanced the BrdU-positive cell ratio compared with both H/R group and H/R+LV-Null group (p < 0.01. Fig 4a–e). In order to investigate whether HO-1 overexpression could exert protective effect, we measured the cell viability. In our result, compared to the control group, H/R markedly decreased the relative cell viability (p < 0.01). However, H/R+HO-1 significantly increased the cell viability compared with the H/R group and H/R+Null group (p < 0.01, Fig. 4f). Protein levels of Bax and Bcl-1 were examined. As expected, H/R upregulated the Bax expression (p < 0.001) and downregulated the Bcl-2 expression (p < 0.001). HO-1 overexpression prevented the increase of Bax level (p < 0.01) and decrease of Bcl-2 level during H/R (p < 0.001), whereas H/R+Null group had no effect (p > 0.05, Fig. 4g, h). These results implied that the protective effect of HO-1 on H9c2 cells was via enhancing proliferation, cell viability, as well as anti-apoptosis.

Effect of HO-1 overexpression on cell proliferation, cell viability, and apoptosis. a–d Immunofluorescence images of proliferating H9c2 cells with BrdU (red) and DAPI (blue) staining in four groups. e BrdU-positive cell ratio was quantitated in different groups from a–d. f Cell viability was determined with CCK-8 kit in four groups, and the relative cell viability was quantitated. g Western blotting assay was performed to measure Bax and Bcl-2 expression and representative images were presented. h Quantization for Bax and Bcl-2 were normalized to β-actin. Data were presented as the mean ± SEM. ***P < 0.001 relative to the control group (n = 5 in each group, experiments were repeated three times to obtain an average value). **P < 0.01 and ***P < 0.001 relative to the control group. ##P < 0.01 relative to the H/R group
HO-1 overexpression protects H9c2 cells from stress-mediated cell death with H/R model
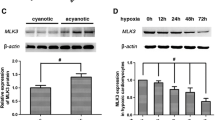
It has been reported that Sirt3 signaling pathway is involved in H/R-induced cardiomyocyte apoptosis (Du et al. 2017). In order to elucidate the mechanism underlying the anti-apoptosis of HO-1 in H9c2 cells, we measured the Sirt3 protein levels. In our study, H/R significantly reduced the expression of Sirt3 in H9c2 cells compared to control group (p < 0.05). HO-1 overexpression potently prevented the decrease of Sirt3 induced by H/R (p < 0.01), while Null treatment group showed remarkable decrease of Sirt3 compared with HO-1 group (p < 0.05, Fig. 5).

The effect of HO-1 overexpression on Sirt3 expression in H9c2 cells during H/R injury. a Representative immunoblot band of Sirt3 expression in different groups. b Quantitative result of Sirt3 levels was analyzed from a, and the results were normalized to the control group (n = 5 in each group). Data were presented as the mean ± SEM. *P < 0.05 relative to the control group. ##P < 0.01 relative to the H/R group (n = 5 in each group, experiments were repeated three times to obtain an average value)
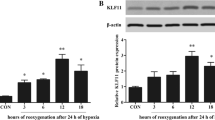
HO-1 inhibitor ZnPP reversed the increase of Sirt3 in H9c2 cells with H/R injury
Since overexpression of HO-1 could increase the expression of Sitr3 in H9c2 cells with H/R injury, we then evaluated whether inhibit the overexpression of HO-1 could eliminate the protective effect. TUNEL assay demonstrated that the H/R+Lv-HO-1+ZnPP group possessed higher amount of TUNEL-positive cells compared with the H/R+Lv-HO-1 group (p < 0.05, Fig. 6a, b, e). In accordance with the TUNEL result, the expression of cleaved caspase-3 was increased after ZnPP treatment compared with the H/R+Lv-HO-1 group (p < 0.05, Fig. 6g), indicating ZnPP could abolish the anti-apoptotic effect of HO-1 overexpression. To further validate the underlying mechanism of overexpression of HO-1 in H/R injury is associated with Sirt3 signaling, we used a Si-RNA of Sirt3 to downregulate the expression of Sirt3. Our data presented that TUNEL-positive cells were higher in Si-Sirt3 treatment group compared with the SiRNA control group. Moreover, the level of cleaved caspase-3 was also increased after Si-Sirt3 treatment with overexpression of HO-1 in H/R-injured H9c2 cells (p < 0.05, Fig. 6c–e). HO-1 inhibitor ZnPP significantly decreased the overexpression of HO-1; interestingly, the level of Sirt3 was also decreased. Si-sirt3 downregulated the expression Sirt3; however, the expression of HO-1 had no change after Si-Sirt3 treatment (Fig. 6f). These results indicated that Sirt3 signaling is involved in the protective effect of HO-1 overexpression in H/R-injured H9c2 cells.

Effects of HO-1 inhibitor ZnPP and SiRNA against Sirt3 on apoptosis in H9c2 cell with H/R model. a–d TUNEL staining representative images in different groups. e TUNEL-positive cells were analyzed from a. ZnPP and Si-Sirt3 both increased the TUNEL-positive cells in H9c2 cells with H/R+Lv-HO-1. f The expression level of HO-1 and Sirt3 were both decreased after the use of ZnPP. The level of Sirt3 was decreased after the application of Si-Sirt3 and the HO-1 expression had no change. g The expression of cleaved caspase3 was increased in both ZnPP and Si-Sirt3 treatment, respectively (n = 5 in each group, experiments were repeated three times to obtain an average value). Data were presented as the mean ± SEM. a P < 0.05
Discussion
In the present study, we revealed the protective effect of HO-1 overexpression on H/R-injured H9c2 cells. We observed that HO-1 overexpression could remarkably decrease cell apoptosis and enhance cell viability in H/R-injured H9c2 cells. This cardioprotective effect of HO-1 overexpression was mediated by anti-apoptosis through Sirt3 signaling pathway and cell autophagy. Inhibition of HO-1 and downregulation of Sirt3 could both reverse the protective effect of HO-1 overexpression.
Previous study has reported that HO-1 overexpression could protect the stability of the mitochondrial membrane and reduce mitochondrial oxidation products (Chen et al. 2016). Our study was inspired by the research and further investigated the mechanism underlying the protective effect of HO-1. Hypoxia/ischemia in heart disease arouses much attention due to its high mortality worldwide (Mortality and Causes of Death 2015). Following ischemia, reperfusion is the secondary period of myocyte injury including oxidative stress, necrosis, and apoptosis (Wang et al. 2015a). Apoptosis is considered to be an important factor in the cell death of cardiomyocytes (Oerlemans et al. 2013; Wei et al. 2018); thus, it is crucial to find an appropriate agent to exert anti-apoptotic effect. A line of studies has reported that pharmacological ways to upregulate HO-1 expression could exert anti-apoptotic effects (Shi et al. 2016; Tang et al. 2018a). HO-1 also exerts cardioprotection by producing biliverdin/bilirubin (Vulapalli et al. 2002). Moreover, HO-1 could protect cardiomyocytes against hypoxia/reperfusion-induced apoptosis through enhancing the Akt signaling pathway, which further inhibited the JNK/c-Jun/caspase-3 signaling pathway (Li et al. 2016a). We first constructed the HO-1 overexpression cell line, which further performed H/R injury. Indeed, our result manifested that the HO-1 overexpression could protect H9c2 myocardia cells against cell apoptosis. This result is in accordance with previous studies (He et al. 2018). HO-1 is a downstream protein from Nrf2, which is response to oxidative/electrophilic stress, and upregulation of Nrf2 could enhance the level of HO-1 (Shelton et al. 2013). Haijie Yu and colleagues have demonstrated that pharmacological way to enhance Nrf2/HO-1 pathway could exert cardioprotective effects (Yu et al. 2016). That is also provided evidence that overexpression of HO-1 could protect cardiomyocytes against H/R injury through anti-apoptotic effect. Our results showed the overexpression of HO-1 decreased the cell apoptosis, and HO-1 inhibitor ZnPP reversed the effect of HO-overexpression. Furthermore, the levels of anti-apoptotic protein Bax and pro-apoptotic protein Bcl-2 were examined to confirm the anti-apoptotic role of HO-1 during H/R (Tang et al. 2018b). As expected, HO-1 overexpression downregulated the Bax levels and upregulated the Bcl-1 levels during H/R injury. These data supported the beneficial effect on cardiomyocytes during the H/R injury.
Sirt3 is a histone deacetylase which is localized primarily in mitochondria and becomes activated by the proteolytic processing at N-terminus (Michishita et al. 2005). Sirt3 is essential for the integration of cellular energy metabolism as well as ROS generation (Kong et al. 2010). A line of studies reported that Sirt3 signaling pathway could protect cardiomyocytes from apoptosis as well as mitochondrial DNA (mtDNA) damage (Pillai et al. 2016; Du et al. 2017). Thus, we hypotheses that HO-1 showed anti-apoptotic effect through Sirt3 signaling pathway. Our result demonstrated that HO-1 reversed the decrease of Sirt3 levels induced by H/R injury in H9c2 cells, and ZnPP could downregulate the expression of Sirt3. Indeed, the Sit3 signaling pathway is involved in the protective effect of HO-1 overexpression. Deficiency of Sirt3 is related to increasing mtDNA damage, which is closely associated with apoptosis. Our results were consistent with the previous studies that downregulation of Sirt3 with SiRNA enhanced the apoptosis level. However, overexpression of Sirt3 alleviated mtDNA damage and showed mtDNA damage repair (Du et al. 2017).
Autophagy is a dynamic process involving degradation and recycling of cellular components in the lysosomes (Klionsky and Emr 2000). It plays an imperative role in transport protein aggregates and damaged or superfluous organelles, which further to maintain cell healthy (Gustafsson and Gottlieb 2009). Growing attention has been paid on the process dysfunction of cardiomyocytes in the study of ischemia/reperfusion injury. Under normal conditions, autophagy is at low levels, and decrease in this process leads to cardiac dysfunction and heart failure (Gottlieb and Mentzer 2010). The levels of autophagy are important in its protective effect. Sirt3 also plays role in inducing autophagy through AMPK-mTOR pathway (Dai et al. 2017) and the deacetylation of FoxO1 (Li et al. 2016b). Autophagy activation could reduce the myocardial infarct size, whereas excessive autophagy could result in cell death (Matsui et al. 2007). HO-1 could protect cardiomyocytes against hypoxia/reoxygenation through induction of autophagy (Chen et al. 2016). Our studies showed that normal condition and H/R injury had no effect on the autophagy level. However, HO-1 overexpression could enhance the cell autophagy, which is a protective process in cardiomyocytes. Studies reported that autophagy is mediated by beclin-1 and AMPK-independent pathways during myocardial ischemia and reperfusion (Takagi et al. 2007). We further observed an increase of beclin-1 levels in HO-1 overexpressed cardiomyocytes. Indeed, our results suggested that HO-1 overexpression induces autophagy to protect myocardial cells from H/R injury.
In conclusion, this study demonstrated that HO-1 overexpression could protect cardiomyocytes against H/R injury through anti-apoptosis and cell autophagy through Sirt3 signaling pathway. Accordingly, our study provided evidence for the future clinical potential targets to treat cardiac hypoxia/ischemia.
Change history
18 August 2021
A Correction to this paper has been published: https://doi.org/10.1007/s00210-021-02142-x
Abbreviations
- H/R:
-
Hypoxia/reoxygenation
- ROS:
-
Reactive oxygen species
- Sirt3:
-
Sirtuin3
- HO-1:
-
Heme oxygenase-1
- BrdU:
-
5-Bromo-2-deoxyUridine
- MtDNA:
-
Mitochondrial DNA
- LC3:
-
Light chain 3
- AMPK:
-
Adenosine 5′-monophosphate (AMP)-activated protein kinase
References
Bonavita F, Stefanelli C, Giordano E, Columbaro M, Facchini A, Bonafe F, Caldarera CM, Guarnieri C (2003) H9c2 cardiac myoblasts undergo apoptosis in a model of ischemia consisting of serum deprivation and hypoxia: inhibition by PMA. FEBS Lett 536(1–3):85–91
Chen D, Jin Z, Zhang J, Jiang L, Chen K, He X, Song Y, Ke J, Wang Y (2016) HO-1 protects against hypoxia/reoxygenation-induced mitochondrial dysfunction in H9c2 cardiomyocytes. PLoS One 11(5):e0153587
Dai SH, Chen T, Li X, Yue KY, Luo P, Yang LK, Zhu J, Wang YH, Fei Z, Jiang XF (2017) Sirt3 confers protection against neuronal ischemia by inducing autophagy: involvement of the AMPK-mTOR pathway. Free Radic Biol Med 108:345–353
Dong C, Zheng H, Huang S, You N, Xu J, Ye X, Zhu Q, Feng Y, You Q, Miao H, Ding D, Lu Y (2015) Heme oxygenase-1 enhances autophagy in podocytes as a protective mechanism against high glucose-induced apoptosis. Exp Cell Res 337(2):146–159
Du Y, Zhang J, Fang F, Wei X, Zhang H, Tan H, Zhang J (2017) Metformin ameliorates hypoxia/reoxygenation-induced cardiomyocyte apoptosis based on the SIRT3 signaling pathway. Gene 626:182–188
Duan Q, Yang W, Jiang D, Tao K, Dong A, Cheng H (2016) Spermine ameliorates ischemia/reperfusion injury in cardiomyocytes via regulation of autophagy. Am J Transl Res 8(9):3976–3985
Fujimoto S, Mizuno R, Saito Y, Nakamura S (2004) Clinical application of wave intensity for the treatment of essential hypertension. Heart Vessel 19(1):19–22
Gottlieb RA, Mentzer RM (2010) Autophagy during cardiac stress: joys and frustrations of autophagy. Annu Rev Physiol 72:45–59
Gustafsson AB, Gottlieb RA (2009) Autophagy in ischemic heart disease. Circ Res 104(2):150–158
Haunstetter A, Izumo S (1998) Apoptosis: basic mechanisms and implications for cardiovascular disease. Circ Res 82(11):1111–1129
Hausenloy DJ, Yellon DM (2013) Myocardial ischemia-reperfusion injury: a neglected therapeutic target. J Clin Invest 123(1):92–100
He W, Su Q, Liang J, Sun Y, Wang X, Li L (2018) The protective effect of nicorandil on cardiomyocyte apoptosis after coronary microembolization by activating Nrf2/HO-1 signaling pathway in rats. Biochem Biophys Res Commun 496(4):1296–1301
Klionsky DJ, Emr SD (2000) Autophagy as a regulated pathway of cellular degradation. Science 290(5497):1717–1721
Kong X, Wang R, Xue Y, Liu X, Zhang H, Chen Y, Fang F, Chang Y (2010) Sirtuin 3, a new target of PGC-1alpha, plays an important role in the suppression of ROS and mitochondrial biogenesis. PLoS One 5(7):e11707
Li C, Zhang C, Wang T, Xuan J, Su C, Wang Y (2016a) Heme oxygenase 1 induction protects myocardiac cells against hypoxia/reoxygenation-induced apoptosis : the role of JNK/c-Jun/caspase-3 inhibition and Akt signaling enhancement. Herz 41(8):715–724
Li J, Chen T, Xiao M, Li N, Wang S, Su H, Guo X, Liu H, Yan F, Yang Y, Zhang Y, Bu P (2016b) Mouse Sirt3 promotes autophagy in AngII-induced myocardial hypertrophy through the deacetylation of FoxO1. Oncotarget 7(52):86648–86659
Maines MD (1997) The heme oxygenase system: a regulator of second messenger gases. Annu Rev Pharmacol Toxicol 37:517–554
Matsui Y, Takagi H, Qu X, Abdellatif M, Sakoda H, Asano T, Levine B, Sadoshima J (2007) Distinct roles of autophagy in the heart during ischemia and reperfusion: roles of AMP-activated protein kinase and Beclin 1 in mediating autophagy. Circ Res 100(6):914–922
Melo LG, Agrawal R, Zhang L, Rezvani M, Mangi AA, Ehsan A, Griese DP, Dell'Acqua G, Mann MJ, Oyama J, Yet SF, Layne MD, Perrella MA, Dzau VJ (2002) Gene therapy strategy for long-term myocardial protection using adeno-associated virus-mediated delivery of heme oxygenase gene. Circulation 105(5):602–607
Michishita E, Park JY, Burneskis JM, Barrett JC, Horikawa I (2005) Evolutionarily conserved and nonconserved cellular localizations and functions of human SIRT proteins. Mol Biol Cell 16(10):4623–4635
Mortality GBD, C. Causes of Death (2015) Global, regional, and national age-sex specific all-cause and cause-specific mortality for 240 causes of death, 1990–2013: a systematic analysis for the Global Burden of Disease Study 2013. Lancet 385(9963):117–171
Murphy E, Steenbergen C (2008) Mechanisms underlying acute protection from cardiac ischemia-reperfusion injury. Physiol Rev 88(2):581–609
Oerlemans MI, Koudstaal S, Chamuleau SA, de Kleijn DP, Doevendans PA, Sluijter JP (2013) Targeting cell death in the reperfused heart: pharmacological approaches for cardioprotection. Int J Cardiol 165(3):410–422
Paine A, Eiz-Vesper B, Blasczyk R, Immenschuh S (2010) Signaling to heme oxygenase-1 and its anti-inflammatory therapeutic potential. Biochem Pharmacol 80(12):1895–1903
Pillai VB, Bindu S, Sharp W, Fang YH, Kim G, Gupta M, Samant S, Gupta MP (2016) Sirt3 protects mitochondrial DNA damage and blocks the development of doxorubicin-induced cardiomyopathy in mice. Am J Physiol Heart Circ Physiol 310(8):H962–H972
Shelton LM, Park BK, Copple IM (2013) Role of Nrf2 in protection against acute kidney injury. Kidney Int 84(6):1090–1095
Shi X, Li Y, Hu J, Yu B (2016) Tert-butylhydroquinone attenuates the ethanol-induced apoptosis of and activates the Nrf2 antioxidant defense pathway in H9c2 cardiomyocytes. Int J Mol Med 38(1):123–130
Takagi H, Matsui Y, Hirotani S, Sakoda H, Asano T, Sadoshima J (2007) AMPK mediates autophagy during myocardial ischemia in vivo. Autophagy 3(4):405–407
Tang S, Ma D, Cheng B, Fang Q, Kuang X, Yu K, Wang W, Hu B, Wang J (2018a) Crucial role of HO-1/IRF4-dependent apoptosis induced by panobinostat and lenalidomide in multiple myeloma. In: Exp cell res, vol 363, pp 196–207
Tang, X., B. Liu, X. Wang, Q. Yu and R. Fang (2018b). Epidermal growth factor, through alleviating oxidative stress, protect IPEC-J2 cells from lipopolysaccharides-induced apoptosis. Int J Mol Sci 19(3):848–856
Tullius SG, Nieminen-Kelha M, Buelow R, Reutzel-Selke A, Martins PN, Pratschke J, Bachmann U, Lehmann M, Southard D, Iyer S, Schmidbauer G, Sawitzki B, Reinke P, Neuhaus P, Volk HD (2002) Inhibition of ischemia/reperfusion injury and chronic graft deterioration by a single-donor treatment with cobalt-protoporphyrin for the induction of heme oxygenase-1. Transplantation 74(5):591–598
Vulapalli SR, Chen Z, Chua BH, Wang T, Liang CS (2002) Cardioselective overexpression of HO-1 prevents I/R-induced cardiac dysfunction and apoptosis. Am J Physiol Heart Circ Physiol 283(2):H688–H694
Wang B, Zhong S, Zheng F, Zhang Y, Gao F, Chen Y, Lu B, Xu H, Shi G (2015a) N-n-butyl haloperidol iodide protects cardiomyocytes against hypoxia/reoxygenation injury by inhibiting autophagy. Oncotarget 6(28):24709–24721
Wang H, Sun HQ, Zhu X, Zhang L, Albanesi J, Levine B, Yin H (2015b) GABARAPs regulate PI4P-dependent autophagosome:lysosome fusion. Proc Natl Acad Sci U S A 112(22):7015–7020
Wei C, Li H, Wang Y, Peng X, Shao H, Li H, Bai S, Xu C (2016) Exogenous spermine inhibits hypoxia/ischemia-induced myocardial apoptosis via regulation of mitochondrial permeability transition pore and associated pathways. Exp Biol Med (Maywood) 241(14):1505–1515
Wei W, Shurui C, Zipeng Z, Hongliang D, Hongyu W, Yuanlong L, Kang Z, Zhaoliang S, Yue G, Chang L, Mei X (2018) Aspirin suppresses neuronal apoptosis, reduces tissue inflammation, and restrains astrocyte activation by activating the Nrf2/HO-1 signaling pathway. Neuroreport 29:524–531
Yan WJ, Liu RB, Wang LK, Ma YB, Ding SL, Deng F, Hu ZY, Wang DB (2018) Sirt3-mediated autophagy contributes to resveratrol-induced protection against ER stress in HT22 cells. Front Neurosci 12:116
Yu H, Shi L, Zhao S, Sun Y, Gao Y, Sun Y, Qi G (2016) Triptolide attenuates myocardial ischemia/reperfusion injuries in rats by inducing the activation of Nrf2/HO-1 defense pathway. Cardiovasc Toxicol 16(4):325–335
Author information
Authors and Affiliations
Contributions
F.S. designed the study. X.L., Y.Y., and C.L. performed the experiments and collected the data. C.L. and F.S. analyzed and interpreted the experimental data. F.S. and C.L. prepared the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
This article has been retracted. Please see the retraction notice for more detail:https://doi.org/10.1007/s00210-021-02142-x
About this article

Cite this article
Meng, X., Yuan, Y., Shen, F. et al. RETRACTED ARTICLE: Heme oxygenase-1 ameliorates hypoxia/reoxygenation via suppressing apoptosis and enhancing autophagy and cell proliferation though Sirt3 signaling pathway in H9c2 cells. Naunyn-Schmiedeberg's Arch Pharmacol 392, 189–198 (2019). https://doi.org/10.1007/s00210-018-1575-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00210-018-1575-4