
Abstract
Genotype × environment interaction effects can be exploited by breeding for specific adaptation to well-defined subregions within a target region. Previous work showed that genotype × location interaction for dry matter (DM) yield of lucerne (Medicago sativa L. subsp. sativa) cultivars in northern Italy is large and associated with soil type and level of summer drought stress of locations, suggesting the presence of two contrasting subregions. Thirteen farm landraces collected across the region and four control varieties were evaluated for DM yield in four artificial environments created at one site by the factorial combination of soil type (sandy loam or silty clay) and drought stress level (almost nil or high) for: (1) exploring the possibility to reproduce in artificial environments the adaptation patterns occurring across the region; (2) investigating the adaptation pattern of landraces and its relationship with environmental factors at collecting sites; and (3) providing a preliminary comparison of wide- versus specific-adaptation strategies based on yield gains predicted from selection of populations. Different soils filled large (24.0×1.6×0.8-m deep), bottomless containers in concrete. Water amounts were controlled by irrigation under a moving rain shelter. Cultivars varied largely for adaptation pattern across the artificial environments, mainly due to cultivar × stress interaction. Better response to stress conditions of landraces was closely associated with the level of summer drought at collecting sites (r=0.82), highlighting the importance of evolutionary adaptation. The additive main effects and multiplicative interaction-modelled responses of control cultivars successfully reproduced those observed across locations, candidating the artificial environments as a cheaper alternative to more selection locations when breeding for wide or specific adaptation. The latter implied about 40–50% greater estimated gains relative to breeding for wide adaptation.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Breeding for specific adaptation to well-defined subregions within a target region is major strategy for exploiting genotype × environment (GE) interaction to increase crop yields (Ceccarelli 1989; Simmonds 1991; Annicchiarico 2002a). In addition, this strategy enforces sustainable agriculture by two means, namely (1) maximizing the potential of different areas by fitting cultivars to an environment, instead of altering the environment by costly or environment-unfriendly inputs to fit widely adapted cultivars, and (2) contributing to safeguard crop biodiversity by increasing the number of varieties under cultivation (Bramel-Cox et al. 1991; Ceccarelli 1996). However, a specific-adaptation strategy needs be justified on the ground of costs and benefits. Its advantage depends on the extent of repeatable genotype × location (GL) interaction effects (Annicchiarico 1997), which should be high enough to ensure definitely greater selection gains relative to breeding for wide adaptation (Annicchiarico 2002b). Other practical considerations, such as the commercial importance of candidate subregions and the availability of test sites within subregions, may contribute to decisions on the adaptation strategy.
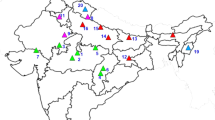
GL interaction effects for dry matter (DM) yield of lucerne cultivars in northern Italy, analysed by additive main effects and multiplicative interaction (AMMI) modelling, proved large and implied cross-over interaction of top-ranking material between contrasting locations (Annicchiarico 1992). There was evidence of specific adaptation to the area where the variety was selected and/or where its genetic base originated. The GL effects were largely repeatable in an independent data set (Annicchiarico 2000), suggesting the opportunity to exploit them by recommending specifically adapted cultivars and, in the long run, by breeding for specific adaptation. Site classification for GL effects consistently revealed the presence of two contrasting subregions and was mainly related to soil type and level of summer drought stress of locations. These subregions are reported as A and C in Fig. 1. The former is characterized by sandy loam-to-loam soil, and by limited drought stress, mainly due to irrigated cropping; the latter tends to clay soil, as well as substantial drought stress due to rain-fed cropping and somewhat lower rainfall. Subregion B in Fig. 1 is intermediate geographically, for response of varieties and for environmental characteristics. While targeting the whole northern Italy, our breeding programme is based at Lodi, i.e. in subregion A (Fig. 1), a position that makes it difficult to breed for wide adaptation to the region or for specific adaptation also to subregion C. Adopting an additional selection site in subregion C would be a convenient solution that is hindered, however, by the high costs entailed. An alternative option may be the selection of material in artificial environments that reproduce the variation across the region for crucial environmental factors affecting GL interaction, as proposed by Federer and Scully (1993) and accomplished in a few instances, e.g. for bread wheat in Australia (Cooper et al. 1995) and durum wheat in Italy (Annicchiarico and Mariani 1996).

Geographical subdivision of the target region into three subregions, and position of collecting sites of lucerne landraces
In Italy, lucerne landraces have largely been grown until 2002, when marketing their seed has been prohibited, and represent the core of the genetic base used by breeding programmes (Russi and Falcinelli 1997). With reference to a set of farm landraces collected across northern Italy and to a smaller set of control varieties of known adaptation pattern, this study aimed at: (1) exploring the possibility to reproduce in artificial environments at Lodi the adaptation patterns occurring across northern Italy, (2) investigating the adaptation pattern of landraces and its relationship with environmental factors at collecting sites and (3) providing a preliminary comparison of wide- vs. specific-adaptation strategies based on the extent of expected yield gains.
Materials and methods
Experimental procedures
For seed marketing, Italian farm landraces of lucerne have been grouped into 14 commercial ecotypes depending on their area of production, of which seven are located in northern Italy. Twenty landraces were collected in 1995 and 1996 from farmers that had been multiplying their seed for at least 20 years. This study included a subset of 13 landraces that roughly represented each of the seven commercial ecotypes in proportion to its market share and tended to maximize, whenever possible, the geographical variation within ecotype. Figure 1 reports the geographical position, and Table 1 the name, geographical co-ordinates, soil type according to FAO (1990, p 70) soil texture classes and level of summer drought stress of the collecting sites. Drought stress was estimated as the difference between potential evapotranspiration (ETP) and average rainfall plus irrigation over June, July and August, using long-term ETP and rainfall values of the nearest meteorological station in the report by Dal Monte et al. (1995). The varieties ‘Europe’, ‘La Rocca’ and ‘Prosementi’, well-characterized and contrasting for adaptation pattern across the region (Table 2), acted as ‘probe genotypes’ as defined by Wade et al. (1996), i.e. material of which the known adaptive response is exploited to assign each test environment to a given class of target environments (subregion). Our recent variety ‘Lodi’ was added to the study to further verify whether selection in subregion A produces material specifically adapted to this subregion.
The variation across the region for major environmental factors associated with GL interaction was reproduced in Lodi by four artificial environments created by the factorial combination of type of soil (sandy loam or silty clay) and level of drought stress in summer (almost nil or high). Local soil, which is sandy loam close to loam (clay = 12%, sand = 55% silt = 33%), or silty clay soil (clay = 41%, sand = 9%, silt = 50%) imported from subregion C, filled large (24.0×1.6×0.8-m deep), bottomless containers in concrete laid in a field. Irrigated or rain-fed cropping during an ordinary summer season were simulated in each environment by irrigation under moving rain-shelter equipment that covered the crop in rainy days over 12 weeks, starting from 10 June. During this period, drought-stress environments received a 40-mm irrigation after 4 weeks and 10 weeks, whereas no-stress environments received a 90-mm irrigation after 1, 4, 7 and 10 weeks. Water amounts were computed on the basis of average summer rainfall in subregion C for stress environments, and average summer rainfall plus ordinary level of irrigation in subregion A for no-stress environments.
Within each artificial environment, the 17 entries were grown in a randomized complete block design with four replicates. Each plot consisted of 70 plants arranged in a 7×10 layout and spaced 7.5 cm between rows and on the row. The harvest area comprised 40 plants, excluding each edge row and edge plant on the row. About 1-month-old seedlings previously grown in Jiffy pots were transplanted at the end of September 1996. DM yield was recorded over six harvests in 1997, five harvests in 1998 and the first harvest of 1999. All environments were fertilized with 50 kg of N, 150 kg of P2O5 and 150 kg of K2O prior to transplanting.
Statistical analysis
Variation for total DM yield over harvests was partitioned into ordinary sources of variation for the analysis of variance (ANOVA) of multi-environment trials, namely genotype (i.e. cultivar), environment, block within environment, GE interaction and pooled error. The variation for environment and GE interaction was further partitioned into contrasts relative to soil, drought stress and their interaction. In particular, the GE ij effects relative to the genotype i and the environment j were partitioned into genotype × soil (GS), genotype × drought stress (GD) and genotype × soil × drought stress (GSD) interaction effects relative to the k soil and the l drought stress level:
Estimated GS ik and GD il effects for the genotypes were simultaneously represented by a biplot.
GE ij interaction effects were also partitioned by AMMI analysis into:
where u in and v jn are eigenvectors of the genotype i and the environment j, respectively, and l n is the singular value for the principal component (PC) axis n; and d ij is the deviation from the model. The scaling of eigenvectors through multiplication by √l n , as in the latter expression, allows for a straightforward estimation of the GE ij effects expected on the PC axis n by multiplication of the scaled genotype and location scores on that axis (Gauch, 1992, p 86). Statistical testing of PC axes was performed by the F R test recommended by Piepho (1995). Scaled scores of genotypes and environments on the first two PC axes were jointly represented in a second biplot. Relationships of GS ik and GD il effects with scaled PC scores of cultivars, or between these variables and environmental variables at the collecting site of landraces, were investigated by simple correlation analysis.
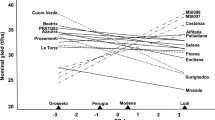
Nominal yield responses of genotypes, corresponding to expected yields from which the site main effect (irrelevant for genotype ranking on the site) has been eliminated, were estimated according to Gauch and Zobel (1997) by pooling mean yield and GE interaction effects as modelled by AMMI-1, thereby allowing for graphically representing the adaptive responses as straight lines as a function of the scaled PC 1 score of artificial environments. These responses were compared with nominal DM yield responses of varieties subject to multi-locational testing in Annicchiarico (1992), which could conveniently be expressed by the AMMI-1 model after eliminating from the original data set two mountain locations excluded from subregion definition in Fig. 1 and related to GL interaction variation on a second PC axis.
Since drought stress exerted a distinctly greater effect than soil type on the occurrence of GE interaction, a specific-adaptation strategy targeted separately a low-stress subregion by selecting across no-stress artificial environments, and a high-stress subregion by selecting across stress environments. This prospect was compared with a wide-adaptation scenario implying the parallel selection across all artificial environments. The comparison, relative to selection of populations as represented by the sample of tested material, was based on predicted yield gains as proposed by Annicchiarico (2002a, p 67; 2002b). Genotypic (s 2 g ) and GE interaction (s 2 ge ) variance components were estimated from ANOVA mean squares according to model 3 in Table 4.1 of Annicchiarico (2002a, p 23), i.e. assuming genotype as random factor and environment as fixed, for the following three sets of environments representing the relevant adaptation targets: (1) no-stress environments, (2) stress environments and (3) all environments. For each adaptation target, the predicted yield gain (ΔG) was estimated as:
where i is the standardized selection differential, h2 is the broad-sense heritability on an entry mean basis, and s p is the phenotypic standard deviation. Indicating by E and R the number of environments and experiment replicates, respectively, hypothesized for the selection work, h2 was estimated as:
, and s p was estimated by the square root of the denominator of the same formula. E was equal to 2 for each of the two specific-adaptation targets and to 4 for the wide-adaptation target (implying same number of overall selection environments, i.e. 4, for the two adaptation strategies). R was set to 1, thereby increasing to 80 the number of possibly tested genotypes in the current environments, assuming ten selected genotypes per cycle (i.e. 12.5% selection intensity, i=1.621, Falconer 1989, p 355). A simpler comparison of the two selection strategies was provided by the yield comparison in no-stress and in stress environments of the top-ranking entry chosen according to a wide- and a specific-adaptation prospect. The procedure for predicting yield gains in target subregions from yield responses in artificial environments described by Basford et al. (2004) was not applicable here due to the limited number of cultivars also subject to previous multi-locational testing.
AMMI analysis was performed by the software IRRISTAT (version 4.3), released by the International Rice Research Institute. The statistical analysis system (SAS) software was used for the remaining analyses.
Results
Cultivars differed for mean yield at the P≤0.05 level in the ANOVA (Table 3). Only five cultivars and no control variety showed lower mean yield (P≤0.05) than the top-yielding genotype, i.e. landrace 17 (Tables 1, 2). The variation between environments was significant (Table 3), mainly due to higher yield from no-stress relative to stress environments (+31%). In addition, soil type had an effect, with silty clay soil being higher yielding than sandy loam soil (+10%), probably because of its greater water holding capacity. GE interaction was highly significant, mainly due to GD interaction (Table 3), whereas GS interaction just attained P≤0.06 significance. The estimated values of components of variance across all environments were 0.99 t/ha2 for genotype and 4.40 t/ha2 for GE interaction, indicating the much greater extent of GE effects relative to genotype main effects.
Landraces 10, 15, 19 and 18, collected in subregions A or B (Fig. 1), and varieties ‘Lodi’ and ‘La Rocca’, showed sizeable positive GE effects towards no-stress environments (Fig. 2a). Landraces 7, 3 and 8, originated in subregions C or B, and ‘Europe’ showed the opposite pattern and were, therefore, better adapted to stress environments. GE effects suggesting some degree of specific adaptation to soil type were showed by a few landraces (17, 16 and 1) collected in subregions A or B and by ‘La Rocca’ towards sandy loam soil, and by landrace 3 collected in subregion C towards silty clay soil (Fig. 2a).

Genotype × drought stress and genotype × soil type interaction effects (a), and score on the first (PC 1) and the second (PC 2) GE interaction principal component axis (b), for lucerne cultivars grown in artificial environments
In the AMMI analysis, PC 1 and PC 2 reached P≤0.01 and P≤0.05 significance, respectively (Table 3). PC 1 accounted for 47% and PC 2 for 30% of GE interaction variation. The AMMI biplot is reported in Fig. 2b. Based on the environment scores, PC 1 maximized the GE effects along a gradient from no-stress to stress environments; PC 2 represented mainly a contrast between soil types within stress environments, suggesting that specific adaptation to soils emerged under stress conditions (as confirmed by the presence of GS interaction at P≤0.01 in a further ANOVA limited to stress environments, and in agreement with the trend towards genotype × soil × stress interaction in the ANOVA reported in Table 3). As a result, the indications of Fig. 2a, b were almost identical for genotype responses to stress levels (the specific adaptation to no-stress and to stress being indicated by low and high cultivar value, respectively, on the horizontal axis), but they differed somewhat for genotype responses to soil types (represented by cultivar values on the vertical axis), e.g. there was no evidence of specific adaptation of landrace 1 to sandy loam soil in Fig. 2b. Accordingly, a very high correlation (r=0.99) emerged between genotype × stress interaction effects of cultivars (defined as positive for better response to drought) and genotype PC 1 score, whereas the correlation between genotype × soil type interaction effects and PC 2 scores of material was only moderately high (Table 4). No correlation was found between mean yield of genotypes and any measure of genotype adaptation.
Relatively better response of landraces to drought stress conditions was closely associated with high level of summer drought in their environment of origin (r=0.82) and its contributing variables, i.e. low water for the crop and high ETP over the summer period (Table 4). Indeed, the somewhat unexpected response to stress levels of some landraces in relation to their subregion of origin (e.g. the specific adaptation of landraces 7 and 19, both from subregion B, respectively to stress and no-stress conditions; the lack of specific adaptation to no-stress conditions of landrace 16, collected in subregion A) could be explained by the specific level of drought stress in their environment of origin (Table 1), especially as determined by irrigation level at the farm. There was a slight relationship between specific adaptation to soil type of artificial environments and soil content of sand in the environment of origin of the landraces, and no relationship between mean yield across environments and drought stress or soil type at collecting sites (Table 4).
Figure 3a reports the AMMI-1 nominal yield responses of a subset of top-yielding cultivars as a function of site score on the first GL interaction PC axis in the reference multi-locational data set. PC 1 was significant at P≤0.01, according to the F R test. Site classification into subregions that is showed in Fig. 3a summarizes previous findings, which are reported in Annicchiarico (1992) together with the name and the geographical position of coded locations. With regard to the three reference cultivars, ‘La Rocca’ was top yielding in subregion A, ‘Prosementi’ was best ranking in subregion B and tended to a wide-adaptation pattern, and ‘Europe’ was top yielding in subregion C. These responses were compared with AMMI-1 nominal yields of the three reference varieties across artificial environments, which are reported in Fig. 3b together with those of other top-performing material. Based on this model, including only GE effects significant at P≤0.01, the most contrasting environments for genotype responses were the no-stress, sandy loam one, representing subregion A, and the stress, silty loam one, representing subregion C. For subregion B, not represented by any artificial environment but characterized by intermediate soil type and drought stress level, genotype responses may be inferred by those around zero value of environment PC 1 score. The superiority of ‘La Rocca’ in the no-stress, sandy loam environment, ‘Europe’ in the stress, silty loam environment, and ‘Prosementi’ around zero value of environment PC 1 score (Fig. 3b) indicated that the set of artificial environments could successfully reproduce the adaptation patterns observed across the target region and the genotype ranking in individual subregions. As a result, responses to subregions of other cultivars may be inferred from their responses to artificial environments. These responses suggested the specific adaptation of ‘Lodi’ to its selection subregion A, and the trend for most landrace material towards a specific adaptation pattern (Fig. 3b).

Nominal yields of top-yielding lucerne cultivars as a function of the site score on the first GL interaction PC axis in a reference data set (a) or the score on the first GE interaction PC axis of artificial environments (b)
Given the major effect of drought stress on the extent of GE interaction across the artificial environments reproducing the GL effects across the region, a specific-adaptation strategy could conveniently target a low-stress subregion on the one hand (including subregion A and less stressful areas of subregion B) and a high-stress subregion on the other (including subregion C and more stressful areas of subregion B). The predicted yield gains over the target environments for this strategy were distinctly greater than those predicted over all environments for a wide-adaptation strategy (Table 5), since the minimization of GE interaction effects due to GD interaction allowed for by this strategy resulted in higher heritability and phenotypic standard deviation values. The average yield gain predicted for specific adaptation (1.387 t/ha) was about twice as large as that for wide adaptation (Table 5).
The top-yielding cultivar differed when assessed on the basis of mean yield across all environments, no-stress environments and stress environments (Table 6). The yield difference of the top-ranking material relative to mean value of all entries, which provides an indication of the yield gain possibly associated with the selection of a given population as genetic base for a breeding programme, was also computed (Table 6). Compared with widely adapted material, specifically adapted one implied greater gains of 42% for no-stress (4.59/3.24×100) and 41% for stress environments, respectively.
Discussion
The definition of breeding strategies has been the target of GE interaction analyses in a minority of studies (Basford and Cooper 1998), which, in turn, have been followed by a documented assessment of novel strategies suggested by their results in just a few cases (e.g. barley in Syria, Ceccarelli 1996; Ceccarelli et al. 1998). Our work provides one such example, since earlier information on GL interaction in the target region and on possibly contributing environmental factors (Annicchiarico 1992, 2000) has been exploited for defining and preliminarily assessing alternative adaptation strategies and selection environments. While being limited to lucerne in northern Italy in its practical implications, our experience is in the stream of the ‘thinking globally, acting locally’ concept expressed by Fischer (1996) to define the plant breeding effort, global as a trend but local in its forms, towards an increasing exploitation of favourable GE interaction effects.
Despite the high level of heterogeneity and heterozygosity that usually characterizes their genetic structure, lucerne cultivars may show large GE interaction even within relatively small regions, such as Wisconsin (Casler and Undersander 2000) or northern Italy. For the latter region, the evaluation in artificial environments has suggested that drought stress has a greater impact than soil type on the variation in adaptive responses. These environmental factors tend to be associated across the region, owing to the fact that the higher water holding capacity of clayish soils facilitates rain-fed cropping. This study has also revealed the wide variation in adaptive responses among landraces originated in the region and highlighted, based on the close relationship between these responses and the characteristics of the environment of origin of the material, the major role of evolutionary adaptation in this context. The agronomic value of these landraces cannot reliably be evaluated in only one environment, as it may be largely underestimated in the presence of drought stress levels that contrasts with those of the environment of origin. Although there is large evidence of landrace material specifically adapted to drought stress environments (e.g. Muñoz et al. 1998; van Oosterom et al. 2003), the present study indicates that landraces may also be specifically adapted to favourable conditions, provided that they evolved under such conditions. It is noteworthy that several landraces showed an agronomic value comparable with, or even superior to, that of improved varieties, confirming their interest as fundamental genetic resources in breeding for the region. Lucerne landraces still play a major role as genetic resources in another European country with a long-standing breeding activity, i.e. France (Julier 1996).
The ability of the set of artificial environments to reproduce adaptation patterns occurring across the region may be exploited to breed for wide or for specific adaptation, without necessarily adopting additional selection locations. Since the influence of soil type on GE interaction effects was not negligible, at least under stress conditions, the parallel selection across contrasting soils has the advantage of selecting for wide adaptation to soil type, while breeding for wide or specific adaptation with regard to drought stress.
Selection schemes for lucerne basically relate to individual plants. Therefore, the comparison of adaptation strategies based on selection of populations (Table 5) is only preliminary and shall be validated by ongoing research aimed at estimating actual yield gains relative to phenotypic and genotypic selection. In particular, the comparison based on predicted yield gains may be reliable in so far as the variation among populations may represent that for additive genetic effects among individuals sorted out by the same germplasm pool. The interest of the yield comparison among the three top-ranking landraces chosen according to different adaptation targets (Table 6) is reinforced by the fact that these cultivars tend to similar within-population variation for DM yield (data not reported) and, as such, may provide similar yield gains when chosen as the unique genetic base for selection of individuals. Despite their limitations, the current comparisons of adaptations strategies provided very similar results, suggesting that breeding for specific adaptation may be about twice as efficient as breeding for wide adaptation to the region. It should be noted that the comparisons implied same number of total selection environments (i.e. four) for each strategy and, hence, similar costs.
Our artificial environments, given the limited number of possibly tested entries, may be used for further evaluation of elite material issued by earlier phenotypic selection. In particular, they may be used for genotypic selection based on progenies from polycross or selfing. Earlier stages of selection may be performed under field conditions in the local sandy loam soil by imposing a moderately high drought stress when breeding for wide adaptation (a condition approaching the zero value of environment PC 1 score in Fig. 3b), and by separate selection environments that contrast for drought stress level when pursuing a specific-adaptation strategy. The adoption of specific selection environments also in early selection stages is expected to further increase the overall advantage of breeding for specific adaptation (here assessed only with reference to late selection stages). Maximizing selection gains can be particularly important for species, such as lucerne, in which the perennial cycle and the selection for total biomass (rather than seed yield) hinder the breeding progress.
References
Annicchiarico P (1992) Cultivar adaptation and recommendation from alfalfa trials in northern Italy. J Genet Breed 46:269–278
Annicchiarico P (1997) Additive main effects and multiplicative interaction (AMMI) analysis of genotype-location interaction in variety trials repeated over years. Theor Appl Genet 94:1072–1077
Annicchiarico P (2000) Variety × location interaction and its implications on breeding of lucerne: a case study. In: Veronesi F, Rosellini D (eds) Lucerne and medics for the XXI century. University of Perugia, Perugia, pp 35–43
Annicchiarico P (2002a) Genotype × environment interactions: challenges and opportunities for plant breeding and cultivar recommendations. FAO plant production protection paper 174. FAO, Rome
Annicchiarico P (2002b) Defining adaptation strategies and yield stability targets in breeding programmes. In: Kang MS (ed) Quantitative genetics, genomics and plant breeding. CABI, Wallingford, pp 365–383
Annicchiarico P, Mariani G (1996) Prediction of adaptability and yield stability of durum wheat genotypes from yield response in normal and artificially drought-stressed conditions. Field Crops Res 46:71–80
Basford KE, Cooper M (1998) Genotype × environment interactions and some considerations of their implications for wheat breeding in Australia. Aust J Agric Res 49:153–174
Basford KE, Federer WT, DeLacy IH (2004) Mixed model formulation for multi-environment trials. Agron J 96:143–147
Bramel-Cox PJ, Barker T, Zavala-Garcia F, Eastin JD (1991) Selection and testing environments for improved performance under reduced-input conditions. In: Sleper D, Bramel-Cox PJ, Barker T (eds) Plant breeding and sustainable agriculture: considerations for objectives and methods. CSSA Special Publication, vol 18. ASA, CSSA, SSSA, Madison, pp 29–56
Casler MD, Undersander DJ (2000) Forage yield precision, experimental design, and cultivar mean separation for alfalfa cultivar trials. Agron J 92:1064–1071
Ceccarelli S (1989). Wide adaptation: how wide? Euphytica 40:197–205
Ceccarelli S (1996) Positive interpretation of genotype by environment interaction in relation to sustainability and biodiversity. In: Cooper M, Hammer GL (eds) Plant adaptation and crop improvement. CABI, Wallingford, pp 467–486
Ceccarelli S, Grando S, Impiglia A (1998) Choice of selection strategy in breeding barley for stress environments. Euphytica 103:307–318
Cooper M, Woodruff DR, Eisemann RL, Brennan PS, DeLacy IH (1995) A selection strategy to accommodate genotype-by-environment interaction for grain yield of wheat: managed-environments for selection among genotypes. Theor Appl Genet 90:492–502
Dal Monte G, Perini L, Brunetti A (1995) Indici agroclimatici. Ufficio Centrale Ecologia Agraria, Rome
Falconer DS (1989) Introduction to quantitative genetics, 3rd edn. Longman, New York
FAO (1990) Guidelines for soil profile description. FAO, Rome
Federer WT, Scully BT (1993) A parsimonious statistical design and breeding procedure for evaluating and selecting desirable characteristics over environments. Theor Appl Genet 86:612–620
Fischer KS (1996) Research approaches for variable rainfed system. Thinking globally, acting locally. In: Cooper M, Hammer GL (eds) Plant adaptation and crop improvement. CABI, Wallingford, pp 25–35
Gauch HG (1992) Statistical analysis of regional yield trials: AMMI analysis of factorial designs. Elsevier, Amsterdam
Gauch HG, Zobel RW (1997) Identifying mega-environments and targeting genotypes. Crop Sci 37:311–326
Julier B (1996) Traditional seed maintenance and origins of the French lucerne landraces. Euphytica 92:353–357
Muñoz P, Voltas J, Araus JL, Igartua E, Romagosa I (1998) Changes over time in the adaptation of barley releases in north-eastern Spain. Plant Breed 117:531–535
van Oosterom EJ, Bidinger FR, Weltzien ER (2003) A yield architecture framework to explain adaptation of pearl millet to environmental stress. Field Crops Res 80:33–56
Piepho HP (1995) Robustness of statistical tests for multiplicative terms in the additive main effects and multiplicative interaction model for cultivar trials. Theor Appl Genet 90:438–443
Russi L, Falcinelli M (1997) Characterization and agronomic value of Italian landraces of lucerne (Medicago sativa). J Agric Sci (Camb) 129:267–277
Simmonds NW (1991) Selection for local adaptation in a plant breeding programme. Theor Appl Genet 82:363–367
Wade LJ, McLaren CG, Samson BK, Regmi KR, Sarkarung S (1996) The importance of environmental characterization for understanding genotype by environment interactions. In: Cooper M, Hammer GL (eds) Plant adaptation and crop improvement. CABI, Wallingford, pp 549–561
Acknowledgements
We gratefully acknowledge S. Proietti for excellent technical assistance.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H.C. Becker
Rights and permissions
About this article
Cite this article
Annicchiarico, P., Piano, E. Use of artificial environments to reproduce and exploit genotype × location interaction for lucerne in northern Italy. Theor Appl Genet 110, 219–227 (2005). https://doi.org/10.1007/s00122-004-1811-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00122-004-1811-9