
Abstract
Characterization of crop-growing environments in relation to crop’s genotypic performance is crucial to harness positive genotype-by-environment interactions (GEI) in systematic breeding programs. Given that, the study aimed to delineate the impact of diverse environments on crop phenology and yield traits of dwarf-statured field pea, pinpointing location(s) favoring higher yield and distinctiveness within breeding lines. We tested twelve field pea breeding lines across twenty locations in India, covering Central Zone (CZ), North Western Plain Zone (NWPZ), North Eastern Plain Zone (NEPZ), and Northern Hill Zone (NHZ). Across these locations, maximum and minimum temperatures during flowering (TMAXF, TMINF) and reproductive period (TMAXRP, TMINRP) ranged 18.9–28.3, 3.3–18.0, 15.0-30.8, and 7.9-22.1oC, respectively. Meanwhile, notable variations in phenological and agronomic traits (coefficient of variation) were observed: flowering (31%), days to maturity (21%), reproductive period (18%), grain yield (48%), and 100-seed weight (18%). Combined ANOVA demonstrated an oversized impact of environment (81%) on yield, while genotype and GEI effects were 2% and 14%, respectively. The variables TMINF, TMINRP, and cumulative growing degree-day showed positive correlations with yield, while extended vegetative and maturity durations negatively influenced yield (p < 0.05). Additionally, linear mixed-models and PCA results explained that instability in crop phenology had significant influence on field pea yield. Seed weight was markedly varied within the locations (9.9–20.8 g) and both higher and lower seed weights were associated with lower yields (Optimal = 17.1 g). HA-GGE biplot-based on environment focus-scaling demonstrated three mega-environments and specific locations viz. Kota (CZ), SK Nagar (CZ), Raipur (CZ), Sehore (CZ), and Pantnagar (NWPZ) as the ideal testing-environments with high efficiency in selecting new genotypes with wider adaptability. The study findings highlight distinct impact of environments on crop phenology and agronomic traits of field pea (dwarf-type), hold substantial value in designing efficient field pea (dwarf-type) breeding program at mega-environment scale.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Field pea (Pisum sativum L. 2n = 14) is a cool-season pulse crop extensively cultivated in the Indian subcontinent, covering 0.76 million hectare (FAOSTAT 2022). Although originally a crop of temperate climates, it has been domesticated over the years and is now adapted to the tropical and subtropical climates of the Indian subcontinent (Siddique et al. 2013; Allchin 2017). However, in India, the productivity of field pea (1.38 t ha− 1) is quite lower than its global average yield of 1.97 t ha− 1 (FAOSTAT 2022). Consequently, considerable research attention is dedicated to enhancing the crop’s productivity potential through systematic breeding programs (Jimenez-Lopez et al. 2020; Parihar et al. 2022a). Field pea is a rich source of protein, slowly digestible starch, essential minerals, and dietary fiber while being low in fat, making it an excellent nutritious supplement to cereals (Singh and Pratap 2016; Rasskazova et al. 2020). Currently, it serves as a competitive crop for diversification of cereal-based cropping systems in irrigated agro-regions (Kumar et al. 2018; Baishya et al. 2019). The dwarf-type field pea cultivars exhibit significant differences from tall-type cultivars/breeding lines in terms of yield, adaptability to adverse environments, and responsiveness to inputs (Annicchiarico et al. 2019; Parihar et al. 2023).Evaluation and interpretation of multi-location data concerning crop performance across diverse climates can offer valuable insights to plant breeders, aiding in the development of cultivars adaptable to various environments (Rao et al. 2022; Das et al. 2019). Yet, identifying high-yielding and stable genotypes that excel in unpredictable and challenging conditions poses a significant challenge (Khan et al. 2021). To tackle this, breeders may focus on developing genotypes specifically tailored to particular environments or mega-environments. Hence, creating site/zone-specific breeding strategies becomes crucial, considering crop eco-physiological responses, ecological characteristics, and genotype-by-environment interaction (GEI). Field pea, originating from temperate climates, exhibits sensitivity to temperatures in India’s tropical/subtropical regions (Siddique et al. 2013; Lamichaney et al. 2021). The optimal temperature range for field pea is 15-25oC (Guilioni et al. 2003). The crop has higher temperature sensitivity compared to other cool-season pulses like chickpea, lentil, and studies indicate that temperature extremes (both lower and higher temperatures) have potential adverse effects on crop growth and developments (Sadras et al. 2012). Considering the substantial variability in the weather variables across India’s field pea-growing regions, these environmental factors can profoundly influence yields. However, investigations into the crop-environment relationship in these contexts remain scarce. Eco-physiological models can aid in identifying such genotypes, particularly in areas with unfavorable climates (Rötter et al. 2018). Crop phenological development, strongly influenced by growing environments, holds significant importance in determining yields in legume crops, including field pea. Additionally, understanding how crops interact with the environments of major field pea-growing regions would be invaluable in formulating strategic guidelines for field pea research in the country, an area that has yet to be adequately addressed.The All India Coordinated Research Project (AICRP) under Indian Council of Agricultural Research (ICAR), has conducted multi-location trials involving advanced breeding lines of dwarf-type field pea in collaboration with state agricultural universities across diverse climatic and geographical conditions in India (Bhuvaneswari et al. 2017; Das et al. 2019). AICRP testing locations serve as representative field pea growing regions, exhibiting significant diversity in terms of climatic conditions such as topography, rainfall, latitude, and altitude. Understanding GEI behavior is very important in breeding programs. This parameter provides important information to assess genotype stability across environments, forming breeding zones by leveraging adaptations of genotypes to specific environments and indicating genotypes suitable for particular environments to maximize genetic gains (Dias et al. 2018; Parihar et al. 2018). Various methods exist for analyzing and interpreting multi-environment trial (MET) data (Thangavel et al. 2011; Wang et al. 2016; Baraki et al. 2020; Singh et al. 2020). Characterizing locations based on representativeness, discriminativeness, and desirability indices following multi-genotype and multi-environment analysis provides criteria for selecting ideal testing locations for a crop species (Yan et al. 2007; Parihar et al. 2022b).
The study aimed to investigate the sensitivity of phenological events and yield traits in advanced dwarf-type field pea breeding lines across twenty test environments covering diverse agro-ecologies of India. The primary objectives of the study were (i) to demonstrate the variable crop response of dwarf-type field pea to diverse agro-regions and understanding G × E interaction (ii) to elucidate the impact of crop-stage-specific weather variables on crop performance and yield traits using multi-location data, and (iii) to identify and characterize core testing sites within each mega-environment for cultivar evaluation, release, and cultivation across zones in the Indian subcontinent.
Materials and methods
Study environments and weather variables
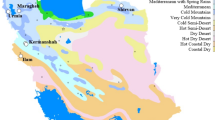
The trial locations encompassed all field pea (dwarf-type) growing ecologies across India (Fig. 1), spanning latitudes from 21.40°N (Raipur, Chhattisgarh) to 33.98°N (Srinagar, Jammu and Kashmir), longitudes from 72.19°E (SK Nagar, Gujrat) to 91.31°E (Agartala, Tripura), and altitudes from 18.0 m (Berhampore) to 1617.0 m (Srinagar) above sea level. These locations reside within distinct, well-established zones for cultivating dwarf-type field pea, namely CZ (Central Zone), NWPZ (North Western Plain Zone), NEPZ (North Eastern Plain Zone), and NHZ (Northern Hill Zone), each characterized by diverse ecological conditions for field pea crops. While the Southern Zone of India is a major pulse-growing area, its environmental conditions are not conducive to field pea cultivation due to higher than ambient temperatures during the crop season. Each individual zone comprises several states: the CZ includes Madhya Pradesh, Chhattisgarh, Gujarat, South part of Rajasthan, Bundelkhand region of Uttar Pradesh; NEPZ encompasses Eastern Uttar Pradesh, Assam, Bihar, Jharkhand, West Bengal; NWPZ comprises the Plains of Uttarakhand, Punjab, Haryana, Western Uttar Pradesh, Rajasthan; NHZ includes Jammu and Kashmir, Hills of Uttarakhand, and the North Eastern Hill region. The study included seven, four, six, and three locations in CZ, NWPZ, NEPZ, and NHZ, respectively (Table 1).

The multi-location study encompassed major field pea (dwarf-type) growing areas across diverse agro-ecological zones in India. Triangle symbols in green, pink, red, and blue represent the Central Zone (n = 7), North Western Plain Zone (n = 4), North Eastern Plain Zone (n = 6), and Northern Hill Zone (n = 3), respectively. These symbols correspond to location numbers 1 to 20 as listed in Table 1
Multi-location testing of genotypes and crop management
During the year 2017–2018, station trials were conducted with ten superior genotypes of dwarf-type field pea contributed by developers from seven research centers, along with their respective zonal checks. Based on their performance at respective stations, one or two of the most superior genotypes were selected for multi-location testing. As a result, seven research centers from four states of India contributed a total of twelve genotypes (advanced breeding lines) for multilocation testing, as detailed in Supplementary Table 1. These twelve genotypes were developed through hybridization. Subsequently, these genotypes underwent nationwide testing in twenty locations across India during the winter season of 2018–2019 under the auspices of the All India Coordinated Research Project (AICRP) on MULLaRP in Kanpur, India. The selected genotypes were evaluated in each location following a randomized complete block design (RCBD) with three replications. The field was prepared by ploughing, harrowing, and planking, with 20 kg N, 40 kg P2O5, and 40 kg K2O applied during field preparation using diammonium phosphate (DAP, 46% P2O5, 18% N), urea (46% N), and muriate of potash (60% K2O). The quantity of DAP was determined based on the rate of P applied, and then the quantity of N supplemented with the applied dose of DAP was calculated. The remaining amount of N was applied using urea. Each genotype was planted in six rows, each spanning a length of 4 m, following a planting geometry of 25.0 cm × 5.0 cm. To maintain a weed-free crop, one or two rounds of hand weeding were carried out. Additionally, during the crop season, 1–4 irrigations were applied, considering the specific rainfall patterns of each location. Essential plant protection measures were also implemented to ensure healthy crop growth.
Phenological observations
The crop’s phenological events like days to 50% flowering (DTF) and days to maturity (DTM) were determined by visual observation (2-day interval) for each genotype. The days taken after sowing until 50% of plants in a plot had one open flower represented the DTF. Likewise, the days taken after sowing to full crop maturity i.e., when all pods (upper pods in particular) turned to a light-yellow color, was calculated as DTM. The number of days between DTF and DTM, for each genotype was computed and represented as reproductive period (RP). The ratio of days to 50% flowering (DTF) to days to maturity (DTM) was denoted as [DTF/DTM].
Estimation of grain yield and 100-seed weight
For estimating the grain yield, the inner 4 rows each of 4 m length was harvested separately. The harvested seeds were allowed to dry under sunlight to reduce moisture content to about 10–12%. Initially, plot wise assessment of grain yield was done, which was subsequently converted to kg ha− 1. A sample of hundred seeds were counted in three replications and weighed for 100-seed weight estimation.
Temperature intensity and growing degree-day (GDD) calculation
Utilizing the crop season weather data from all testing locations, we determined the maximum, minimum, and average temperatures (°C) during flowering, denoted as TMAXF, TMINF, TAVGF, respectively following the calculations as detailed by Lamichaney et al. (2021). For example, to calculate the maximum temperature intensity during the flowering stage, we computed the average of the daily maximum temperature (oC) during flowering time for each genotype, which represents the average maximum temperature during the seven-day period (i.e., at 50% flowering ± 3 days). Likewise, TMINF, TAVGF were calculated following same calculation as mentioned above using the daily ambient minimum temperature and ambient average temperate, respectively. Meanwhile, we calculated the maximum, minimum, and average temperatures (in °C) during the reproductive period by averaging the daily temperature from 50% flowering to maturity, designated as TMAXRP, TMINRP, TAVGRP, respectively. The cumulative daily rainfall received during the vegetative (sowing to 50% flowering), reproductive (50% flowering to maturity), and full crop season (sowing to maturity) of each genotype was calculated and denoted as RFV, RFRP, RFT, respectively. Crop stage-specific weather variables were computed for each genotype across all locations. Cumulative growing degree–days were calculated for the vegetative, reproductive, and full crop season of each field pea genotype and denoted as GDDV, GDDRP, and GDDFCS, respectively. The cumulative GDD value at different crop growth stages was calculated using the following formula:
Where Tmax and Tmin are the daily maximum and minimum temperatures. The calculation was done using 0°C as the base-temperature for field pea (Lake et al. 2021).
Statistical analysis
Analysis of variance (ANOVA) was used to explain the significant differences observed in all treatments, by investigating the effects of genotypes (G), environments (E), and genotype-environment interaction (G × E) on grain yield data. The R-program 3.2.1 was used to devise the contribution of each factor and their interactions with grain yield. The broad sense heritability (H) was calculated using the following formula:
Where, σ2p and σ2g denotes for phenotypic and genotypic variance, respectively; SD and SE represents standard deviation and standard error, respectively; and n denotes for replication number.
To approximate the stability of genotypes across different locations, a numerical and graphical analysis was performed using the HA-GGE biplot analysis (Yan and Holland 2010). This analysis displays the effects of genotype (G) and genotype-environment interaction (G × E) on multi-environment data through a graphical representation. The HA-GGE biplot analysis was conducted using the calculated heritability in R-program 3.2.1. The basic model given by Yan 2010 was followed for GGE biplot analysis. The statistical detail for the HA-GGE biplot analysis is elaborated in Parihar et al. (2022b).
Multivariate linear regression models were derived to explain the relative weightage of the selected variables as a function of grain yield. The regression line for a multivariable regression is as follows;
Where, Y = dependent variable, X1 − n = independent variables, a = constant (y-intersect), b1 − n = regression coefficient of the variable.
Principal component analysis was performed using PAST software (version 4.03) to analyze the percent contribution of the major components considering the weather variables and plant yield attributes and graphically represent the variables vectors on principal coordinates. Heat-map cum cluster analysis was done using the online statistical program Clustvis - a web tool for visualizing clustering of multivariate data (BETA) (https://biit.cs.ut.ee/clustvis/) visually representing cluster presentation of genotypes and environments.
Results
Site characteristics and weather variables
Six out of twenty locations experienced TMAXF > 25oC, with Agartala recording the highest (28.3oC) (Supplementary Fig. 1). Bhatapara had the highest TMINF at 18.0 °C, while notably lower temperatures (< 5 °C) were observed in Hisar and Ranchi. The range observed for TMAXRP and TMINRP was 15.0–30.8 °C and 7.9–22.1 °C, respectively, with nine locations having TMAXRP exceeding 25 °C. Bhatapara registered TMINRP above 20 °C, significantly higher than the all location mean TMINRP (11.7 °C). GDDV was lowest in Srinagar (84.1 °C-day) and varied from 537 to 935 °C-day in other locations. GDDRP was notably lower in NHZ (Srinagar, Agartala, Almora) compared to other zones. NHZ locations received substantial rainfall (126–614 mm) during the crop season, whereas other locations/zones ranged between 0 and 86 mm (Supplementary Fig. 1).
Crop phenological developments and yield traits
The results indicated that Srinagar environment notably extended DTF (+ 86 days), DTM (+ 87 days), DTF/DTM (+ 27%) over the mean values across all locations (Fig. 2). Among the locations, Samba, Srinagar, Almora exhibited higher DTF, DTM, and DTF/DTM. Bhatapara, Agartala, and Raipur showcased a noticeable shorter duration of maturity (DTM) by 25–33 days. The reproductive period was shorter in Raipur and Bhatapara (> 10 days) while longer in Pantnagar, Samba, Hisar, and Ranchi (> 5 days) compared to the mean value of 48 days across all locations (Fig. 2). Locations like SK Nagar, Durgapura, and Hisar demonstrated a significant yield increase (2787–3731 kg ha− 1; > 1000 kg over the all location mean), while Almora and Dholi reported substantially lower yields (217–719 kg ha− 1). The parameters DTF, DTM, DTF/DTM followed the order of NHZ > NWPZ > NEPZ > CZ, while RP was in the order of NWPZ > NEPZ > CZ > NHZ (Supplementary Fig. 2). CZ (2.10 t ha− 1) and NWPZ (2.11 t ha− 1) exhibited higher grain yields, followed by NEPZ (1.44 t ha− 1), with NHZ recording the lowest (0.98 t ha− 1). NEPZ and NHZ reported higher seed weights compared to CZ and NWPZ (p < 0.05) (Supplementary Fig. 2). Among the genotypes, IPFD 18 − 8 and IPFD 18 − 2 emerged as the most productive with all location average of 1921.3 kg ha− 1 and 1940.3 kg ha− 1, respectively, while HUDP 1711 was recorded with the lowest all location mean yield of 1564.8 kg ha− 1 (Supplementary Tables 2 & 3). Among the genotypes, IPFD 18 − 2 demonstrated the highest yield variability [coefficient of variation (CV) = 59.6%] within the locations, while HUDP 1711 exhibited the lowest yield variations (CV = 38.9%) (Supplementary Table 3).

Location-wise changes in crop phenological events (a-d), seed weight (e), and grain yield (f) of field pea compared to all location mean values. Error bars represent standard error of genotypes mean values. Locations sharing the same color bar signify a particular zone [CZ (light brown), NWPZ (purple), NEPZ (light green), and NHZ (bright pink)]. For detailed environmental information and abbreviations please see Table 1
Correlations (crop-environment relationships)
Field pea grain yield exhibited significant positive correlations with TMINF (r = + 0.15; p = 0.025) and TMINRP (r = + 0.23; p < 0.001) (Fig. 3). Additionally, all GDD variables displayed significant positive associations with yield (r = + 0.13 to + 0.27; p < 0.05), with the highest for GDDFCS. Conversely, significant negative correlations were observed between yield and rainfall variables (RFV, RFRP) (p < 0.001) (Fig. 3). Specifically within CZ, total crop season rainfall (RFT), TMAXF, and TMAXRP exhibited positive correlations with yield. In contrast, total seasonal rainfall and TMAXRP showed negative correlation with grain yield in NEPZ. Moreover, DTF and DTM demonstrated a negative correlation with yield, whereas the correlation between yield and RP was non-significant (Fig. 3). An evident quadratic (second-order polynomial) relationship existed between HSW (ranging between 8.7 and 26.2 g) and yield (p < 0.01), with the optimal seed weight associated with the highest yield being 17.1 g (Supplementary Fig. 3).

Distribution pattern of crop and weather variables of dwarf-type field pea genotypes grown in different locations across India (data point = 256) (a). Relationship of field pea (dwarf-type) yield with phenological parameters, 100-seed weight, and crop-stage specific weather variables in different zones (b-f). Green, red and gray color horizontal bars represents significant positive, significant negative and non-significant correlations at probability level of 5%. TCC0.05 = threshold correlation coefficient at p = 0.05 to discriminate significant from non-significant results. DTF, days to flowering; DTM, days to maturity; RP, reproductive period; HSW, 100-seed weight; TMAXF maximum temperature during flowering (oC); TMINF minimum temperature during flowering (oC); TMAXRP, maximum temperature during reproductive period (oC); TMINRP minimum temperature during reproductive period (oC); GDDV cumulative growing degree-days of vegetative period (oC-day); GDDRP cumulative growing degree-days of vegetative period (oC-day); GDDFCS cumulative growing degree-days of full crop season (oC-day); RFV cumulative rainfall during vegetative period (mm); RFRP cumulative rainfall during reproductive period (mm); RFT cumulative rainfall during full crop season (mm)
Multivariate analysis results (PCA, cluster analysis, linear mixed model)
According to PCA results, DTF, DTM, RFRP, RFV displayed negative associations with yield (Fig. 4a). Cluster analysis demonstrated that environments grouped into three sub-clusters: [Srinagar, Samba, Almora], [Raipur, Bhatapara], and the remaining fifteen locations in another cluster (Fig. 4b). PCA considering only genotypic variations (Supplementary Table 4) illustrated negative association between yield and DTM and strong positive associations with HSW (Supplementary Fig. 4). Results from multivariate linear regression models elucidated that weather variables altered significantly within the locations, significantly impacting yield (Table 2). Nonetheless, the extent of influence (estimated t-stat value) of different variables notably varied within the zones. For instance, among the independent variables [TMAXF, TMINF, GDDV, GDDRP, RFV, RFRP], TMAXF had a noticeable influence on yield in CZ, while TMINF was found to be a significant grain yield-influencing factor in CZ (negatively) and NHZ (positively). A higher GDDV negatively influenced the yield in NEPZ, while the significant impact of RFRP on grain yield was evident in CZ and NWPZ. Considering overall variability across the location, the regression models highlighted that reproductive period (positive) and vegetative period (negative) were significant factors affecting field pea grain yield (Table 3).

Principal component analysis with vector axis of component variables to represents inter-correlations within variables (n = 256) (a). Cluster analysis with heat map presentation of different crop and crop-stage specific weather parameters to explain their associations within environments (b). GY, grain yield, DTF, days to flowering; DTM, days to maturity; RP, reproductive period; HSW, 100-seed weight; TMAXF, maximum temperature during flowering (oC); TMINF minimum temperature during flowering (oC); TMAXRP, maximum temperature during reproductive period (oC); TMINRP minimum temperature during reproductive period (oC); GDDV cumulative growing degree-days of vegetative period (oC-day); GDDRP cumulative growing degree-days of vegetative period (oC-day); GDDFCS cumulative growing degree-days of full crop season (oC-day); RFV cumulative rainfall during vegetative period (mm); RFRP cumulative rainfall during reproductive period (mm); RFT cumulative rainfall during full crop season (mm). Please see Table 1 for environment detail
Genotype× environment interactions and HA-GGE biplot
The genotype (G), environment (E), and GEI accounted for 2.0%, 80.7%, and 14.2% of the overall variation in yield, respectively (Table 4). Meanwhile, zone-wise ANOVA estimates revealed that G, zone and genotype × zone accounted for 4.6%, 83.6%, and 10.4%, respectively (p < 0.001) (Table 4). Efficiently explaining 50.6% of the total variation, the HA-GGE biplot outlined a seven-vertex polygon biplot (Fig. 5). It identified three mega-environments (each with at least three locations): [Samba, Dholi, Almora, Agartala, Berhampore, Pantnagar], [SK Nagar, Raipur, Durgapura], and [Kota, Sehore, Bhatapara, Hisar, Varanasi, Sagar]. Considering both mean and stability, PANT P 415 and PANT P 418 were identified as the most superior and adaptable genotypes among tested genotypes (Fig. 5). The representativeness scale within the studied environments ranged from +3.14 to + 3.98, while the discriminative power varied from − 0.31 to + 0.84 (Fig. 5). The desirability index, representing the overall performance of a location based on both ‘discriminatory’ power and ‘representativeness’ ranged from − 1.25 to + 3.15. Locations such as Kota (CZ), SK Nagar (CZ), Raipur (CZ), Sehore (CZ), and Pantnagar (NWPZ) were identified as the ideal testing environments, demonstrating high efficiency in selecting new genotypes with wider adaptability (Fig. 5).

‘Which-won-where’ (a), and Mean vs. Stability (b) view of test locations based on heritability-adjusted GGE (HA-GGE) biplot analysis of dwarf-type field pea genotypes across twenty testing locations. No transformation of data (transform = 0); and data were centered by means of the environments (centering = 2). Numbers correspond to genotypes and environment as listed in Tables 1 and 2, respectively. Representativeness (c), discriminativeness (d), and desirability index (e) of different study locations for yield potential of 16 promising lines of dwarf type field pea genotypes based on heritability-adjusted GGE (HA-GGE) biplot analysis. Please see Table 1 for environment detail
Discussion
The results indicate that the major field pea (dwarf-type) growing regions exhibit significant variations in temperature conditions specific to different crop stages. Notably, the highest temperature difference among all locations was observed for TMAXRP (15.8oC) followed by TMINF (14.8oC), TMINRP (14.1oC), TMAXF (9.5oC). These differences in crop-season temperatures certainly are the reason for the greater impact size of environmental factors on the crop’s phenological development and productivity of field pea. The climate of NHZ locations was distinct from other zones in terms of temperature conditions and rainfall, contributing significantly to a greater scale of variations in crop performance and yield traits. Thus, enhancing our comprehension of crop responses to weather variables and subsequently devising strategies to narrow yield gaps within specific locations becomes crucial.
Correlation findings across all zones suggest that maximum temperatures (TMAXF and TMAXRP within the observed range) do not significantly influence field pea yields. However, zone-wise correlation results indicate that higher maximum temperatures during the reproductive period adversely affect yields in NEPZ. Reports suggest that temperatures exceeding 22–25 °C, especially above 25 °C during grain filling, can affect field pea yields (Guilioni et al. 2003; Sadras et al. 2013). Additionally, a notable positive correlation between TMINF and TMINRP and grain yield implies that lower minimum temperatures (or cold stress) during grain filling limit crop productivity. Notably, in eight of twenty locations, TMINF was < 8 °C, while TMINRP was < 10 °C in seven locations, signifying substantial exposure to lower temperatures with significant yield implications. Studies have highlighted field pea’s sensitivity to low temperatures (chilling injury at < 15 °C).The influence of ambient temperatures on yield is intricate, variable, and site-specific, contingent on other vital growth factors that determine yield. Results indicate an inverse relationship between higher rainfall and yield. Legumes like field pea are generally sensitive to both excessively high soil moisture conditions (Deak et al. 2019) and higher rainfall events, especially with increased intensity, which might affect belowground crop growth, including nodulation in winter legumes (Ali et al. 2000).
Our findings underscore the significant impact of the environment on the eco-phenological development of the crop, particularly on flowering and maturity durations. Specifically, locations with low temperatures like Srinagar and Almora (NHZ) and Samba (NWPZ) experienced prolonged vegetative phases. Srinagar notably showed the most significant extension, surpassing the average vegetative phase by more than 80 days across all locations. Environmental effects on phenological dynamics are particularly noticeable in legumes (Sennhenn et al. 2017). Although field pea is primarily cultivated in temperate climates like Canada, Russia, and China (Joshi et al. 2023), its domestication has allowed adaptation to Indian subtropical climates, resulting in a shortened crop season compared to other cool-season legumes (Tafesse et al. 2020). Multiple studies suggest that the length of the grain-filling period directly influences yields in cool-season legumes, including field pea (Farooq et al. 2017; Sita et al. 2017). Notably, CZ and NWPZ emerged as the most productive field pea growing zones with comparable yields. However, these agro-ecological zones (CZ, NWPZ) exhibit significant differences in the reproductive period, spanning from 44 to 54 days. This outcome implies that the effect of the grain-filling period on yield might have been influenced by other crucial factors affecting yield, such as weather conditions. Considering the substantial influence of diverse environments, this study emphasizes the necessity of developing breeding lines with narrow elasticity in crop phenology, particularly in response to variable climatic conditions. This adaptation is essential for improved crop performance across environments, especially in Indian climates where extremes of both cold (NHZ, NWPZ) and heat (CZ and NEPZ) are prevalent in field pea (Anonymous 2019).
Our research highlights that environment-induced alterations in phenological development have a direct influence on grain yield. Specifically, both the vegetative and maturity durations exhibit negative correlations with yields. Our multivariate linear regression models demonstrate that these periods significantly influence yield but at varying degrees. The statistical analysis emphasizes a more pronounced negative impact of prolonged vegetative phases on yield compared to a positive influence from the reproductive period. The consistent decrease in yield due to extended vegetative and maturity durations are observed across genotypes, albeit at different levels. Notably, nine out of twelve genotypes exhibited significant yield reductions with increased vegetative duration (Supplementary Table 3). These strong correlations predominantly arise from substantial changes in crop phenological development in the NHZ and partly from NWPZ. Therefore, our study underscores that prolonged vegetative and overall maturity periods, especially in low-temperature regions, pose significant challenges for field pea in sustaining yields. Consequently, efforts should prioritize the identification of photo-thermo-insensitive breeding lines to foster the development of phenologically stable cultivars adaptable to India’s diverse agro-ecologies (Pratap et al. 2014; Gowda et al. 2015; Basu et al. 2016). Furthermore, since the NHZ is an unconventional area for field pea, creating adaptable cultivars for this region can greatly enhance crop cultivation, addressing the escalating demand for pulses in the country. The environment significantly impacted seed size/weight, ranging from 9.9 to 20.8 g. Seed weight exhibited a quadratic response function concerning yield (Supplementary Fig. 3), indicating an optimal seed size of 17.1 g linked to higher yield. Seed size is notably influenced by pod size and the number of effective pods in field pea (Jiang et al. 2020). Therefore, understanding the effect of crop-growing environments on seed size/weight in relation to pod number, seed set percentage, and mechanisms that balance source-sink dynamics during grain filling, particularly in stressful environments, is crucial (Poggio et al. 2005; Lamichaney et al. 2021). Given the considerable variability observed in seed size, there is a critical need to concentrate breeding efforts on stabilizing this trait to ensure yield stability across diverse locations.
A comprehensive understanding of GEI characteristics, particularly in specific locations, is pivotal in national-scale plant breeding endeavors (Baraki et al. 2020; Parihar et al. 2022b). Identifying mega-environments and key locations for breeding/testing assumes fundamental importance in this context (Tabasum et al. 2010; Yan et al. 2011). Presently, unlike major cool-season crops like chickpea and lentil, there are few systematic field pea breeding stations in India aiming to develop cultivars for the entire country or region. This limitation results in a primary reason for broader environmental size effects, where promising entries developed/bred from one zone/agro-ecology may not perform well in others due to significant variations in climatic conditions. HA-GGE biplot analysis, explaining 50% of the total variation, identified seven key vertices within the polygon biplot. The findings from the biplot are highly relevant to the study, with environmental size effects accounting for a high percentage (81%) of total yield variations (Yan et al., 2000). The “which-won-where” biplot revealed three mega-environments among the tested locations. This analysis highlighted distinct performance differences, with three out of the twelve breeding lines showing their best performance in different mega-environments. Our results showcase the identification of superior genotypes for specific mega-environments, such as genotype RFP 12-08 (developed from Ranchi) for mega-environment 1, PANT P418 (developed from Pantnagar) for mega-environment 2, IPFD 18-8 (developed from Kanpur) for mega-environment 3, and PANT P 415 (developed from Pantnagar) suitable for Srinagar conditions. Additionally, the HA-GGE biplot demonstrated the relative utility of environments in terms of selection response, with strategic mega-environments identified among the tested locations. The biplot effectively scaled the discriminating power and representativeness of environments with wider ranges, identifying Kota (CZ), SK Nagar (CZ), Raipur (CZ), Sehore (CZ), and Pantnagar (NWPZ) as ideal environments for testing field pea genotypes based on their discriminative power, representativeness, and desirability index. Such selection of ideal testing locations will help in better characterization of the genetic yield potential of advanced breeding lines and offer cost-effective multi-environment testing of genotypes with minimum numbers of testing locations (Yan et al. 2011; Alam et al. 2014; Baraki et al. 2020). The study information holds significant value in designing efficient breeding programs both on regional (zone) and national scales. Greater emphasis is warranted on the crop’s thermo-insensitivity, phenological stability, and seed size as traits to enhance crop performance across diverse climates of India. Furthermore, considering the limited genotypic diversity in the crops, breeding efforts are warranted to enhance genetic diversity and improve resilience against challenging edapho-climatic conditions.
Conclusions
The study highlighted significant impacts of diverse environments on field pea (dwarf-type) phenological development, seed traits, and yield. The environmental effect on yield accounted for 81%, while the contribution of genotype and genotype-by-environment interaction (GEI) was minimal, at 2% and 14%, respectively. The study indicated the substantial influence of temperature conditions on crop yield potential, particularly adverse effects of lower temperatures (cold-stress) observed in some locations of NHZ and NWPZ. Conversely, in NEPZ, higher temperatures led to lower yields. Delays in crop phenological development, both in the vegetative and crop maturity duration, due to lower temperature conditions, resulted in yield reductions, as confirmed by linear regression and PCA results. Moreover, seed size as a trait exhibited high variability across different environments, highlighting the need for a more detailed understanding of this trait in relation to environmental factors and associated crop parameters to achieve greater stability in the trait as well as in the yield. The study indicated a potential scope in harnessing GEI (given its higher contribution over the genotype factor). However, the overwhelming influence of the environment poses a challenge for efficient breeding programs aimed at developing materials with wider adaptation in the Indian environment. In this context, the identification of mega-environments and characterizing specific locations becomes crucial for strategic breeding and testing within national crop improvement programs. The study identified three distinct mega-environments within the tested locations. Locations such as Kota (CZ), SK Nagar (CZ), Raipur (CZ), Sehore (CZ), and Pantnagar (NWPZ) were pinpointed as ideal testing environments with high efficiency in selecting new genotypes with broader adaptability. These study findings hold significant utility in developing crop-breeding strategies for dwarf-type field pea at both regional and national scales. Additionally, strategic breeding approaches should emphasize the crop’s thermo-insensitivity, phenological stability, and seed size traits to achieve broader adaptation and enhanced yield performance. A systematic pre-breeding program to enhance genetic variability would further complement these efforts.
References
Alam AM, Somta P, Jompuk C, Chatwachirawong P, Srinives P (2014) Evaluation of mungbean genotypes based on yield stability and reaction to mungbean yellow mosaic virus disease. Plant Pathol J 30(3):261–268
Ali M, Dahan R, Mishra JP, Saxena NP (2000) Towards the more efficient use of water and nutrients in food legume cropping. In: Linking Research and Marketing Opportunities for Pulses in the 21st Century: Proceedings of the Third International Food Legumes Research Conference 2000 Springer Netherlands, pp. 355–368
Allchin FR (2017) Early cultivated plants in India and Pakistan. In: The domestication and exploitation of plants and animals, Routledge, pp. 323–330
Annicchiarico P, Collins RP, De Ron AM, Firmat C, Litrico I, Hauggaard-Nielsen H (2019) Do we need specific breeding for legume-based mixtures? Adv Agron 157:141–215
Anonymous (2019) Project coordinators Report (2018-19). Indian Council of Agricultural Research. Indian Institute of Pulses Research, Kanpur. All India Coordinated Research Project on MULLaRP
Baishya LK, Rathore SS, Sarkar D, Jamir T, Rajkhowa DJ (2019) Crop and varietal diversification for enhancing productivity and profitability of rice fallow system in eastern himalayan region. Indian J Agric Sci 89(5):800–805
Baraki F, Gebregergis Z, Belay Y, Berhe M, Zibelo H (2020) Genotype × environment interaction and yield stability analysis of mung bean (Vigna radiata (L.) Wilczek) genotypes in Northern Ethiopia. Cogent Food Agric 6(1):1729581
Basu PS, Singh U, Kumar AN, Praharaj CS, Shivran RK (2016) Climate change and its mitigation strategies in pulses production. Indian J Agron 61:S71–82
Bhuvaneswari S, Sharma SK, Punitha P, Shashidhar KS, Naveenkumar KL, Prakash N (2017) Evaluation of morphological diversity of field pea [Pisum sativum subsp. arvense (L.)] germplasm under sub-tropical climate of Manipur. Leg Res 40(2):215–223
Das A, Parihar AK, Saxena D, Singh D, Singha KD, Kushwaha KP, Chand R, Bal RS, Chandra S, Gupta S (2019) Deciphering genotype-by-environment interaction for targeting test environments and rust resistant genotypes in field pea (Pisum sativum L). Front Plant Sci 10:825
Deak EA, Martin TN, Fipke GM, Stecca JD, Tabaldi LA, Nunes UR, Winck JE, Grando LF (2019) Effects of soil temperature and moisture on biological nitrogen fixation in soybean crop. Aust J Crop Sci 13(8):1327–1334
Dias KO, Gezan SA, Guimarães CT, Nazarian A, da Costa e Silva L, Parentoni SN, de Oliveira Guimarães PE, de Oliveira Anoni C, Pádua JM, de Oliveira Pinto M, Noda RW (2018) Improving accuracies of genomic predictions for drought tolerance in maize by joint modeling of additive and dominance effects in multi-environment trials. Heredity 121(1):24–37
FAOSTAT (2022) Food and Agriculture Organization of the United Nations. Rome: FAO. Available online at: http://www.fao.org/faostat/en/#data (assessed February, 2023)
Farooq M, Nadeem F, Gogoi N, Ullah A, Alghamdi SS, Nayyar H, Siddique KH (2017) Heat stress in grain legumes during reproductive and grain-filling phases. Crop Pasture Sci 68(11):985–1005
Gowda CL, Chaturvedi SK, Gaur PM, Sameer Kumar CV, Jukanti AK (2015) Pulses research and development strategies for India. (https://oar.icrisat.org/9413/1/Pulses%20Res%20and%20Dev-CLLG.pdf)
Guilioni L, Wéry J, Lecoeur J (2003) High temperature and water deficit may reduce seed number in field pea purely by decreasing plant growth rate. Funct Plant Biol 30(11):1151–1164
Jiang Y, Lindsay DL, Davis AR, Wang Z, MacLean DE, Warkentin TD, Bueckert RA (2020) Impact of heat stress on pod-based yield components in field pea (Pisum sativum L). J Agron Crop Sci 206(1):76–89
Jimenez-Lopez JC, Singh KB, Clemente A, Nelson MN, Ochatt S, Smith PM (2020) Legumes for global food security. Front Plant Sci 11:926
Kumar N, Hazra KK, Nath CP, Praharaj CS, Singh U (2018) Grain legumes for resource conservation and agricultural sustainability in South Asia. In: Legumes for soil health and sustainable management. pp. 77–107
Lake L, Guilioni L, French B, Sadras VO (2021) Field pea. Crop physiology case histories for major crops. Academic, pp 320–341
Lamichaney A, Parihar AK, Hazra KK, Dixit GP, Katiyar PK, Singh D, Singh AK, Kumar N, Singh NP (2021) Untangling the influence of heat stress on crop phenology, seed set, seed weight, and germination in field pea (Pisumsativum L). Front Plant Sci 12:635868
Parihar AK, Basandrai AK, Kushwaha KP, Chandra S, Singh KD, Bal RS, Saxena D, Singh D, Gupta S (2018) Targeting test environments and rust-resistant genotypes in lentils (Lens culinaris) by using heritability-adjusted biplot analysis. Crop Pasture Sci 69(11):1113–1125
Parihar AK, Yadav R, Lamichaney A, Mishra RK, Chandra A, Gupta DS, Tripathi K, Hazra KK, Dixit GP (2022a) Field pea breeding. Fundamentals of Field Crop breeding, 2022 May 6. Springer Nature Singapore, Singapore, pp 1237–1321
Parihar AK, Gupta S, Hazra KK, Lamichaney A, Sen Gupta D, Singh D, Kumar R, Singh AK, Vaishnavi R, Jaberson MS, Das SP et al (2022b) Multi-location evaluation of mungbean (Vigna radiata L.) in Indian climates: ecophenological dynamics, yield relation, and characterization of locations. Front Plant Sci 13:984912
Parihar AK, Hazra KK, Lamichaney A, Singh AK, Dixit GP (2023) Delineating the role of plant stature towards heat stress tolerance in field pea (Pisum sativum L). Heliyon 9(3):e14539
Pratap A, Basu PS, Gupta S, Malviya N, Rajan N, Tomar R, Madhavan L, Nadarajan N, Singh NP (2014) Identification and characterization of sources for photo- and thermo-insensitivity in Vigna species. Plant Breed 33(6):756–764
Rao P, SandhyaKishore N, Sandeep S, Neelima G, Rao PM, Das DM, Saritha A (2022) Evaluation of performance and yield stability analysis based on AMMI and GGE-biplot in promising pigeonpea [Cajanus cajan (L.) Millspaugh] genotypes. Leg Res I45(11):1414–1420
Rasskazova I, Kirse-Ozolina A (2020) Field pea Pisum sativum L. as a perspective ingredient for vegan foods: a review. Res Rural Dev 35:125–131
Rötter RP, Hoffmann MP, Koch M, Müller C (2018) Progress in modelling agricultural impacts of and adaptations to climate change. Curr Opin Plant Biol 45:255–261
Sadras VO, Lake L, Chenu K, McMurray LS, Leonforte A (2012) Water and thermal regimes for field pea in Australia and their implications for breeding. Crop Pasture Sci 63(1):33–44
Sadras VO, Lake L, Leonforte A, McMurray LS, Paull JG (2013) Screening field pea for adaptation to water and heat stress: associations between yield, crop growth rate and seed abortion. Field Crops Res 150:63–73
Sennhenn A, Njarui DM, Maass BL, Whitbread AM (2017) Understanding growth and development of three short-season grain legumes for improved adaptation in semi-arid Eastern Kenya. Crop Pasture Sci 68(5):442–456
Siddique KH, Erskine W, Hobson K, Knights EJ, Leonforte A, Khan TN, Paull JG, Redden R, Materne M (2013) Cool-season grain legume improvement in Australia-use of genetic resources. Crop Pasture Sci 64(4):347–360
Singh NP, Pratap A (2016) Food legumes for nutritional security and health benefits. In: Singh U, Praharaj C, Singh S, Singh N (eds) Biofortification of Food crops. Springer, New Delhi, pp 41–50
Singh B, Das A, Parihar AK, Bhagawati B, Singh D, Pathak KN, Dwivedi K, Das N, Keshari N, Midha RL, Kumar R (2020) Delineation of genotype-by-environment interactions for identification and validation of resistant genotypes in mungbean to root-knot nematode (Meloidogyne incognita) using GGE biplot. Sci Rep 10(1):4108
Sita K, Sehgal A, Hanumantha Rao B, Nair RM, Vara Prasad PV, Kumar S, Gaur PM, Farooq M, Siddique KH, Varshney RK, Nayyar H (2017) Food legumes and rising temperatures: effects, adaptive functional mechanisms specific to reproductive growth stage and strategies to improve heat tolerance. Front Plant Sci 8:1658
Tabasum A, Saleem M, Aziz I (2010) Genetic variability, trait association and path analysis of yield and yield components in mungbean (Vigna radiata (L.) Wilczek). Pak J Bot 42:3915–3924
Tafesse EG, Gali KK, Lachagari VR, Bueckert R, Warkentin TD (2020) Genome-wide association mapping for heat stress responsive traits in field pea. Int J Mol Sci 21(6):2043
Thangavel P, Anandan A, Eswaran R (2011) AMMI analysis to comprehend genotype-by-environment (G× E) interactions in rainfed grown mungbean (Vigna radiata L). Aust J Crop Sci 5(13):1767–1775
Wang J, Zhang Z, Gao X (2016) Modeling analysis of Mungbean in Regional trials with partial least squares regression. Adv J Food Sci Technol 11:728–733
Yan W, Holland JB (2010) A heritability-adjusted GGE biplot for test environment evaluation. Euphytica 171:355–369
Yan W, Kang MS, Ma B, Woods S, Cornelius PL (2007) GGE biplot vs. AMMI analysis of genotype-by-environment data. Crop Sci 47(2):643–653
Yan W, Pageau D, Frégeau-Reid J, Lajeunesse J, Goulet J, Durand J, Marois D (2011) Oat mega-environments and test-locations in Quebec. Can J Plant Sci 91(4):643–649
Acknowledgements
The authors express gratitude to the Indian Council of Agricultural Research, New Delhi, and the Indian Institute of Pulses Research, Kanpur, for their provision of essential facilities during the multi-location trials conducted under the AICRP on MULLaRP, based in Kanpur, Uttar Pradesh. The authors extend their sincere thanks to all the breeders who provided their entries for evaluation in these trials. Additionally, heartfelt appreciation is extended to all the center in-charges for generously providing experimental land and other crucial research facilities.
Funding
This research received no specific grant from any funding agency.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Competing interests
The author(s) declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Parihar, A.K., Hazra, K.K., Lamichaney, A. et al. Multi-location evaluation of field pea in Indian climates: eco-phenological dynamics, crop-environment relationships, and identification of mega-environments. Int J Biometeorol (2024). https://doi.org/10.1007/s00484-024-02720-7
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00484-024-02720-7